www.earticle.net
돼지 체외성숙란 및 수정란에서 미토콘드리아 특이적 Rhod-2 발현 양상 확인
제갈호근· 박효진 · 김진우 · 구덕본* 대구대학교 공과대학 생명공학과
Confirmation of Mitochondrial Specific Rhod-2 Expression Patterns in Matured and Fertilized Porcine Oocytes In Vitro
Ho-Geun Jegal, Hyo-Jin Park, Jin-Woo Kim and Deog-Bon Koo*
Department of Biotechnology, College of Engineering, Daegu University, Gyeongsan 38453, Korea
ABSTRACT1)
Mitochondria plays an important role in regulating cytoplasmic calcium (Ca2+) homeostasis for ATP production during early embryonic development in mammals. Accumulation of mitochondrial Ca2+
induces mitochondrial dysfunctions. However, its relevance with change of mitochondrial Ca2+ in matured and fertilized oocytes has not been reported. The aim of this study was to identify the changes of mitochondrial Ca2+ level by using Rhod-2 staining analysis which is known as mitochondrial Ca2+ indicator in matured and fertilized oocytes in vitro. Here, we divided into two groups (Group 1:
G1, compact cumulus cells and homogeneous cytoplasm and Group 2: G2, non-compact cumulus cells and heterogeneous cytoplasm) according to the morphological difference of COCs. Meiotic maturation rate decreased in G2 compared to G1 (p<0.001). Average number of Rhod-2 spots in matured and fertilized oocytes was higher in G2 than in G1 (6 h after IVF: p<0.05). We also investigated the detailed expression pattern of Rhod-2 spots dividing it into four parts (n<10, 10≤n< 20, 20≤n<30, and 30≤n).
The ratio of fertilized oocytes with less than 10 Rhod-2 spots was higher in G1 (p<0.05). In addition, the Rhod-2 intensity in matured and fertilized oocytes in G2 were all higher than that in G1 (matured oocytes; p<0.001 and zygotes; p<0.05). Based on these results, we confirmed that morphological difference of immature oocytes was correlated with mitochondrial Ca2+ accumulation during porcine oocyte maturation and fertilization in vitro.
(Key words: Mitochondrial Ca2+, Rhod-2, Maturation, Fertilization, Porcine Oocytes)
* Corresponding author: Deog-Bon Koo, Department of Biotechnology, College of Engineering, Daegu University, Gyeongsan 38453, Korea. Tel: +82-53-850-6557, E-mail: [email protected]
This is an Open Access article distributed under the terms of the Creative Commons Attribution Non-Commercial License (http://creativecommons.org/licenses/by-nc-nd/3.0/deed.ko), which permits unrestricted non-commercial use, distribution, and reproduction in any medium, provided the original work is properly cited. The moral rights of the named author(s) have been asserted.
Ⅰ. 서론
체외배양 시스템을 통한 돼지 초기배 생산에서 난포란 또는 난모세포의 품질은 궁극적으로 배아 발달에 영향을 미치는 것으로 알려져 있다. 특히, 초기배 품질 및 발달능 력은 난포란의 성장 과정에서 난포 분화(Follicular
differentiation) 및 난자 발생(Oogenesis)을 통해 형태학적, 세포학적, 생화학적 및 분자적 과정을 거치며 이루어진다 (Kempisty et al., 2012). 이러한 난포란의 품질을 판단하기 위해서 현재까지 많은 연구가 이루어지고 있으며, 그 중에 서 형태학적 모습으로 판단하는 방법은 가장 일반적이며, 널리 사용되는 방법 중 하나이다(Jackowska et al., 2009).
[Provider:earticle] Download by IP 118.70.52.165 at Monday, December 20, 2021 7:48 PM
www.earticle.net
난모세포의 형태학적 품질은 난구세포 확장 여부와 난포 란의 세포질 내 균일한 지질 성분 및 분포정도를 기준으로 판단된다(Uyar et al., 2013). 이러한 돼지 난포란 및 난구 세포의 형태학적 품질은 초기배의 발달능에도 영향을 미 치는 것으로 알려져 있다(Ohlweiler et al., 2013). 돼지 난 모세포의 세포질에는 지방성분인 트리글리세라이드가 (Triglyceride)가 많이 존재하고 있으며, 이러한 지질 성분 들은 체외성숙을 통해 수정을 준비하는 난자의 세포질에 서 adenosine triphosphate(ATP)를 생산하는 에너지 대사 에 관여되기 때문에 중요한 역할을 하는 것으로 알려져 있 다(Yang et al. 2018). 세포질이 균일하고 난구세포의 분포 도가 좋은 난모세포는 체외성숙 동안 핵 성숙이 높게 나타 나며, 이후 체외수정을 통한 접합자 형성 및 배반포 단계 로의 배 발달에도 밀접한 관련이 있는 것으로 보고되고 있 다(Varghese et al,. 2011).
일반적으로 칼슘은 세포소기관인 소포체와 미토콘드리 아를 통해서 저장되거나 세포질로 방출되며 다양한 세포 반응을 일으키는 2차 메신저로 잘 알려져 있다. 이러한 칼 슘 이온은 포유동물의 수정과정에서 밀접한 관련성이 있 다고 보고되었다. 포유동물의 체외수정 과정 동안 난모세포 의 세포질 내 칼슘 증가는 칼슘 진동(calcium oscillation)을 일으키고 난모세포의 활성화(oocyte activation)를 유도하 며, 이후 유사분열 및 배아 발달 프로그램을 진행시키는데 중요한 역할을 하는 것으로 알려져 있다(Malcuit et al., 2006).
미토콘드리아는 세포질 내 칼슘 이온이 축적되는 것을 조절하는 완충장치로써 세포질 내 칼슘 항상성에 관여한 다고 보고되었다. 이러한 칼슘 항상성 유지 및 조절은 ATP 생산을 유도하여 에너지를 공급하는 미토콘드리아는 난포란의 성숙, 수정, 배아 발달에 있어서도 필수적인 역할 을 수행하는 기관으로도 알려져 있다(Wang et al., 2009).
특히, 생쥐의 체외수정 동안 미토콘드리아는 세포질의 칼 슘 이온을 미토콘드리아 내로 흡수함으로써 수정 시 정자에 의해 유발되는 세포질 내 칼슘 진동을 조절한다(Dumollard et al., 2006). 생쥐 난자에서 미토콘드리아 기능 억제는 정 자에 의해 유발된 칼슘 진동 패턴을 파괴함으로써 세포질 내 칼슘을 지속적으로 증가 시킨다고 보고된 바 있다(Liu et al., 2001). 한편, 세포질 내 칼슘증가로 인한 미토콘드리 아로의 과도한 칼슘축적은 미토콘드리아 기능 장애 (Mitochondrial dysfunction) 및 미토콘드리아 매개성 세포 죽음에 관련된 신호의 활성화를 유발한다(Santulli et al., 2015). 난모세포에서 미토콘드리아 기능 장애로 인한 발달 장애, 칼슘 과부하, 미토콘드리아 막 잠재력의 감소 및 미
토콘드리아의 DNA 손상은 난모세포의 품질 저하에 따른 여성 난임의 원인으로 알려져 있다(Schatten et al., 2014).
이러한 사실들은 미토콘드리아가 세포질의 칼슘 조절에 관여될 뿐만 아니라 미토콘드리아 내 칼슘 변화는 미토콘드 리아 기능 유지에도 중요하다는 것으로 설명된다(Babayev and Seli, 2015). 그러나, 현재까지 연구에서 돼지 난모세포 의 형태학적인 품질 차이로 인해 발달 과정에서의 미토콘 드리아 칼슘 변화와의 상관관계에 대해 보고된 바가 없다. 따라서 본 연구에서는 형태학적인 돼지 난모세포의 품 질 차이에 따른 돼지 성숙란 및 체외수정란에서 미토콘드 리아 내 칼슘 변화와의 연관성에 대해 확인하고자 하였다. 본 연구에서는 돼지 난모세포에서 형태학적인 측면 차이 로 나눈 두 그룹 (Group 1: G1 and Group 2: G2)으로부터 체외성숙란과 수정란으로부터 미토콘드리아 특이적 칼슘 측정에 사용되는 Rhod-2를 이용해 미토콘드리아 칼슘의 축적 양상을 비교 분석하고자 하였다.
Ⅱ. 재료 및 방법
1. 시약
본 연구에서 특별히 명시하지 않은 화학물질들은 Sigma- Aldrich Korea(Yongin, Korea)에서 구입하여 사용하였다.
2. 난소 채취 및 체외성숙
본 연구에서 사용된 돼지 난소는 지역의 도축장(대구축 산물도매시장 신흥산업(주))에서 채취하여 75㎍/ml potassium penicillin G 가 첨가된 0.9% 생리식염수가 담긴 30-35℃ 조건의 보온병에 넣어 실험실로 운반하였다. 운반 된 난소는 생리식염수로 3회 세척한 후 18 게이지 주사바 늘이 부착된 10ml 주사기를 이용하여 3-6mm 지름의 난포 에서 난포액을 채취하였다. 채취한 난포액을 petridish 에 옮긴 후 실체현미경 하에서 마우스피펫을 이용하여 선별 하였다. 이러한 과정에서 Group 1은 3-5개의 난구세포층 을 가진 난포란에서 세포질 함유 면적이 80% 이상인 난포 란만을 선택하였고, Group 2는 1-2개의 난구세포층과 80%
이하의 세포질을 가지는 난포란을 선택하여 두 그룹으로 분리하여(Warriach and Chohan, 2004; De Vico et al., 2005) 체외성숙을 수행하였다. 이후, 선별된 난포란을 tyrode's lactate-N-2-hydroxyethylpiperazine -ethanesulfonic
[Provider:earticle] Download by IP 118.70.52.165 at Monday, December 20, 2021 7:48 PM
www.earticle.net
acid(TL-HEPES) 용액으로 3회 세척한 후 최종적으로 체외 성숙 배지에서 세척한 다음 배양액 500㎕ 씩 분주된 4-well multi dish(NUNC, Roskilde, Denmark)에 well 당 40-50개 의 미성숙한 난포란을 넣고 38.5℃, 5% CO2조건하에서 성 숙을 유도하였다. 미성숙한 난포란의 체외성숙에 사용된 배지는 North Carolina State University-23(NCSU-23) 기 본 배지에 10% porcine follicular fluid, 10IU/ml human chorionic gonadotropin(hCG), 10IU/ml pregnant mare’s serum gonadotropin(PMSG), 10ng/ml epidermal growth factor(EGF), 10ng/ml β-mercaptoethanol 및 0.57mM cystein(Petters and Wells, 1993)을 첨가하여 사용하였다.
미성숙 난포란들을 22시간 동안 배양시킨 후, PMSG 와 hCG 가 첨가되지 않은 배양액에서 22시간 동안 추가 배양 하였다.
3. 체외수정
돼지 난포란의 체외수정은 Abeydeera와 Day(1997)의 방법을 토대로 일부 변형하여 수행하였다. 체외수정에 사 용된 배지는 113.1mM NaCl, 3mM KCl, 7.5mM CaCl2, 5mM sodium pyruvate, 11mM glucose, 20mM Tris, 2.5mM caffeine sodium benzoate과 1mg/ml BSA로 구성 된 modified Tris-buffered medium(mTBM)을 사용하였다.
신선한 액상 정액은 1주일에 2번 Darby Porcine AI Center(Anseong, Korea)로 부터 구입하였으며, 17℃를 유 지하며 보관하였다. 신선한 정액은 1mg/ml BSA, 100㎍
/ml penicillin G, 그리고 75㎍/ml streptomycin sulfate가 첨가된 PBS에서 원심분리기를 사용하여 3회 세척하였다.
이후, 1,500rpm에서 3분간 원심분리한 후 상등액을 제거하 고 Makler chamber를 이용해 정자수를 계산하였다. 체외 수정을 위한 최종정자의 농도는 1.5×105정자수/ml로 준비 하였다. 두 그룹의 성숙란은 각각 별도로 0.1%
hyaluronidase에서 부드럽게 피펫팅하여 난구세포를 제거 한 후, mTBM에서 3회 세척하였다. 체외수정 배양액을 이 용하여 그룹을 분리하여 60mm 배양접시에 48μl씩 소적 을 만든 후 10-15개의 성숙된 난포란을 넣고 준비된 정자 를 각각 2μl씩 첨가하여 38.5℃, 5% CO2 배양기에서 6시 간 동안 체외수정을 유도하였다.
4. 체외성숙란 및 수정란에서 Rhod-2의 염색
형태학적 품질 차이에 따라 분류한 난포란을 이용하여 체외성숙과 수정을 실시한 후 난자내의 미토콘드리아에서
칼슘을 확인하기 위하여 Rhod-2 AM(ab142780, Abcam, Cambridge, MA, UK)을 사용하여 염색을 실시하였다. 먼 저, 실시간으로 Rhod-2의 spot을 확인하기 위하여 체외성 숙이 완료된 성숙란과 수정 후 3시간 및 6시간째의 수정란 을 그룹별로 구분하여 0.1% PVA가 함유된 PBS에서 3회 세척하였다. 이후, 암전 상태에서 100μM Rhod-2가 첨가 된 체외수정 배양액에 38.5℃, 5% CO2배양기에서 15분 동 안 배양하였다. 이후 iRiStm digital cell imaging system (Logos Biosystems, Inc., Anyang, Korea)을 이용하여 Rhod-2 spot의 수를 확인하였다. 또한, 고정된 상태의 체 외성숙란과 수정란의 Rhod-2을 확인하기 위해 앞서 실시 간 상태에서 사용한 방법과 동일한 방법을 사용한 후 2.5%
glutaraldehyde가 함유된 PBS에서 1시간 동안 상온에서 고 정시켰다. 고정된 성숙란과 수정란을 0.1% PVA가 함유된 PBS 용액으로 5분간 3회 세척한 다음, slide glass에 옮긴 후 iRiStm digital cell imaging system(Logos Biosystems,,, Inc.)을 이용하여 Rhod-2의 intensity을 확인하였다.
5. 통계 분석
본 연구를 통해 얻은 체외성숙란 및 수정란에서 Rhod-2 spot의 분포 양상과 백분율 값은 평균±표준편차(S.D)로 나 타내었으며, t-test를 통해 모든 실험값을 분석하였다.
Image J(NIH, MD) software를 사용하여 이미지 데이터를 분석하였다. 본 연구에서 나타난 결과들에 대한 통계적 유 의차는 p<0.05 이하인 것만을 표기하였다.
Ⅲ. 결과
1. 형태학적 차이로 나눈 두 그룹에서 난구세포 확장, 세 포질 면적 및 핵 성숙율 비교
미성숙 돼지 난포란을 형태학적 측면을 기준으로 두 그 룹(G1 그리고 G2)으로 구분하여 체외성숙을 수행하였다.
G1은 여러 겹의 난구세포층이 고르게 잘 분포되어 있고 균일한 세포질을 가지는 미성숙 난포란 그룹이며, G2는 G1에 비해 상대적으로 난구세포층이 적고 고르지 못하며 세포질이 균일하지 못한 미성숙 난포란 그룹이다. 체외성 숙을 22시간 진행 후 확인한 결과, G1이 G2에 비해 난구세 포의 확장이 넓게 진행되었다. 44시간 동안의 체외성숙이 완료된 후, G1이 G2에 비해 난구세포의 확장이 뚜렷하게
[Provider:earticle] Download by IP 118.70.52.165 at Monday, December 20, 2021 7:48 PM
www.earticle.net
Fig. 1. Comparison of different two groups of porcine COCs according to the different cumulus cells expansion and cytoplasm area. (A) Porcine COCs at GV stage were classified by two groups according to; G1 had more than 80% cytoplasm and 3-5 cumulus cells layers, G2 had less than 80% cytoplasm and 1-2 cumulus cells layers. Morphology of each porcine COCs during IVM (0, 22 and 44 h) were visualized using an optical microscopy (Leica, EC3). Scale bar is 100 μm. (B) The mean percentage of cytoplasm area between G1 and G2 GV stage oocytes. Differences were considered significant at *p<0.05 as compared between G1 and G2.
Table 1. Comparison of meiotic maturation in porcine COCs between G1 and G2 Group No. of oocytes
examined
% of oocytes (n)
GV GVBD M I M II
G1 164 2.1±2.7 (5) 0.7±1.4 (1) 16.8±1.7 (28)a 80.4±3.5 (130)a
G2 188 5.9±2.3 (11) 2.9±2.6 (5) 45.6±3.7 (84)b 42.0±2.0 (78)b
a,bDifferent superscript letters denote a significant difference (p<0.001).
증가하였으며, 두 그룹간의 난구세포 확장 정도의 차이가 나타남을 확인할 수 있었다(Fig. 1A). 또한, 구분한 두 그룹 의 난포란에서 평균 세포질의 면적을 확인한 결과, G1에서 는 84.8±1.7%로 G2에서는 68.4±4.5%로 나타났다(p<0.05, Fig. 1B). 뿐만 아니라, 예상한 바와 같이 G1과 G2의 난포 란을 이용하여 44시간 체외배양을 실시한 후 orcein 염색 을 통해 핵 성숙율을 조사한 결과, G2의 난자에서 MII 단 계까지의 핵 성숙율이 G1에 비해서 유의적으로 감소하는 것을 확인하였다(p<0.001, Table 1). 이러한 형태학적 차이 를 보이는 두 그룹을 사용하여 다음 실험을 수행하였다. 2. 돼지 난포란의 품질차이에 따른 Rhod-2 spot 수의 분석
체외성숙란 및 수정란에서 발현되는 평균적인 Rhod-2 spot 수를 분석하였다. Rhod-2 spot 수의 평균은 각 그룹 별로 전체 Rhod-2 spot 수를 실험에 공시한 난모세포의 수
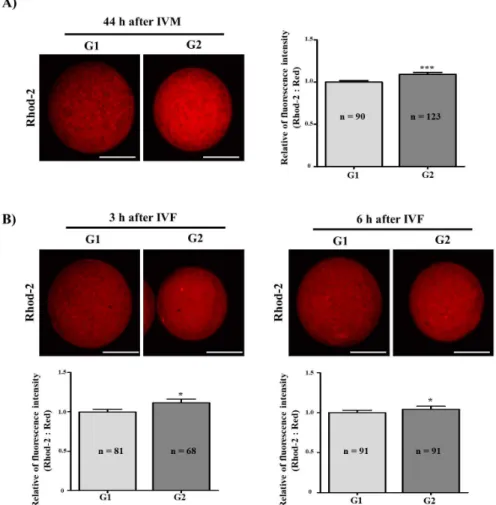
로 나누어 비교하였다. 우선적으로, G1과 G2 두 그룹의 44 시간째 체외성숙란에서 Rhod-2 spot 의 평균수로 비교했 을 때, G2의 성숙란에서 G1보다 평균적인 Rhod-2 spot 이 1.5배 유의적으로 증가하였다(G1:11.5±2.2 vs G2:16.7±4.1, Fold: 1.45, Fig. 2A). 체외수정 후 3시간과 6시간째 수정란 에서 동일한 방법으로 G1과 G2 두 그룹의 Rhod-2 spot 의 평균수를 비교하였다. 그 결과, 체외수정 후 3시간째에 G2 가 G1에 비해 1.3배 많은 Rhod-2 spot 수를 나타냈으나 두 그룹에서 유의적인 차이는 나지 않았다(G1: 11.0±2.5 vs G2: 14.3±2.2, Fold: 1.3, Fig. 2B). 체외수정 후 6시간째에서 는 Rhod-2 spot 의 평균수치가 G2에서 G1에 비해 1.6배정 도 가장 크게 증가하였다(G1: 6.7±2.4 vs G2: 10.6±0.1, Fold: 1.6; p<0.05 Fig. 2B). 이러한 결과를 통해 체외성숙란 및 체외수정 후 3시간과 6시간째의 수정란에서 평균적인 Rhod-2 spot 의 수가 G1보다 G2에서 증가함을 알 수 있었 다.
[Provider:earticle] Download by IP 118.70.52.165 at Monday, December 20, 2021 7:48 PM
www.earticle.net
Fig. 2. Comparison of Rhod-2 spot numbers in matured and fertilized porcine oocytes from G1 and G2 groups. Changes of Rhod-2 spots were compared with G1 and G2 at 44 h after IVM (A) and at 3 h or 6 h after IVF (B). Rhod-2 spots (Red) in two groups were showed by using a iRiSTM image system (Logos Biosystems). Data in the bar graph represent the means±SD from three independent experiments. Differences were considered significant at *p<0.05 as compared between G1 and G2.
Fig. 3. Investigation of detected Rhod-2 spots in matured and fertilized porcine oocytes from G1 and G2 groups. We divided to the 4 groups of detected Rhod-2 spots; n<10, 10≤n<20, 20≤n<30, and 30≤n. Changes of mitochondrial Ca2+ by using a specific mitochondrial staining dye Rhod-2 in matured porcine COCs of G1 and G2 at 44 h after IVM (A) and at 3 h or 6 h after IVF (B). Differences were considered significant at *p<0.05 as compared between G1 and G2.Scale bar is 100 μm.
3. 체외성숙란 및 수정란에서의 Rhod-2 발현 양상 분석
형태학적 차이를 보이는 두 그룹의 난포란들을 체외성 숙 및 수정 후 Rhod-2를 염색하여 spot 의 수를 네 개의 군 (n<10, 10≤n<20, 20≤n<30, 그리고 30≤n)으로 나누어 조 사하였다. 또한, Rhod-2에 의해 염색된 spot 수에 따라 구 별된 네 개의 군에 해당되는 난모세포를 비율로 분석하였 다. 체외성숙란은 44시간 성숙이 완료된 난모세포를 사용 했으며, 체외수정란은 수정동안 그룹 간 변화 차이를 확인 하기 위해서 수정 후 3시간과 6시간째 난모세포로 구분하
여 실험을 진행하였다. 우선적으로, 44시간 성숙이 완료된 난모세포에서 Rhod-2 spot 수에 따라 네 개의 군으로 분리 후, 각 군에 속하는 난모세포 비율을 조사하였다. 체외성숙 란의 경우, G1과 G2 그룹 모두 10개 보다 같거나 많고 20 개 보다 적은 Rhod-2 spot 수(10≤n<20)를 가지는 난포란 들이 주로 관찰되었다. 특히, G1에서는 Rhod-2 spot 이 20 개 보다 적은 군들에서 총 70개 중에서 61개가 속하여 87.1%의 난자 분포 비율을 보였으며, 이는 총 108개 중에 서 82개의 난자 분포 비율(75.9%)을 보인 G2 그룹 보다 높 은 것을 확인할 수 있었다(Fig. 3A).
[Provider:earticle] Download by IP 118.70.52.165 at Monday, December 20, 2021 7:48 PM
www.earticle.net
또한, 체외수정 3시간과 6시간째 수정란에서 G1과 G2의 Rhod-2 spot 수에 따른 수정란의 분포 비율을 비교하였다.
체외수정 후 3시간째와 6시간째 G1의 수정란에서는 Rhod-2 spot 수가 10개 보다 적은(n<10) 군에서 가장 많이 관찰되 었으며(3h; G1: 47.5% 그리고 6 h; G1: 69.8%), G2 그룹에 서는 유의적으로 감소하였다(p<0.05). 체외수정 후 6시간째 수정란에서는 G2 그룹의 Rhod-2 spot 수가 10개 이상이거 나 20미만의 군(10≤n<20)에 속하는 난자의 분포가 G1보 다 유의적으로 높았다(G1: 28.4% vs G2: 44.2%, p<0.05;
Fig. 3B). 체외성숙란 및 수정란에서 Rhod-2 spot 의 수에 따라 분리한 네 개의 군의 해당 난자의 비율을 확인한 결 과, 체외성숙란 및 수정란에서 spot 수가 많은 난자의 비율 이 상대적으로G1에 비해 G2에서 높게 나타났다.
4. 돼지 난포란 품질 차이에 따른 Rhod-2 형광 발현 조사
체외성숙란 및 수정란을 고정 후 Rhod-2의 붉은색 형광 발현에 대한 intensity 을 통해 그룹 간 발현 정도의 차이를 비교하였다. 44시간 체외성숙된 난모세포의 경우 G2가 G1 에 비해 유의적으로 높은 Rhod-2의 형광 발현 intensity 를 가지는 것을 확인하였다(Fig. 4A; p<0.001). 이후, 체외수정 후 3시간째와 6시간째에서도 G1과 G2 두 그룹의 Rhod-2 intensity 를 측정하였다. 체외수정 후 3시간째에서 G2가 G1에 비해 intensity 값이 유의적으로 높게 나타났으며, 6 시간째에서도 G2가 G1에 비해 유의적으로 높은 intensity 값을 나타냄을 확인할 수 있었다(Fig. 4B; p<0.05).
Fig. 4. Comparison of detected Rhod-2 intensity in matured and fertilized porcine oocytes from G1 and G2 groups. Measurement of Rhod-2 intensity was compared with G1 and G2 at 44 h after IVM (A) and at 3 h or 6 h after IVF (B). Histogram values of Rhod-2 intensity were obtained by using Image J (NIG, MD) software. Data in the bar graph represent the means±SEM from three independent experiments. Differences were considered significant at *p<0.05, ***p<0.001 as compared between G1 and G2. Scale bar is 100 μm.
[Provider:earticle] Download by IP 118.70.52.165 at Monday, December 20, 2021 7:48 PM
www.earticle.net
Ⅳ. 고찰
본 연구에서는 형태학적 측면에서 분류한 두 그룹의 미 성숙 난포란을 이용하여 체외성숙 후 품질 차이를 비교하 였다. 특히, 형태학적 기준으로 나눈 G1과 G2 두 그룹의 체외성숙란 및 수정란에서 미토콘드리아 특이적 칼슘 변 화를 Rhod-2 염색을 통해 spot 수와 intensity 값의 차이를 확인할 수 있었다.
Rhod-2는 세포질 또는 미토콘드리아내의 칼슘 지시자로 알려져 있다(Xu and Yang, 2017). 본 연구에서는 돼지 미 성숙 난포란을 형태학적 기준으로 나눈 G1과 G2 두 그룹 으로 생산된 체외성숙란 및 수정란을 이용하여 미토콘드 리아내의 칼슘을 확인하기 위해 Rhod-2 염색을 이용하여 확인하였다. 미토콘드리아의 기능 장애로 인해 발생하는 칼슘 과부하, 불임, 발달 장애 등은 난모세포의 품질과 관 련이 있다(Liu et al., 2001). 그러나 형태학적 난모세포의 품질에 따른 미토콘드리아 칼슘 변화에 대한 연구는 알려 진 바가 없었다. 따라서 본 연구에서는 체외성숙란 및 체 외수정란에서 Rhod-2 spot의 변화 양상을 확인하였다. 체 외성숙란 및 수정 후 3시간째 난자에서는 G2가 G1에 비해 Rhod-2 spot의 수가 증가하는 것을 확인하였으며, 수정 후 6시간째 난자에서는 G2가 G1에 비해 유의적으로 Rhod-2 spot이 많은 것을 확인하였다(p<0.05, Fig. 2). 또한, Rhod-2 spot을 구체적으로 분석하기 위해서 spot 수에 따른 4개의 군(n<10, 10≤n<20, 20≤n<30, 그리고 30≤n)으로 나누어 비교 분석하였다. 체외성숙 44시간째 성숙란에서는 상대적 으로 Rhod-2 spot 수가 10개 보다 적은 군에 분포하는 난 자 비율이 G2 보다 G1 그룹에서 높았으며, 20개 이상의 Rhod-2 spot 군에서는 G2 난자 분포 비율이 높았다. 체외 수정 후 3시간 및 6시간째에서 G1이 G2에 비해서 Rhod-2 spot 수가 10개 미만인 난자의 비율이 G1에서 유의적으로 높았다. 수정 후 6시간째에서 10개보다 많거나 20개 보다 적은 군에서 G2가 G1에 비해 상대적으로 높은 비율을 확 인할 수 있었다(Fig. 3). 또한, 난포란 및 수정란에서의 전 체 Rhod-2의 형광발현을 조사하기 위한 intensity 측정 결 과도 예상한 바와 같이 G2 그룹의 성숙된 난포란 및 수정 란에서 G1보다 유의적으로 높게 나타났다(Fig. 4). 이러한 결과는 난포란의 선별에 있어서 난구세포 및 세포질의 형 태학적 차이는 체외성숙란 및 체외수정란의 미토콘드리아 내 칼슘이온의 축적 양상과 밀접한 관련이 있다는 사실을 제안한다.
이전 연구에 따르면, 소의 동결 보존된 난모세포에서 정
상적인 난모세포보다 세포질 및 미토콘드리아 내 칼슘 축 적으로 인한 비정상적인 칼슘의 농도 변화는 초기배 발달 에서 수정능과 발달 능력을 저하시킨다고 보고되었다 (Wang et al., 2017). 생쥐의 허혈성 심부전에서 미토콘드 리아의 과도한 칼슘축적은 기능장애를 일으키고 ATP 생 산 저하 및 ROS 생성이 증가가 된다고 보고되었다 (Santulli et al., 2015). 또한, 미토콘드리아의 기능장애를 가지고 있는 난모세포에서는 ATP 생산 감소, 스핀들 형성, 염색체 분리 및 수정에 악영향을 미친다고 알려져 있으며, 이러한 난모세포는 낮은 품질을 가지는 것으로 보고되었 다(Liu et al., 2001). 게다가, 비만 쥐의 난모세포에서 비정 상적인 미토콘드리아 내 칼슘 축적은 미토콘드리아의 기 능장애를 일으키며, 세포사멸관련 유전자인 cytochrome-C, Caspase-3의 발현을 통해 세포사멸을 한다고 보고되었다 (Zhao et al., 2017). 최근 Guo 등(2018)은 돼지 난모세포에 서 미토콘드리아 내의 칼슘 분포를 Rhod-2로 확인하였다 고 보고하였다. 이러한 보고를 토대로 본 실험에서도 체외 성숙 돼지 난자에서 Rhod-2 염색을 통해 미토콘드리아 내 칼슘의 분포 양상을 확인하였다. 특히, 실험을 진행하기 전 에 미토콘드리아 분포를 보여주는 Mitotracker와 Rhod-2 가 일치하는 것을 확인하였다(data not shown). 따라서 본 연구에서는 형태학적 품질에 따라 핵 성숙율의 차이를 보 이는 두 그룹을 이용하여 Rhod-2의 발현 정도 및 양상의 차이점을 체외성숙란 및 체외수정란에서 확인할 수 있었 다. 이러한 연구 결과를 기반으로 미토콘드리아 내 칼슘 변화는 미토콘드리아 기능 및 세포 사멸과 밀접한 관련성 을 가지기 때문에 돼지 체외성숙란과 수정란에서 Rhod-2 의 발현은 난자의 품질 여부 및 초기배 발달능을 예측할 수 있는 지표로 사용될 수 있다고 생각한다.
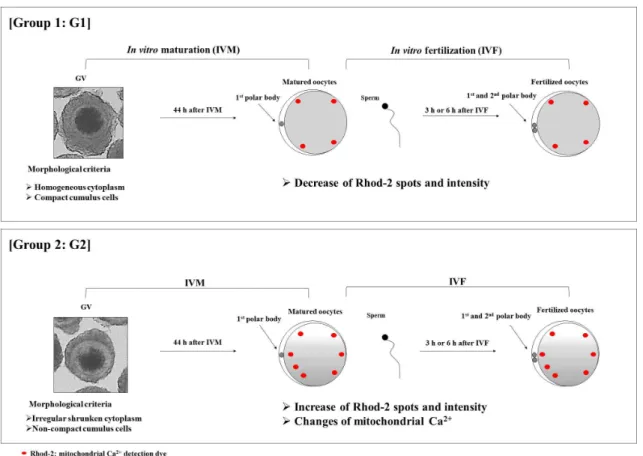
결론적으로 본 연구에서는 난구세포가 균일하고 세포질 이 고르게 분포하여 높은 핵 성숙율을 보인 G1이 G2에 비 해 체외성숙란 및 체외수정란에서 Rhod-2 spot의 수와 intensity 값이 감소함을 확인할 수 있었다(Fig. 5). 이는 성 숙란과 수정란에서 미토콘드리아 내 칼슘 축적이 G2 그룹 에 비해 G1 그룹에서 적게 나타난다는 것을 제시한다. 종 합해볼 때, 본 연구결과에서 미토콘드리아 칼슘 이온을 측 정하기 위해 사용한 Rhod-2의 spot의 수 및 intensity 변화 는 난포란의 형태학적인 품질 측면과 관련성이 있음을 확 인하였다. 따라서, Rhod-2 발현 양상을 통해 예측되는 미 토콘드리아 내 칼슘 변화는 형태학적 분석에 따른 돼지 난 포란의 체외성숙 및 체외수정에 있어서 품질 평가를 위한 지표로써 의미가 있다는 것을 제시한다.
[Provider:earticle] Download by IP 118.70.52.165 at Monday, December 20, 2021 7:48 PM
www.earticle.net
Fig. 5. Schematic diagram shows the difference of Rhod-2 expression pattern in matured and fertilized oocytes.
Ⅴ. 요약
미토콘드리아는 세포질 칼슘 항상성 및 ATP 생산에 중 요한 역할을 하는 세포 소기관으로 이러한 미토콘드리아 의 기능은 성숙과 수정 그리고 배 발달에 매우 중요한 역 할을 한다. 미토콘드리아 칼슘 축적은 기능장애를 일으킨 다. 그러나 돼지 체외성숙란 및 수정란에서 미토콘드리아 칼슘 변화의 관련성에 관한 연구는 보고된 적이 없다. 본 연구의 목적은 미토콘드리아 칼슘 지시자로 알려진 Rhod-2 염색을 이용하여 성숙란 및 수정란에서 미토콘드 리아 칼슘 축적의 변화를 확인하였다. 형태학적 모습의 기 준을 통해 난구세포의 세포층과 세포질의 균질도를 바탕 으로 G1과 G2로 나누어서 체외성숙을 진행하였다. 이후 두 그룹에서 핵 성숙율을 비교하였을 때, G2가 G1에 비해 낮게 나타났다(p<0.001). 돼지 체외성숙란 및 수정란에서 평균적인 Rhod-2 spot 의 수는 G1보다 G2에서 더 많이 나 타났다(6시간째 체외수정란: p<0.05). 다음으로 Rhod-2 spot 수에 따른 난모세포의 비율을 확인하기 위해 Rhod-2
spot 의 수를 4개의 군(n<10, 10≤n<20, 20≤n<30, 그리고 30≤n)으로 나누어 해당 난모세포의 비율을 확인하였다.
체외성숙란 및 체외수정란 모두 G1이 G2에 비해 10개 미 만(n<10)인 Rhod-2 spot 의 수를 가지는 난모세포가 많았 으며, 체외수정란에서는 유의적으로 높았다(p<0.05). 마지 막으로 체외성숙란 및 수정란에서 Rhod-2 intensity 값을 측정하여 두 그룹을 비교하였을 때, G2가 G1에 비해 유의 적으로 높은 것을 확인 할 수 있었다(성숙란; p<0.001 그리 고 수정란; p<0.05). 본 연구의 결과를 토대로 돼지에서 미 성숙 난포란의 형태학적인 품질은 체외성숙 및 체외수정 과정 동안 미토콘드리아 내 칼슘 축적과 관련이 있음을 확 인하였다.
사사
본 연구는 2017년도 대구대학교 교내연구비 과제에 의 해 수행되었음
[Provider:earticle] Download by IP 118.70.52.165 at Monday, December 20, 2021 7:48 PM
www.earticle.net
Ⅵ. 참고문헌
1. Abeydeera, L. R. and Day, B. N. 1997. In vitro penetration of pig oocytes in a modified Tris-buffered medium: effect of BSA, caffeine and calcium.
Theriogenology 48(4):537-544.
2. Babayev, E. and Seli, E. 2015. Oocyte mitochondrial function and reproduction. Curr. Opin. Obstet.
Gynecol. 27:175-181.
3. De Vico, G., Peretti, V. and Losa, G. A. 2005. Fractal organization of feline oocyte cytoplasm. Eur. J.
Histochem. 49:151-156.
4. Dumollard, R., Duchen, M. and Sardet, C. 2006.
Calcium signals and mitochondria at fertilization.
Semin. Cell. Dev. Biol. 17: 314-323.
5. Dumollard, R., Duchen, M. and Carroll, J. 2007. The role of mitochondrial function in the oocyte and embryo. Curr. Top. Dev. Biol. 77:21-49.
6. Guo, J., Niu, Y. J., Shin, K. T., Kwon, J. W., Kim, N.
H. and Cui, X. S. 2018. Fatty acid synthase knockout impairs early embryonic development via induction of endoplasmic reticulum stress in pigs. J. Cell.
Physiol. 233:4225-4234.
7. Igarashi, H., Takahashi, E., Hiroi, M. and Doi, K. 1997.
Aging-related changes in calcium oscillations in fertilized mouse oocytes. Mol. Reprod. Dev.
48:383-390.
8. Jackowska, M., Kempisty, B., Antosik, P., Bukowska, D., Budna, J., Lianeri, M., Rosinska, E., Wozna, M., Jagodzinski, P. P. and Jaskowski, J. M. 2009. The morphology of porcine oocytes is associated with zona pellucida glycoprotein transcript contents.
Reprod. Biol. 9:79-85.
9. Kempisty, B., Jackowska, M., Wozna, M., Antosik, P., Piotrowska, H., Zawierucha, P., Bukowska, D., Jaskowski, J. M., Nowicki, M. and Brussow, K. P.
2012. Expression and cellular distribution of INHA and INHB before and after in vitro cultivation of porcine oocytes isolated from follicles of different size.
J. Biomed. Biotechnol. 2012:742829.
10. Liu, L., Hammar, K., Smith, P. J., Inoue, S. and Keefe, D. L. 2001. Mitochondrial modulation of calcium
signaling at the initiation of development. Cell Calcium. 30:423-433.
11. Lasiene, K., Vitkus, A., Valanciute, A. and Lasys, V.
2009. Morphological criteria of oocyte quality.
Medicina. 45:509-515.
12. Malcuit, C., Kurokawa, M. and Fissore, R. A. 2006.
Calcium oscillations and mammalian egg activation.
J. Cell. Physiol. 206:565-573.
13. Ohlweiler, L. U., Brum, D. S., Leivas, F. G., Moyses, A. B., Ramos, R. S., Klein, N., Mezzalira, J. C. and Mezzalira, A. 2013. Intracytoplasmic sperm injection improves in vitro embryo production from poor quality bovine oocytes. Theriogenology 79:778-783 14. Santulli, G., Xie, W., Reiken, S. R. and Marks, A. R.
2015. Mitochondrial calcium overload is a key determinant in heart failure. Proc. Natl. Acad Sci.
U.S.A. 112:11389-11394.
15. Schatten, H., Sun, Q. Y. and Prather, R. 2014. The impact of mitochondrial function/dysfunction on IVF and new treatment possibilities for infertility. Reprod.
Biol. Endocrinol. 12:111.
16. Uyar, A., Torrealday, S. and Seli, E. 2013. Cumulus and granulosa cell markers of oocyte and embryo quality. Fertil. Steril. 99:979-997.
17. Varghese, A. C., Ly, K. D., Corbin, C., Mendiola, J.
and Agarwal, A. 2011. Oocyte developmental competence and embryo development: impact of lifestyle and environmental risk factors. Reprod. Biomed. Online.
22:410-420.
18. Wang, L. Y., Wang, D. H., Zou, X. Y. and Xu, C. M.
2009. Mitochondrial functions on oocytes and preimplantation embryos. J. Zhejiang. Univ. Sci. B.
10:483-492.
19. Warriach, H. M. and Chohan, K. R. 2004. Thickness of cumulus cell layer is a significant factor in meiotic competence of buffalo oocytes. J. Vet. Sci. 5:247-251.
20. Wang, N., Hao, H. S., Li, C. Y., Zhao, Y. H., Wang, H. Y., Yan, C. L., Du, W. H., Wang, D., Liu, Y., Pang, Y. W., Zhu, H. B. and Zhao, X. M. 2017.
Calcium ion regulation by BAPTA-AM and ruthenium red improved the fertilisation capacity and developmental ability of vitrified bovine oocytes.
Sci. Rep. 7:10652.
[Provider:earticle] Download by IP 118.70.52.165 at Monday, December 20, 2021 7:48 PM
www.earticle.net
21. Xu, Y. R. and Yang, W. X. 2017. Calcium influx and sperm-evoked calcium responses during oocyte maturation and egg activation. Oncotarget. 8:89375- 89390.
22. Yang, S. G., Park, H. J., Kim, J. W., Jung, J. M., Kim, M. J., Jegal, H. G., Kim, I. S., Kang, M. J., Wee, G., Yang, H. Y., Lee, Y. H., Seo, J. H., Kim, S. U. and Koo, D. B. 2018 Mito-TEMPO improves development competence by reducing superoxide in preimplantation porcine embryos. Sci. Rep. 8:10130.
23. Zhao, L., Lu, T., Gao, L., Fu, X., Zhu, S. and Hou, Y.
2017. Enriched endoplasmic reticulum-mitochondria interactions result in mitochondrial dysfunction and apoptosis in oocytes from obese mice. J. Anim. Sci.
Biotechnol. 8:62.
(Received 08 January 2019, Revised 19 March 2019, Accepted 25 March 2019)
[Provider:earticle] Download by IP 118.70.52.165 at Monday, December 20, 2021 7:48 PM