대한임상병리사회지 : 제 24 권 1 호
1992
스파르가눔에 서
Superoxide
Dismutase의 정 제 및 부분 특성서울보건전문대학 임상병리과 나 동 진
Key words : SOD from sparganum , CU , Zn-SOD type
1 .
서 론최근 인체 기생층의 숙주 감염경로와 숙주 내 생활기전, 숙주의 면역작용을 조정하는 기전을 밝 히는 일환으로 충체 및 이들의 분비물에서 특정 효소를 분리하고, 그들의 생화학적 특성을 밝히는 연구가 많은 학자들에 의해 진행되고 있다.
기생충이 숙주에 감염될 때 숙주의 방어기전이 발생되는데, 숙주는 기생충의 충체 및 이들이 분비 하는 각종 단백질에 의해 면역계를 형성하여 이들 공격을 방어하게 된다. 한편 이들 기생충들은 숙주 의 방어기전에 대해 여러 가지 단백질과 분비물로 숙주의 방어기전에 대응한다. 그리고 이들은 여러 요인으로 숙주의 면역반응기전에 중요한 역할을 하는데, 그 요인 중 대표적인 것의 하나가 효소 단 백질이다. 이들 효소 단백질 중 단백질 분헤효소 l , 2),
단백질 분해효소 억제인자3) 및
anti-oxidant en- zyme
4) 등이 숙주의 면역반응 억제인지-로 밝혀진 바 있다.기생충이 감염되어 숙주의 기관, 조직을 이행하 면 기관과 조직 내에는 각종 형태의 염증과 병변 이 발생되며, 이들 염증세포는
superoxide radical
,hydrogen peroxide , singlet oxygen , hydroxyl radical
과 같은
oxygen radical
퉁이 생 성 된 다. 이 들radi-
cal은 기생충의 세포, 막 단백질, 핵산 동에 손상을 주므로 이들 숙주세포는
superoxide dismutase
,cat- alase , glutahione peroxidase
동과 같은 여 러anti- oxidant
enzyme을 갖고 있 어reactive oxygen radi-
cal로부터 보호된다4),
Anti-oxidant enzyme
중su- peroxide dismutase(SOD: superoxide , superoxide oxidereductase
,EC
1. 15. 1. 1) 는 호기 적 대사를 하는 기관에서 생성되는superoxide
(02 -)를 산소와 과산 화수소로 전환 (202-+2H+
•H
20
2+0
2 ) 시킴으로서 superoxide의 독성 을 제 거 하는 효소이 다,Superox-
ide
radical 은NADPH
의존성 산화5, 6\ xanthine의 산화7), 그리고 과립구가 식균작용을 하는 동안8)과human natural k
il1er cell
이cytolytic
초기 과정 9) 퉁 생물학적 반응에 의해 생성된다고 보고하였다. 그 리고 기생부위 및 이행부위의 염증에서 생성된su- peroxide
radical 은 기생충에서 생성된 SOD에 의해 즉시 제거된다고 보고하였다10)현 재 몇 종류의 기 생 충에 서
anti-oxidant enzyme
및
oxygen
radical 에 관한 보고가 있는데 lI~14), 특히 SOD는 원충류 15) , 선충류 l6, 17), 흡충류 18, 19) 및 조충류20) 풍에서 SOD는 숙주의 면역반응을 조절함으로서 숙주와 기생충의 상호관계에 중요한 역할을 할 것 으로 생각된다고 보고하였다.스파르가눔 (sparganum) 이 각종 동물 및 인체 감염은 procercoid가 감염된 물벼룩과
plerocercoid (sparganum)
가 감염 된 양서 류, 파충류, 조류, 포유 류 및 이들 이행숙주 (transport host) 에 의해 이루 어진다. 이들이 포유동물에 경구 감염되면 위나 장 벽을 통하여 기생부위로 이동하여 기생한다. 그런 데 현재까지 이들 스파르가눔 충체에서 SOD를 순 수 정제하고, 이들 효소의 생화학적 특징들을 보고 한 자료는 아직 발견치 못하였다.본 연구는 스파르가눔 충체의 SOD가 그들의 생 리적 기능에 미치는 영향, 숙주의 면역반응에 미치 는 영향 그리고 숙주와 기생충의 상호관계 둥을 규명하는 기초 자료를 제시하고자 우선 스파르가 눔 세포질에서 SOD를 순수 정제하여 그들의 생화 학적 특성을 밝히고자 본 실험을 실시 하였다.
n.
재료 및 방법1.
실험재료스파르가눔 충체는 자연 감염된 뱀(유혈목이
Natrix trigria
lateralis) 의 피히- 및 근육조직에서 적 출하였다. 적출된 충체는 생리 식염수와 증류수로 여러 번 세척한 후 냉동 건조시킨 디-음-70
0C
냉 동실에 보관하면서 제반실험에 시료로 사용하였다.2.
효소의 활성도 측정SOD의 활성 측정 은 hypoxanthine과
xanthine oxidase(Sigma)
의 존재 히-에 생 성 되 는 superoxide에 의 해neotetrazilium chloride(NTC : Sigma)
의 환원을 억제하는 반응원리에 따라 측정하였다21) 효소 활성도는 NTC의 환원을
50%
억제히·는 효소의 양 을1
unit 로 하였다.3.
단백질 정량정제괴-정 중 각 분획 및 정제된 효소의 단백질 량은
bovine serum albumin (Sigma)
을 표준 단백 질로 사용하여
Lowry et a
l. 22)의 방법 에 따라 정 량하 였다.4.
조효소 정제1) 세 포질
superoxide
dismutase의 추출 냉동건조된 충체 일정량에 냉각된50 mM potas- sium phosphate buffer(pH
7.8) 을 소량 첨가하여teflon pestle
homogenizer로 균질화 한 후 냉동원심 기(Sorvall RC-5B)
로4
UC
에 서20 , 000
xg로1
시 간 원심분리하였다.이들 상층액을 조효소로 사용하였다.
2) Superoxide
dismutase(SOD) 의 정제 SOD를 정 제 하기 위 해20 mM Tris-HCl buffer (pH
7.2) 로 조효소를 투석 한 다음 동일 완충용액 으로 평 형 된DEAE-Trisacryl M column(2.6 x 15 cm :
LKB) 에 통과 시켰다. 이때 유속은 시간 당60
ml로 하고 분획은fraction collector(LKB 2212
HELILAC) 를 이 용해3ml
씩 100 개 의 분획 을 얻었다. 흡착된 단백질의 용출을 위해 30 번부터 100 번 분획 까지 0 ,’--‘--‘-‘‘‘‘~O각 분획은 파장 280nm로 고정된
spectrophoto-
meter로 단백질 홉광도를 측정하였고, 효소 활성도 를 측정한 후 활성이 높은 분획들을 수집하여 냉 동 건조기
(EYELA Freeze Dryer
FD-1) 로 농축시 킨 다음 정제 단계의 시료로 이용하였다. 이들 부 분 정 제 된 시 료는10 mM citrate buffer(pH 5.0)
로투석한 후 동일 완충용액으로 평형된
CM-Trisa-
cryl M column
(1.6 x 6 cm :
LKB) 에 흡착시킨 디음 등일 완충용액으로 세척히-여 결합되지 않은 단백 질을 제거하고,0-0.5
1\1 NaCl 로 농도구배를 주었 다. 이때 유속은 시간 당40
ml 로 하고, 2ml 씩 분 획을 받아 단백질 흡광도와 효소 활성도를 각각 측정하였다. 활성이 높은 분획을 모아 농축한 후 다읍 정제 단계의 시료로 사용하였다 Ion-exchange column
chromatography 를 실 시 하여 부분 정 제 된 시 료는0.2 M
KCl을 함유한50 mM phos- phate buffer(pH
7.8) 로 평 형 된Sephacryl S-200HR column(
1.6 x 75 cm :
Pharmacia) 에 시간 당20 ml
의 유속으로 통과시켜 1.
4 ml
씩 80 개의 분획을 얻 었다. 그리고 이들 분획의 단백질 흡광도와 효소 활성도를 측정하여 활성이 높게 나타난 분획들을 수집 농축한 후 제만실험에 사용하였다.5.
정제된Superoxide
dismutase의 생화학적^i^~
1) Cyanide의 영 향
Cyanide가 정제된 SOD의 활성에 미치는 영향은
Dryer et a
l. 23) 의 방법 에 따라cyanide
농도를O.
1mM에서 5.0mM 까지 변화시키면서 효소 활성도 를 측정하였다.
2)
Azide의 영향Azide7}
정제된 SOD의 활성에 미치는 영향은Misra and
Fridovich24) 의 방법에 따라azide
농도를 1mM 에서 20mM 까지 변화시키면서 활성도를 측 정 하였다.3)
pH의 영향완충용액의 pH 가 SOD의 활성에 미치는 영향은
Salin and
McCord35) 의 방법 에 따라50 mM sodium acetate buffer(pH 4.0) , 50 mM Tris-HCl buffer(pH 6.0-8.0) , 50 mM sodium carbonate buffer(pH 10.0
-1 1.0) 동을 완충용액으로 효소 활성도를 측정하
였다.
4)
훈자량 측정정제된 효소의 분자량 측정은
Sephacryl S-
200HR
column을 사용하여 측정하였다. 정제된 효소는
0.2 M
KCl을 함유한50 mM phosphate buffer (pH
7.8) 으로 평형된Sephacryl S-200HR column
(1.6 x 75 cm)
에 시 간당 20ml의 유속으로 통과시 켜1.
4 ml
씩 80 개의 분획을 얻어 이들의 단백질 흡광도와 효소 활성도를 각각 측정하였다. 분획 중 활
성이 가장 높게 나타나 분획의
Kav
값을 산정히여 효소의 분자량을 산정히-였다. 이때 표준단백질
(standard marker protein: bovine serum albu- min ; 66
,000
,ovalbumin ; 43
,000
,chymotrypsinogen
; 25
,000
,ribonu
cIease A ;
13 ,000) 을 사용하였다.5)
정 제 단계 별SDS-PAGE
정제 단계별 SOD 의 정제상태를 비교 관찰하기 위 하여 Laemmli36) 의 방법 을 변 형 히 여 전 기 영 꽁히
였다. 조효소와 정제 단계별 효소를 7.5%~12 .5%
polyacrylamide linear gradient
gel 을 사용하여 30mA 로 4 시간 전기 영꽁하였다. 영등이 끝난gel
은
coomassie brilliant blue
R-250(Sigma) 로 염색하 고, gel 에 나타난 단백질 band로 정제상태를 관찰 하였다. 이때 표준단백질의 상대적 이동거리 (Rf) 를 산출하고, 순수정 제 된SOD
단백 질 band의 이 꽁거리를 대응시켜 효소의 분자량을 산정하였다.사용된 표준단백 질은
phosphorylase b (94
,000) bo- vine serum albumin (67
,000)
,ovalbumin (43
,000)
,carbonic anhydrase(30 , 000) , soybean trypsine inhib- itor(20
,100)
, α-lactalbumin(14
,400)
등이 다.ill. 결 과
충체의 세포질에 함유되어 있는 SOD를 정제하 기 위해 조효소륜
DEAE-Trisacryl M
column 에 통 과시켜 단백질 흡광도를 측정한 결과20 mM Tri-
HCl buffer(pH
7.2) 로 용출시킨 분획에서 한 개의 peak( 분획 10~20) 가 나타났고, 0~0.5M NaCI
농 도구배를 주어 용출시킨 분획에서는 4711 의peak
(분획 50~58, 60~69,
71-75
, 79-86) 가 나타났 다. 그리고 이들 분획들에 효소 활성도를 조사한 결과 분획 11 번에서 21 번까지 효소활성이 니-타났 다 (datanot shown).
이들 활성이 나타난 분획을 모아 농축한 후 다음 정제 단계인CM - Trisacryl M
column에 통과시킨 결과 단백질 흡광도는
10mM citrate buffer(pH
5.이로 용출한 분획에서 한 개의 peak( 분획 5~13) 와0-0.5M NaCI
농도구배를 주 어 용출시킨 분획에서는 3 개의(분획36-43
,55- 59
, 60-65) 가 나타났다. 이들의 효소 활성도를 측 정한 결과 분획 37 번에서 42 번까지 효소 활성이 나타나 이들 분획들을 농축한 후 다음 단계의 시 료로 사용하였다 (datanot shown).
이들ion-ex- change
chromatography를 실 시 하여 부분 정 제 된 시 료는Sephacryl S-200HR
column 에 통괴-시 킨 결 과 3 개의 단백질 peak( 분획45-54
,59-64
,66-
69) 를 얻었으며, 이들의 효소 활성도를 측정한 결 과 분획 58-64 에서 활성이 나타나 이들 분획들을 농축하여 제반실험의 시료로 사용하였다. 이들 충 체에서 정제된 SOD의 활성도를 측정한 결과 이들 의 활성도는90
unit/g 이었고, 단백질량은2
1.7 mg/
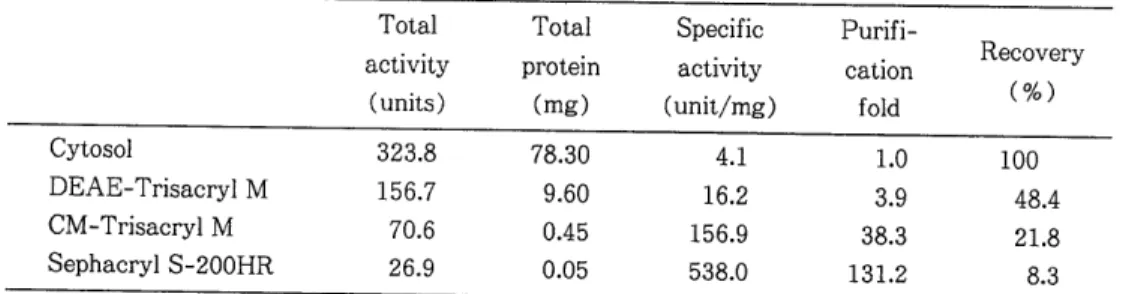
g으로서 비활성도 (specific activity) 는 4.1 이었다.
조효소 및 각 정제 단계별 효소의 정제도를 관찰 한 결과
Table
1 에서 보는 바와 같다.Table 1. Purification of cytosolic superoxide dismutase from Sparganum Total Total Specific Purifi-
Recovery aC
tIvlty protem actlvlty cat
lOn
(units) (mg) (unitfmg) fold (%)
Cytosol 323.8 78.30 4.1
1.0 100
DEAE-Trisacryl M 156.7
CM-Trisacryl M 70.6
Sephacryl S-200HR 26.9
총단백 질
(total protein)
량에 서 조효소는78.3
mg 이었으나 최종 정제된 효소에서는
0.05
mg으로 감소되었다. 총활성도 (total activity) 에서 조효소는323.8
unit 이었으나 최종 정제된 효소는26.9
unit로 서 역시 감소되었다. 그러나 비활성도 (specificactivity)
는4.1
unit 에 서538.0
unit로 증가되 었 다.이상의 결과 최종 정제된 효소는 조효소에 비해서 13 1.2 배의 정제도 (purification fold) 를 보였다.
정제된 SOD의 생화학적 성상에서
SOD
활성에9.60 16.2 3.9 48
.40
.45 156.9 38.3 2
1.8
0.05 538.0 13
1.2 8.3
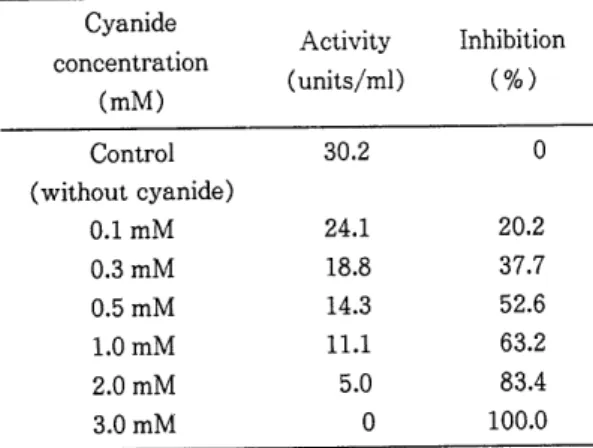
cyanide7}
미치는 영향을 관찰한 결과Table
2 에서 보는 바와 같이cyanide
농도가 증가할수록 효소 활성도가 점차 감소하여 3mM 에서 활성도가100
%
억제됨을 관찰하였다. Azide에 의한 SOD의 억 제효과를 관찰한 결과Table
3 에서 보는 바와 같이 10mM과20 mM
azide 에서 각각 13.6% 과2
1.5%
로 억제 되었다. 완충용액의 pH가 효소 활성도에 미치는 영향을 관찰한 결괴-
Table
4 에서 보는 바와 같이 pH가 증가할수록 활성도가 점차 증가하디-가pH
10 에서 최고 활성을 보여 대조군에 비해 6.2 배 로 증가하였다. 정제된 SOD는 cyanide 에 억제되었 으나 azide에 의해서는 억제되지 않았으며, 완충용 액의 pH가 10.으로 증가하면 효소 활성도가 최대 로 증가하는 것으로 나타났다. 이상의 결과 본 실 험에서 정제된 SOD 가 구리와 아연을 함유한SOD (Cu ,
Zn-SOD) 로 추축된다.Table 2. Effect of cyanide on activity of cytosolic superoxide dismutase in Sparganum.
Cyanide
Activity Inhibition concen
tI‘at
lOn
(mM) (units/ml) ( %)
Control 30.2 0
(without cyanide)
O.lmM 24.1 20.2
0.3mM 18.8 37.7
0.5mM 14.3 52.6
1.
0mM 1
1.1 63.2
2.0mM 5.0 83.4
3.0mM 0 100.0
Reaction mixture considered of 0.2 ml of 0.5 M sodi.
um phosphate buffer(pH 7.5) , 0.1 ml of 16% triton X-100 , 0.01 ml of 10 mM EDTA , 0.3 ml of
1.2 mM neotetrazolium chloride(NTC) , 0.1 ml of 4 mM hypoxanthine , 0.01 ml of xanthine oxidase , cyanide and enzyme in a total volume of 2.0 m
l.Table 3. Effect of azide on activity of cytosolic superoxide dismutase in Sparganum.
Azide
Activity Inhibition concentrat
lOn
(mM) (units/m
l)( %)
Control 30.2 0
(without cyanide)
1.
0mM 30.2 0
5.0mM 30.2 0
10.0 mM 26.1 13.6
20.0 mM 23.7 2
1.5
Reaction mixture considered of 0.2 ml of 0.5 M sodi- um phosphate buffer(pH 7.5) , 0.1 ml of 16% triton X-100 , 0.01 ml of 10 mM EDTA , 0.3 ml of
1.2 mM neotetrazolium chloride(NTC) , 0.1 ml of 4 mM hypoxanthine , 0.01 ml of xanthine oxidase , cyanide and enzyme in a total volume of 2.0 m
l.Table 4. Effect of pH on activity of cytosolic superoxide dismutase in Sparganum.
Activity
Acαti\、va라tionpH (units/ml) (fold)
Control(pH 7
.5)30.2
1.0
4.0 3.0 0.1
6.0 12.1 0
.48.0 63
.42.1
10.0 187.2 6.2
1
1.0 75.5 2.5
Same as the standard assay , except that it contains 0
.5M sodium acetate(pH 4.0) , 0.5 M Tris-HCl(pH 8.0) , 0.5 M sodium carbonate(pH 10.0) as the buff- er.
정 제 된 효소의 분자량을
Sephacryl S-200HR col-
umn 으로 측정한 결과 분획 62 번에서 가장 높은 peak 가 나타나 이 분획의
kav
값을 계산한 결과native
분자량은 33kDa 이 었 다. 또한SDS-PAGE
를 실시하여 정제된 효소의 정제도 및 분자량을 확인한 결과 최종 정제된 효소에서 하나의 단백질
band
(lane
D) 가 관찰되었다. 이들의 분자량을 측정 한 결과16
kDa으로 나타나 스파르가눔 세포질에 서 정제된 SOD는 2 개의 동일한 단백질로 구성된 dimer 임을 확인할 수 있었다. 또 조효소가 정제 과 정을 거치면서 순수하게 정제되었음을 확인하였다.N. 고 찰
현재
superoxide
dismutase(SOD) 가 함유하고 있 는 금속에 따라Mn-SOD
,Fe-SOD
,Cu
,Zn-SOD
동 으로 구분하는데, 이들 효소는 많은 진핵세포와 원 핵세포에서 정제되고 또 그 특성들에 관하여 여러 연구자들이 보고하였다23 , 25-33) • 이들 중Cu
,Zn-
SOD는 고동 동식물의 세포질에 존재하며,
cyanide
에 의해 활성이 억제되며, azide 에는 영향을 받지 않는 것으로 보고되었다34).
여러 종류의 기생충에서 SOD를 정제하고, 그 특 성 이 보고되 었는데, 선충류인
Trichinella spiralis
16),홉충류인
Paragonimus
ψestermaπi18) 과Fasciola he- patica
l9) 동의 세포질에서 정제된Cu
, Zn-SOD가cyanide
농도(1mM-3
mM) 에서 효소 활성도가100%
억제되었다고 보고하였고,azide
농도 (5mM-20
mM) 에서 효소 활성이 억제되지 않는다고 보 고하였다. 본 실험에서 스파르가눔 세포질에서 정 제된 SOD는3 mM
cyanide 에서100%
억제되었으Di, 10mMJ?i- 20mM.9.l azideoJlA-l z.t-4 13.6%, 21.
~<{]-o-1-~.JI, Aafill~ Jt~-cpH 10oJlA-l 3:ltH.9.J
~5% Q:)ftl]!iO]<;ct-c.l-. Aaftll-'8
Jt~.9.J ~_~-(]-€:--pH 10.0oJl
-A(]-~ &~t:j-. 3:.~ *7-}~.g-34kDao]TJ:1,
o]~.g_Al
d':lt:Jl~ ~7}(6.21:ll]) -o}-~_9_TJ:1,pH 1l.OoJ]A1.C 16kDa<{]
%~ t;±~~~ -T-_~-3~dimer 'll%
'?:[9-
_2_
if]
2=1 {} ~ ~-g_- 21!_ O:jT sct t:j-.
:1_c.] .J2 Aj fill-'8
Jl~ ~lsct t:]-.
.C ~Jl~_9.j
1311:ll] AJfil]!iOl<;ct-c.l-. Oj-"J_9.j
~JJl-~.g_T. spiralis, P. westermani, F. hepatica %oJl Al Aa
A-ll~SOD9.} l:ll
~~ 0d'-"J~- &~-c.]-. .:E
~ ~7-]-~oJ]A-j T.spiralis.g- 36 kDa
16l,P. westermani.g- 34 kDa
18l:l_c.j .J2 F.hepatica.g_ 34 kDa
19l _9_~271] .9-j
~-~ ~ t;±~ ~~
T--Acl
~dimertl-.JI &.JI
-o]-~-E t-J],
~1l
~oJ]
A~Aafill~
SOD.9.j native
~7-l-~.g-33kDaclTJ:1, SDS- P AGE
_~-JoJ] Al
~7-l-
~o l 16 kDa 01] Al
~71].9-J band 7}- y-E}y- ol
~ ~J.]
-~~ ~ t;±l1lj ~~-T--AcJ-'8
dimer~ ~Aj~-c.}.
cl-AJ-.9-j
~.i!} ~:i!}..S7}~ Ail.¥.~oJ] A-j Aa
A-ll~SO D-e
~.A}~.cyanide9.} azide7l- Jl
~.9-j ~-Ac}
oJ] u]
~l-Eq3 i>J=, pH7}
Jl~ ~-Ac}oJ] n]
~1~-
"Ji>J= -%
;,~§}"§-[~-Ac}-"J-cl 1!%-fr(T. spiralis),
-~%ff-(P.
westermani, F. hepatica)oJ]A-j
Aaftll~Cu, Zn-SOD31}
%A}~ Jl~~ ;,~ Zf~-c.}.
:]_~~~ ~J!j-g~~ ~.¥.~~~ ~c.] ~~~
SOD-E azideoJ]-E q3
i>J=~- ~.A] ~o}o]
~Aj
fill~Jl~-E
-T-C-]9-J-
o}~% ?:1-%~Cu, Zn-SOD
qJ-~v.
~.7-J-'t}
{J-~-'8 B~ojl),l ~J!j-_E!__7}~%;(l]1J"O}.J2, Ol
~ A-ll.¥.~oJlA-l
superoxide
dismutase(SOD)~DEAE-, CM-Trisacryl M ion-exchange chromatog- raphy 3J Sepharcryl S-200HR column chromatog-
raphy~
1lJ.]-8-}-0:j €-9-
Aafil]-8-}~t:]-. Aafill~SOD.9.j
~_~-a£
-i::- Noritaka et al(1982) . .9.j BJ-l:fl Oll .9-j ill]
~(Cu, Zn-SOD)c]TJ:1,
AJfill~ Jl~.9.j l:l]~-Ac]£-E538.0 unite]
~_9_TJ:1, ~.7-}~.g-33 kDao]
~.Jl,SDS- PAGEoJ] .9-Ji>Jl 16kDa.9.J
t;±~~~ T-_~-a~dimero]
-Ac}o] 100%
~fil]:s:.]~.JI,20mM azideoJ];,-j 21.5%
Q:]ftll:s:.J~t:J-. Aafill~ Jt~-c