426
Copyright © 2021 The Korean Society of Fisheries and Aquatic Science pISSN:0374-8111, eISSN:2287-8815
서 론
대왕붉바리는 온대성어류인 붉바리
(Epinephelus akaara♀)
와아열대성대왕바리(E. lanceolatus♂)
의교잡종으로여름철 고수온기간을이용해집중양성할수있는대체양식어종으로 국내에서개발되었다.
염분은수서생물의서식지분포를결정 짓고생리적과정에영향을미치는중요한환경요인중하나이 며,
염분변화는어류의대사에직접적으로영향을미칠수있다(Boeuf and Payan, 2001; Nordlie, 2009; Anni et al., 2016).
일 반적으로해양경골어류는염분7.0-58.5 psu
의넓은범위에도 견뎌낼수있는광염성이며(Sampaio et al., 2003),
염분변화에 노출되었을때이온조성과삼투질농도를일정하게유지할수있으며
,
이온과삼투조절이일어나는동안에너지가소모된다(Nordlie, 2009; Lisboa et al., 2015).
어류의삼투조절에사용 되는에너지는10-50%
이상으로(Boeuf and Payan, 2001),
삼 투조절에의해소비되는에너지는어류의체액과서식지의염 분간에삼투질농도차에비례하므로(Handeland et al., 1998),
어류의혈장삼투질농도와유사한염분의해수에서사육할경 우삼투조절에필요한에너지를줄일수있다(Febry and Lutz, 1987; Woo and Kelly, 1995).
대사율은해양생물에서유기적 으로에너지를소비하는모든생리적과정의총체이며,
삼투조 절과관련된에너지비용의예측은산소소비측정으로가능하 다(Anni et al., 2016).
그러므로어류양식에서삼투적균형유 지는생존,
성장및건강도등어류의양성관리를위해매우필대왕붉바리 치어(Epinephleus akaara♀×E. lanceolatus♂)의 생존, 성장 및 생리학적 반응에 미치는 염분의 영향
신윤경*·최영재·길현우
국립수산과학원 남해수산연구소
Survival, Growth and Physiological Response of the Juvenile Hybrid Grouper (Epinephleus akaara♀×E. lanceolatus♂) Exposed to Different Water Salinity Levels
Yun Kyung Shin*, Young Jae Choi and Hyun Woo Gil
Aquaculture Industry Research Division, South Sea Fisheries Research Institute, National Institute of Fisheries Science, Yeosu 59780, Korea
This study aimed to determine the appropriate salinity condition for optimal cultivation of the juvenile hybrid grou- per Epinephleus akaara ♀×E. lanceolatus ♂ in conditions of decreasing salinity. The survival, growth, metabolic rate, stress, and histological response were investigated in juvenile hybrid grouper exposed to different salinities for 30 days. At 0 psu, the survival rate of the juvenile hybrid grouper was 60% on the 2nd day of exposure and all individu- als died on the 3rd day of exposure. At salinities above 3 psu, all animals survived throughout the exposure period.
Growth rate for body weight of the juvenile hybrid grouper was the highest in the control and the lowest at 10 psu.
Plasma osmolality of the juvenile hybrid grouper exposed to different salinities was 313-355 mg Osmol/kg at salini- ties above 10 psu, and then decreased to 225-264 mg Osmol/kg at salinities below 5 psu. The oxygen consumption rate tended to decrease as the salinity decreased. Stress responses of the juvenile hybrid grouper were analyzed with decreasing salinity. Therefore, it is considered that a salinity of more than 20 psu is suitable for the culture of the juvenile hybrid grouper.
Keywords: Hybrid grouper, Salinity, Survival, Growth, Stress response
*Corresponding author: Tel: +82. 61. 690. 8971 Fax: +82. 61. 685. 9073 E-mail address: [email protected]
This is an Open Access article distributed under the terms of the Creative Commons Attribution Non-Commercial Licens (http://creativecommons.org/licenses/by-nc/3.0/) which permits unrestricted non-commercial use, distribution, and reproduction in any medium, provided the original work is properly cited.
Received 11 June 2021; Revised 12 July 2021; Accepted 20 August 2021 저자 직위: 신윤경(연구관), 최영재(연구사), 길현우(연구사) https://doi.org/10.5657/KFAS.2021.0426
Korean J Fish Aquat Sci 54(4), 426-433, August 2021
대왕붉바리 치어의 생리에 미치는 염분의 영향
427
요하다
.
또한교잡종의염분적응력과염분에따른생리적반응 과관련한국내정보는붉바리와대왕붉바리의성장,
생존및스 트레스에미치는염분의영향(Lim et al., 2016)
외매우미흡 한실정으로,
염분에의한스트레스를줄이고성장을증가시키 기위한양성기술의요소로서염분적응력에대한연구는반드 시필요하다.
따라서본연구는대왕붉바리의염분내성범위
,
염분변화에따 른대사율,
스트레스및조직학적반응을분석하여대왕붉바리 의적정양성을위한염분조건을구명하기위하여수행되었다.
재료 및 방법
실험에사용된대왕붉바리치어는
Golden Seed Project
사업 의일환으로전라남도무안군에위치한청솔수산에서2019
년8
월에생산한개체들을2020
년1
월에국립수산과학원남해수 산연구소생물사육실(Yeosu, Korea)
로옮긴후3
톤유수식수 조에서수온26±1°C,
염분34.1±0.5 psu (
일반해수),
용존산 소농도9.0±1.0 mg/L
를유지하며2
달간순치하였다.
순치기 간동안먹이는배합사료(Nova floating ∅3-4 mm; Aller aqua, Christiansfeld, Denmark)
를어체중의1%
씩오전8
시경,
오후1
시와6
시경으로나누어매일3
회씩공급하였다.
적정양성을 위한염분및하한염분의범위를분석하기위하여실험수온은26±0.5°C,
실험염분은0 (
지하수), 3, 5, 10, 15, 20, 25±0.5 psu
및일반해수(
대조구, 34.1±0.5 psu)
였다.
각염분별로450 L
수조에서실험어50
마리씩수용하여30
일동안생존율,
성장 및생리학적반응을조사하였다.
실험은같은방법으로2
회반 복하였다.
실험수온은아쿠아트론(You won engineering Co., Ulsan, Korea)
을이용하여유지하였으며,
염분은지하수와해 수를혼합하여실험염분을맞추면서유수식으로수행하였다.
실험염분은 수질측정기(Professional plus; YSI Inc., Yellow Springs, OH, USA)
를사용하여매일오전,
오후1
회씩하루2
회점검하였다.
실험에는800
개체를사용하였으며,
개체의크 기는평균전장10.2±0.6 cm,
평균체중량19.5±3.1 g
이었다.
생존율은오전,
오후8
시경에12
시간간격으로점검하여사망 개체를선별하여생존율로 환산하여구하였으며,
사망개체는 실험어류가뒤집히고아가미뚜껑의운동이정지되었을때사 망으로판정하였다.
실험어의성장률은각실험구별생존한개체를대상으로전장 과체중을측정하여아래와같은공식에따라구하였다
.
전장은digital vernier calipers
를사용하여0.1 mm
단위로계측하였고,
체중은전자저울을사용하여0.01 g
까지측정하였다.
전장성장률
(growth rate for total length, %)=(
실험종료시 전장-
실험개시시전장)/(
실험개시시전장)×100
체중성장률
(growth rate for body weight, %)=(
실험종료시 체중-
실험개시시체중)/(
실험개시시체중)×100
염분변화에따른생리학적반응은각실험염분에노출시킨후
30
일째살아있는개체를대상으로산소소비율,
간과신장의조 직내SOD (superoxide dismutase), CAT (catalase),
코티졸, GSH (glutathione)
의변화,
혈액내삼투질농도및아가미의조 직학적반응을분석하였다.
산소소비율은respirometer cham- ber (volume, 2 L)
와산소호흡측정기(Orbis 3600; Orbis, Zu- rich, Switzerland)
를이용하여실험전후respirometer cham- ber
내용존산소의차로서구하였다.
삼투질농도는삼투압측정 기(OSMOMET 030; Gonotec, Berlin, Germany)
를이용하였 다.
간과신장의조직내코티졸, GSH, SOD
및CAT
반응은조직을분쇄하여
10,000 rpm
으로원심분리한후세포상층액을추출하여
-80°C
에보관하여분석에사용하였다. SOD, CAT,
코 티졸은어류ELISA kit (Cusabio Biotech, Wuhan, China)
를 사용하였으며, Versamax SoftMax pro 6 (Morecular Devices, San Jose, CA, USA)
를이용하여SOD
와코티졸은파장450 nm, CAT
는파장520 nm
에서분석하였다.
조직내GSH
분석은cold PBS containing 1 mM EDTA
에서조직을분쇄하여15
분 간4°C
에서10,000 rpm
으로원심분리하여Kinetic Method
를 이용하여5
분간격으로30
분간405-414 nm
파장으로분석하 였다.
간의조직학적분석은조직을적출하여Bouin's solution
에24
시간동안고정하고, 24-36
시간동안흐르는물에서수세한다음
70-100%
에탄올을이용하여단계적으로탈수하였다.
이후
paraplast (McCormick, Baltimore, MD, USA)
을이용하 여포매하고, microtome (RM2235; Leica, Wetzlar, Germany)
을이용하여두께
4-6 μm
의횡단면과종단면의연속절편을제작하였다
.
제작된조직절편은Mayer's hematoxylin-eosin (H- E)
비교염색하여광학현미경(BX50; Olympus, Tokyo, Japan)
으로관찰하였다.
실험결과에사용된유의성검정은SPSS 19.0
을이용하여one-way ANOVA
로통계처리후Duncan̓s mul- tiple range test
로사후검증하였다.
결 과
염분별 생존율과 성장
대왕붉바리치어의생존율은
0 psu
노출의경우2
일째60%
였으며노출
3
일째모두폐사하였다(Fig. 1).
반면3 psu
이상 염분실험구에서는노출30
일동안모두생존하였다(Fig. 1).
각실험염분에노출시킨대왕붉바리의전장은
10.2±0.6 mm
에서30
일간노출시킨후11.6-12.9 mm
으로염분변화에따른 유의한차이를보이지않았다(P>0.05, Table 1).
반면체중량 은19.5±3.1 g
에서노출30
일후염분10 psu
에서31.6±7.5
g, 20 psu
에서40.9±11.7 g,
그리고 대조구(34.1 psu)
에서44.4±10.5 g
이었다.
체중성장률은대조구에서128.7%
로모 든염분실험구에비해유의하게높았으며(P<0.05), 20 psu
에서109.4%, 25 psu
에서91.4%, 15 psu
에서80.1%
그리고10 psu
에서62.0%
으로체중성장은10 psu
에서가장낮았다(Table 1).
신윤경
ㆍ
최영재ㆍ
길현우428
염분변화에 따른 생리학적 반응 삼투질농도 변화
염분에따른대왕붉바리치어의혈액삼투질농도는
Table 2
와같으며,
염분10, 15, 20, 25
및대조구(34.1 psu)
에노출시 킨경우313-355 mOsmol/kg
으로대조구와유의한차이를보 이지않았다(P>0.05).
반면, 0, 3, 5 psu
염분노출구의경우대 왕붉바리치어의삼투질농도는225-264 mOsmol/kg
으로대조구와
10 psu
이상의모든실험구에비해유의하게감소하였다
(P<0.05).
염분별 산소소비율 변화와 스트레스 반응
대왕붉바리 치어의 염분변화에 따른 산소소비율의 변화를
Fig. 2
에나타내었다.
염분3 psu
와5 psu
에서산소소비율은각 각62.9±11.4 mg O
2/kg fish/h
와63.7±12.7 mg O
2/kg fish/h
으로유사하였으며(P>0.05),
염분15 psu
와20 psu
에서각각77.4±19.6
및82.3±13.4 mg O
2/kg fish/h
으로유의한차이가 없었다(P>0.05).
그리고25 psu
에서는93.4±13.97 mg O
2/kg fish/h
으로대조구(34.1 psu)
에서96.6±13.97 mg O
2/kg fish/h
와유사하였다(P>0.05),
전체적으로염분이감소할수록대왕붉Table 1. Total body length (mm) and total wet body weight (g) in the juvenile hybrid grouper Epinephleus akaara♀×E. lanceolatus♂ exposed to different experimental salinities for 30 days
Parameter Salinity (psu)
10 15 20 25 control
Initial total length (mm/fish) 10.2±0.6
Final total length (mm/fish) 11.6±0.8 12.1±0.8 12.6±1.1 12.4±0.9 12.9±0.9
1GRL (%) 13.6±0.1 18.5±0.1 23.4±0.2 21.4±0.1 26.9±0.2
Initial body weight (g/fish) 19.5±3.1
Final body weight (g/fish) 31.6±7.5 35.2±8.1 40.9±11.7 37.4±9.1 44.6±9.7
2GRW (%) 62.0±2.2 80.1±2.5 109.4±4.3 91.4±2.9 128.6±4.6
1GRL (growth rate for total length,%)=(final total length- initial total length)/initial total length×100. 2GRW (growth rate for body weight,
%)=(final body weight-initial body weight)/initial body weight×100. Data are expressed as mean±SE (n=50).
0 20 40
0 5 10 15 20 25 30
Survival rate (%)
Exposure time (Day)
0 psu 3 psu 5 psu 10 psu
15 psu 20 psu 25 psu control
a a ab b b c c
0 20 40 60 80 100 120 140
3 5 10 15 20 25 control
Oxygen consumption rate (mg O
2/kg fish h)
Salinity (psu)
Fig. 2. Changes in oxygen consumption rate of the juvenile hybrid grouper Epinephleus akaara♀×E. lanceolatus♂ with decreasing of salinity. Different letters above each column indicate significant differences (P<0.05) between salinities, according to Duncan’s multiple range test.
0 20 40 60 80 100
0 5 10 15 20 25 30
Survival rate (%)
Exposure time (Day)
0 psu 3 psu 5 psu 10 psu
15 psu 20 psu 25 psu control
a a ab b b c c
0 20 40 60 80 100 120 140
3 5 10 15 20 25 control
Oxygen consumption rate (mg O
2/kg fish h)
Salinity (psu)
Fig. 1. Survival rate (%) of the juvenile hybrid grouper Epinephle- us akaara♀×E. lanceolatus♂ exposed to different salinities for 30 days.
Table 2. Plasma osmolarity (mOsmol/kg) in the juvenile hybrid grouper Epinephleus akaara♀×E. lanceolatus♂ exposed to different experi- mental salinities
Parameter Salinity (psu)
0 3 5 10 15 20 25 control
Plasma osmolality (mOsmol/kg) 225±0.7 233±30.4 264±5.6 355±2.1 313±1.4 334±0.7 350±2.1 344±7.7
Sea water osmolality (mOsmol/kg) 2 101 158 305 442 576 719 981
Plasma osmolality in 0 psu was sampled on survival. Data are expressed as mean±SE (n=5).
대왕붉바리 치어의 생리에 미치는 염분의 영향
429
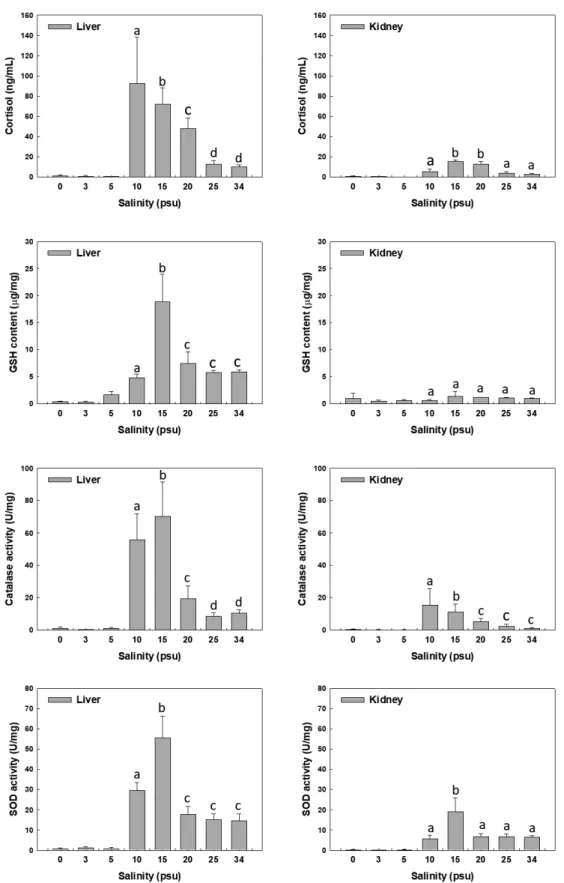
Fig. 3. Changes of cortisol, GSH, SOD and CAT in liver and kidney of the juvenile hybrid grouper Epinephleus akaara♀×E. lanceolatus♂
with decreasing of salinity. Different letters above each column indicate significant differences (P<0.05) between salinities, according to Duncan’s multiple range test. GSH, glutathione; SOD, superoxide dismutase.
바리치어의산소소비율은감소하는경향을보였으며
,
대조구 에서산소소비율이가장높았다(Fig. 2).
염분별
30
일동안노출시킨후생존한개체를대상으로간과 신장에서코티졸, GSH, SOD
및CAT
반응을Fig. 3
에나타내 었다.
코티졸은간에서,
대조구의경우약10 ng/mL
의농도를 유지하지만염분감소에따라급격히증가하는양상을나타냈 다.
특히염분20 psu
이하에서대조구에비해농도증가가유의 하게관찰되었다(P<0.05).
신장의경우에는15, 20 psu
노출시 코티졸이약간증가하는것이관찰되었으나유의한차이는없 었다(P>0.05). GSH
는간에서대조구의경우약6 μg/mg
의농 도를유지하지만15 psu
노출시유의하게증가하는것이관찰 되었다(P<0.05).
반면신장의경우염분감소에따라전체적으 로GSH
농도가다소증가한것이관찰되었으나,
유의성은없었 다(P>0.05). CAT
는간에서대조구의경우약9 U/mg
의활성도 를유지하고, SOD
의경우13 U/mg
의활성도를유지하지만,
염 분감소에따라활성도가증가하는것이관찰되었다.
특히10, 15 psu
노출시간에서SOD
와CAT
활성도가급격히증가하였 다(P<0.05).
전반적으로염분5 psu
이하에서간과신장에서코 티졸, GSH, SOD
및CAT
반응활성도가나타나지않았으며,
염분10 psu
와15 psu
에서는유의하게급격히증가하는경향을보였으며
, 25 psu
이상에서는대조구와유사한상태를유지하는것으로나타났다
(Fig. 3).
염분에따른대왕붉바리치어아가미의조직학적변화를
Fig.
4
에나타내었다.
대조구에서아가미는 새판의규칙적인 배열 등을비롯한정상적인조직상을보였다(Fig. 4A).
하지만, 25 psu
와20 psu
에노출된개체의아가미에서는일부염세포와점 액세포의활성과변성이확인되었으며, 20 psu
에서는새판상 피층의비후와모세혈관의충혈이나타났다(Fig. 4B and 4C).
15 psu
실험구개체의아가미에서는새판상피층의비후,
염세포세포질의호산성염색성증가및점액세포의공포화등조직 학적변성이뚜렷하였다
(Fig. 4D). 10 psu
이하의실험구에서 는염세포의붕괴와점액세포의소실(Fig. 4E and 4F),
벽주세 포의붕괴에의한모세혈관의융합,
새판상피층의소실(Fig. 4
F and 4G)
등염분농도가감소함에따라조직학적변성이증가되는경향을보였다
.
고 찰
염분은수서생물의생리에직접적으로영향을미치므로 생 태학적으로중요한환경요인중하나이다
(Urbina and Glover,
2015).
이는환경과수서동물간의삼투적차이가수서생물의생리적과정에영향을미치며
,
폐사를일으킬수있음을의미한 다(Edwards and Marshall, 2013).
또한염분변화에따른어류 의삼투조절능력은해양환경에서어류의생존과성장을예측할 수있는지표이다(Stewart et al., 2016)
본연구에서30
일동안 각실험염분에노출시킨대왕붉바리치어는0 psu
에서72
시간 째모두폐사하였으며폐사직전삼투질농도는225 mOsmol/
kg
이었다.
반면3 psu
이상에서30
일간모두생존하여대왕붉바리치어가생존이가능한염분의한계는
3 psu
이상으로분석되었다
.
반면, Lim et al. (2016)
에따르면대왕붉바리치어(
평균 전중10.0±0.4 g)
의경우염분8 psu
에서40
일노출기간동안 생존율이90%
였으며,
붉바리는8 psu
에서전량폐사하여대왕 붉바리가붉바리에비해염분내성이강한것으로보고하였다.
또한바리류(Epinephelus spp.)
는광염성으로염분10-33 psu,
수온22-28°C (Chen et al., 2016)
에서서식할수있는것으로알 려져있고,
붉바리(Epinephelus akaara)
는염분5 psu
에서는폐 Fig. 4. Histological changes of gill with decreasing of salinity during 30 days of the hybrid grouper Epinephleus akaara♀×E. lanceolatus♂.A, control (34.1 psu). ×400; B, 25 psu, arrowhead: activated chloride cell. ×400; C, 20 psu, arrowhead: vacuolated mucous cell, circle:
hypertrophy of epithelial cell. ×400; D, 15 psu, arrowhead: vacuolated mucous cell, circle: degeneration of chloride cell. ×200; E, 10 psu, arrowhead: destruction of chloride cell. ×200; F, 5 psu, circle: fusion of capillary. ×200; G, 3 psu. circle: destruction of epithelial layer. ×200;
C, capillary; Cc, chloride cell; El, epithelial layer; Gf, gill filament; Gl, gill lamella; Hc, hemocyte; Mc, mucous cell; Pc, pillar cell.
대왕붉바리 치어의 생리에 미치는 염분의 영향
431
사하지않으며
, 3 psu
에서반수치사시간[LT
50(h)]
은31.3
시간 으로보고(Wu and Woo, 1983)
되고있다.
따라서위에보고된 바리류가광염성종임을감안한다면본연구에서붉바리암컷 과대왕바리수컷의교잡종인대왕붉바리치어는염분3 psu
이 상의염분에서30
일노출기간동안모두생존하여생존하한염 분은3-5 psu
으로분석된다.
염분변화는
ROS (reactive oxygen species)
생성을유발시키 는산화스트레스요인이며,
호르몬과식세포작용또는면역세 포분화와같은세포면역에도영향을미친다(Dominguez et al., 2005; Lushckak, 2011; Yin et al., 2011).
또한간의효소활성은 스트레스의크기,
오염원과질병을판단할수있는생물지표이 며,
어류의건강상태를나타낼수있다(Wagner and Congleton, 2004).
본연구에서대왕붉바리치어의간과신장의코티졸
, GSH,
SOD, CAT
에대한산화스트레스반응결과간에서코티졸은염분감소에따라유의하게증가하였다
. GSH
는간조직15 psu
에서급격히증가하였으나10 psu
이하의실험구에서유의하 게감소하였으며, SOD
와CAT
는15 psu, 10 psu
에서유의하 게증가하였다.
반면신장의경우, GSH
는유의한반응을보이 지않았으며, SOD
와CAT
는염분15 psu
와10 psu
에서증가하 였다.
염분감소에따른SOD
와CAT
활성의증가는Acipenser naccarii
에서 보고되고 있으며(Álvarez and Nicieza, 2005), yellowfin fish sea bream Acanthopagrus latus
와Asian seabass Lates calcarifer
은염분48 psu
에서6 psu
로감소함에따라코 티졸이증가하였다(Mozanzadeh et al., 2021).
이러한결과는 본연구의대왕붉바리치어와유사한결과를나타내었다.
반 면sablefish Anoplopoma fimbria (Kim et al., 2017)
와golden pompano Trachinotus ovatus (Ma et al., 2016)
에서는염분감소에따른
SOD
와CAT
활성이감소하여염분변화에따라종간에다양한변화를나타내었다
.
이러한염분변화에따라나타 나는산화스트레스반응들의다양한결과들은종특이성을가 지며,
어류의발달단계및적응기간,
실험조건등에따라차이(Mozanzadeh et al., 2021)
가있는것으로추정된다.
또한염분에따른아가미의조직학적반응은
15 psu
이하에서새판상피층의비후
,
염세포및점액세포의공포화등변성이뚜렷하여(Anni et al., 2016)
염분감소에따라조직학적변성이증가되는 경향을보였다.
이러한현상은저염분에서장기간사육할경우 염분이만성스트레스요인으로작용하여성장및대사장애를 일으키는요인이될수있을것으로여겨진다.
대왕붉바리치어의삼투질농도는대조구에서
344 mOsmol/
kg
이고염분10 psu
이상에서는313-355 mOsmol/kg, 5 psu
이하에서는225-264 mOsmol/kg
이었다.
전갱이류인Trachi- notus marginatus
의 혈장 삼투질농도는 염분4-20 psu
에서351.1±2.3-365.5±5.4 mOsmol/kg
이었다(Anni et al., 2016).
붉바리
Epinephelus akaara
의경우염분30 psu
에서20, 12, 7 psu
으로감소시킨경우노출시간에따라혈장내삼투질농도는300-500 mOsmol/kg
의범위에서변화를보였다(Woo and Wu, 1982).
한편black sea bream Mylio macrocephalus (Woo and Wu, 1982)
의 경우에는 염분3-30 psu
의 범위에서 삼투질농 도320-380 mOsmol/kg
을보고하였다.
일반적으로해산어류 의혈장내삼투질농도는서식지보다낮게유지되며, 400 mOs- mol/kg
이하를보고(Nordlie, 2009)
하고있어본연구에서염 분10-34.1 psu
에노출시킨대왕붉바리치어의혈장삼투질농 도는해산어류의일반적인삼투질농도특성을나타내고있다.
어류가염분에순치하는동안아가미의구조적,
생화학적및 생리적 변화가 발생하는 과정에서 에너지가 요구되며,
어류 가 삼투조절에 사용되는 에너지는 총에너지의10-50%
이다(Boeuf and Payan, 2001).
그러므로어류가이온조절또는삼투 조절에사용되는에너지의차이는어류의혈장과환경간에나타나는삼투적인농도차에의존하는것으로보고된다
(Hande-
land et al., 1998).
이는어류의혈장과유사한삼투질농도를가 지는염분에서사육한어류는삼투조절이나이온조절에사용되 는에너지가적어성장과다른대사과정에에너지를재분배할 수있음을의미한다(Anni et al., 2016).
본연구에서대왕붉바 리치어의산소소비율은염분감소에따라감소하는경향을보 였으며,
대조구(
자연해수, 34.1 psu)
에서유의하게높았고, 15 psu
이하에서유의하게낮았다.
체중증가에따른성장률은대 조구(
자연해수, 34.1 psu)
에서128.7%
로가장높았고, 15 psu
와20 psu
에서각각101.0%, 109.7%
으로유사하여대왕붉바 리의성장률은대조구를제외하고체액삼투질농도가유사한 해수에서성장률이높았으며,
이는sea bream Sparus sarba
의 삼투질농도가유사한해수에서높은성장률을(Woo and Kelly,
1995)
보인결과와유사하다.
그러나본연구에서대왕붉바리치어의성장률이대조구
(
자연해수, 34.1 psu)
에서가장높았으 며,
이는산소소비율이높고,
삼투질농도차에의한에너지비용 이높다(Moser and Hettler, 1989; Woo and Kelly, 1995)
는점 을감안한다면상반된결과를보여주고있다.
일반적으로경골 어류에서염분내성에대한증가는연령에의존(Zydlewski and McCormick, 1997)
하거나크기에의존(Farmer et al., 1978;
Nordlie et al., 1982)
하는경향이있다.
본연구에서대조구에 서대왕붉바리치어의성장률및산소소비율증가등생리적활 성도증가는부화하여자치어시기부터자연해수에순치되어성 장해온결과로보이며,
이러한결과로부터삼투조절에이용되 는에너지의량은어류성장을증가시키는데충분한요인은아 닌것(Lisboa et al., 2015)
으로여겨진다.
따라서본연구결과를종합하여보면염분
3-15 psu
가대왕붉 바리치어의생존에영향을미치지않는다할지라도산소소비 율의현저한감소,
산화스트레스효소의유의한반응및저염분 에기인한아가미의조직학적변성등으로인해장기간양성할 경우다양한생리적기능장애가발생할것으로판단되며,
양성 사육관리를위해염분20 psu
이상이적합할것으로여겨진다.
사 사
본 연구는
2021
년도 국립수산과학원 수산과학연구사업(R2021007)
의지원으로수행되었습니다.
References
Álvarez D and Nicieza AG. 2005. Is metabolic rate a reliable predictor of growth and survival of brown trout Salmo trutta in the wild?. Can J Fish Aquat Sci 62, 643-649. https://doi.
org/10.1139/f04-223.
AnniI SA, Biancjini A, Barcarolli IF, Junior ASV, Robaldo RB, Tesser MB and Sampaio LA. 2016. Salinity influ- ence on growth, osmoregulation and energy turnover in juvenile pompano Trachinotus mrginatus Cuvier 1832.
Aquaculture 455, 63-72. https://doi.org/10.1016/j.aquacul- ture.2016.01.010.
Boeuf G and Payan P. 2001. How should salinity influence fish growth?. Comp Biochem Physiol Part C Toxicol Pharmacol 130, 411-423. https://doi.org/10.1016/S1532- 0456(01)00268-X.
Chen TS, Wu YC and Chi SC. 2016. Decreasing salinity of sea- water moderates immune response and increases survival rate of giant groupers post betanodavirus infection. Fish Shellfish Immuno 57, 325-334. https://doi.org/10.1016/j.
fsi.2016.08.050.
Dominguez M, Takemura A and Tsuchiya M. 2005. Effects of changes in environmental factors on the non-specific im- mune response of nile tilapia Oreochro misniloticus L.
Aquac Res 36, 391-397. https://doi.org/10.1111/j.1365- 2109.2005.01220.x.
Edwards SL and Marshall WS. 2013. Principles and patterns of osmoregulation and euryhalinity in fishes. In: Euryhaline fishes, fish physiology 32. McComick SD, Farrel AP, Braun- er CJ, eds. Academic Press, Cambridge, MA, U.S.A., 1-44.
https://doi.org/10.1016/B978-0-12-396951-4.00001-3.
Farmer G, Ritter J and Ashfield D. 1978. Seawater adaptation and parr-smolt transformation of juvenile atlantic salmon
Salmo salar. J Fish Res Board Can 35, 93-100. https://doi.
org/10.1139/f78-013.
Febry R and Lutz P. 1987. Energy partitioning fish: the activity related cost of osmoregulation in euryhaline cichlid. J Exp Biol 128, 63-85. https://doi.org/10.1242/jeb.128.1.63.
Handeland SO, Berge A, Bjornsson BT and Stefansson SO.
1998. Effect of temperature and salinity on osmoregulation and growth of atlantic salmon (Salmo salar L.) smolts in sea water. Aquaculture 168, 289-302. https://doi.org/10.1016/
S0044-8486(98)00356-1.
Kim JH, Park HJ, Kim KW, Hwang IK, Kim DH, Oh CW and Kang JC. 2017. Growth performance, oxidative stress, and non-specific immune responses in juvenile sablefish Ano-
plopoma fimbria by changes of water temperature and sa-
linity. Fish Physiol Biochem 43, 1421-1431. https://doi.
org/10.1007/s10695-017-0382-z.
Lim SG, Han SB and Lim HK. 2016. Effects of salinity on the growth, survival and stress responses of red spotted grou- per Epinephleus akaara and hybrid grouper Epinephleus
akaara♀×E. lanceolatus♂. Korean J Fish Aquat Sci 49,
612-619. https://doi.org/10.5657/KFAS.2016.0612.Lisboa V, Barcarolli IF, Sampaio LA and Bainchini A. 2015.
Effect of salinity on survival, growth and biochemical pa- rameters in juvenile Lebranche mullet Mugilliza (Perci- formes: Mulilidae). Neotrop Ichthyol 13, 447-452. https://
doi.org/10.1590/1982-0224-20140122.
Luschak VI. 2011. Environmentally induced oxidative stress in aquatic animals. Aquat Toxicol 101, 13-30. https://doi.
org/10.1016/j.aquatox.2010.10.006.
Ma Z., Zheng P, Guo H, Jiang S, Qin JG, Zhang D and Liu X. 2016. Salinity regular antioxidant enzyme and Na+ K+ -ATPase activities of juvenile golden pompano Trachinotus
ovatus (Linnaeus 1758). Aquac Res 47, 1481-1487. https://
doi.org/10.1111/are.12606.
Moser ML and Hettler WF. 1989. Routine metabolism of juve- nile spot Leiostomus xanthurus (Lacépède) as a function of temperature, salinity and weight. J Fish Biol 35, 703-707.
https://doi.org/10.1111/j.1095-8649.1989.tb03021.x.
Mozanzadeh MT, Safari O, Oosooli R, Mehrjooyan S, Najafaba- di MZ, Hoseini SJ, Saghavi H and Monem J. 2021. The ef- fect of salinity on growth performance, digestive and antiox- idant enzymes, humoral immunity and stress indices in two euryhaline fish species: Yellowfin seabream Acanthopagrus
latus and Asian seabass Latescalcarifer. Aquaculture 534,
736329. https://doi.org/10.1016/j.aquaculture.2020.736329.Nordlie FG. 2009. Environmental influences on regulation of blood plasma/serum components in teleost fishes: a review.
Rev Fish Biol Fish 19, 481-564. https://doi.org/10.1007/
s11160-009-9131-4.
Nordlie FG, Szelistowski WA and Nordlie WC. 1982. On- togenesis of osmotic regulationin the striped mullet,
Mugil cephalus L. J Fish Biol 20, 79-86. https://doi.
org/10.1111/j.1095-8649.1982.tb03896.x.
Sampaio LA, Tesser MB and Burkert D. 2003. Toleerância de juvenis do pampo Trachinotus marginatus (Teleostei, Ca- rangidae) ao choque agudo de salinida de em laboratório.
Cienc Rural 33, 757-761. https://doi.org/10.1590/S0103- 84782003000400027.
Stewart HA, Noakes DLG, Cogliati KM, Peterson JT, Iver- son MH and Schreck CB. 2016. Salinity effects on plasma ion levels, cortisol and osmolality in chinook salmon fol- lowing lethal sampling. Comp Biochem Physio Part A Mol Integr Physiol 192, 38-43. https://doi.org/10.1016/j.
cbpa.2015.11.011.
Urbina MA and Glover CN. 2015. Effects of salinity on osmo- regulation, metabolism and nitrogen excretion in the amphi-
대왕붉바리 치어의 생리에 미치는 염분의 영향
433
dromous fish Galaxias maculatus. J Exp Mar Biol Ecol 473, 7-15. https://doi.org/10.1016/j.jembe.2015.07.014.
Wagner T and Congleton JL. 2004. Blood chemistry correlates of nutritional condition, tissue damage and stress in migra- tion juvenile chinook salmon Oncorhynchus tshawitscha.
Can J Fish Aquat Sci 61, 1066-1074. https://doi.org/10.1139/
f04-050.
Woo NYS and Kelly SP. 1995. Effects of salinity and nutrition- al status on growth and metabolism of Sparus sarba in a closed seawater system. Aquaculture 135, 229-238. https://
doi.org/10.1016/0044-8486(95)01003-3.
Woo NYS and Wu RSS. 1982. Metabolic and osmoregulatory changes in response to reduced salinities in the red grou- per Epinephelus akaara (Temminck & Schlegel), and the black sea bream Mylio microcephalus (Basilewsky). J Exp Mar Biol Ecol 65, 139-161. https://doi.org/10.1016/0022- 0981(82)90041-7.
Wu RSS and Woo NYS. 1983. Tolerance of hypo-osmotic sa- linities in thirteen species of adult marine fish: Implications for estuarine fish culture. Aquaculture 32, 175-181. https://
doi.org/10.1016/0044-8486(83)90279-X.
Yin F, Peng S, Sun P and Shi Z. 2011. Effects of low salinity on antioxidant enzymes activities in kidney and muscle of juvenile silver pomfret Pampus argenteus. Acta Ecol Sin 31, 55-60. https://doi.org/10.1016/j.chnaes.2010.11.009.
Zydlewski J and McCormick SD. 1997. The ontogeny of salin- ity tolerance in the American shad Alosa sapidissima. Can J Fish Aquat Sci 54, 182-189. https://doi.org/10.1139/f96- 251.