대한소화기학회지 2001;37:254-262
서 론
5)B형 간염 바이러스(hepatitis B virus, 이하 HBV로 약함) 는 불완전한 이중 나선 DNA 바이러스로서, 약 3,200개의 염기로 구성되어 있다. 작은 틈새(nick portion)를 가지는 HBV는 4개의 유전자로 구성되어 있으며, 이러한 유전자들
접수: 2000년 12월 19일, 승인: 2001년 3월 6일 연락처: 김학철, 570-711, 전북 익산시 신용동 344-2
원광의료원 소화기내과
Tel: (063) 850-1060, Fax: (063) 855-2025
*이 논문은 2000년 원광대학교 교내연구비의 일부가 지원되었음.
*이 논문의 요지는 1999년 제 5차 대한간학회 춘계학술대회에서 구연 발표되었음.
은 한 유전자가가 다른 유전자와 중복되거나 혹은 양측의 유전자와 중첩되기도 한다.1 각각의 해독틀(open reading frame, 이하 ORF로 약함)을 지닌 4개의 유전자로 C-유전자 (pre-core/core gene), S-유전자(pre-s/ S), P-유전자 및 X-유전 자가 있으며, 이들로부터 각각 HBeAg/HBcAg, HBsAg, DNA polymerase 및 X 단백질 등이 합성된다. 이러한 바이 러스 유전자 산물들의 발현은 mRNA의 합성을 지시하는 4 개의 촉진자(prom otor)들에 의하며, 이들은 2개의 강화인자 (enhancer) 영향을 받아 조절된다.2 - 5
이중 X-유전자는 465개의 염기로 구성되어 있으며, 이에 따라 추론되는 아미노산은 154개로, 주 기능은 교차 활성화 (cis-/trans-activating)로 알려져 있다.6 ,7 이러한 X-유전자는 기능적으로 중요한 3구간으로 나누기도 하고,6 혹은 전반부
만성 B 형 간염 바이러스 보유자에서 X - 유전자 부위의 염기 변이
원광대학교 의과대학 내과학교실
김지웅・송우건・김학철
Mu t a t i o n s o f X-g e n e i n C h r o n i c H e p a t i t i s B V i ru s C a r r i e r s
J i W o o n g K i m M .D ., W o o k e r n S o n g M .D . a n d H a a k C h e o u l K i m M .D
Dep artment of Internal Medicine, Wonkwang University Colleg e of Medicine, Iksan, Korea
Background/Aim s: X-gene product of hepatitis B virus (HBV) is known to cis-/trans-activate a number of cellular and viral promotors. We studied the mutations of X gene and investigated the relationship between the mutations and the severity of disease in chronic HBV carriers. Methods: HBV DNA samples were obtained from the sera of 11 chronic asymptomatic carriers (ASC group), 16 chronic hepatitis patients (CH group), and 23 cirrhotic patients with HBV (LC group). PCR and direct sequencing analysis were performed for X gene. Results: LC group was older than ASC group and CH group . In X gene, there were many mutation points and 17 hot-spots were found. Their variability was 1.17 %. The mutations were not found in the direct repeat (DR) and TATA box binding portion (TBP), but hot-spots were found frequently in the 2nd TATA-box area, negative regulatory element (lNRE) and C/EBP. The nucleotide mutations occurred frequently at 1762 and 1764 nucleotide. The associated mutations at 1762 and 1764 nucleotide were accompanied by the mutation at one of 1653, 16 13 and 163 1 nucleotide. Even in those accompanied mutation, there were not interlinked between them , but has trend to cross-link between them . Conclusions: DR and TBP portion in X gene might be an essential portion for HBV, and especially the 2nd TATA-box, NRE and C/EBP might be related to the severity of liver disease. Moreover there might be inter-, or cross-linked relationship between the mutations. (Korean J Gastroenterol 200 1;37 :254- 262)
K ey W ords: HBV, X-gene, Nucleotide Mutation, Functional Unit, Severity
김지웅 외 2인. 만성 B형 간염 바이러스 보유자에서 X-유전자 부위의 염기 변이 2 5 5
의 아미노산 염기 부위와 중-후반부의 당 염기 부위 (C-terminal)등 두 부분으로 구분하기도 한다.8 바이러스의 전사 등에 관련된 기능적인 부위가 중-후반부 부위에 위치 하며, 이 부위에는 direct repeat 2 (이하 DR2로 약함), 역조 절 인자(negative regulatory element, 이하 NRE로 약함) 및 넓은 의미의 Core Promotor (이하 CP라 약함)부위 등이 존
재한다.3 ,8 ,9 광의의 CP 부위는 약 190여 개의 염기로, 상부
의 조절 서열(upstream regulatory sequence, 이하 URS로 약 함)과 기본 core 촉진자(basic core promotor, 이하 BCP로 약함)로 구성되어 있으며, BCP는 URS에 의해 조절된다고 한다. 이러한 CP는 여러 기능적인 부위가 약간씩은 중첩되 어 상존하지만, URS 부위에는 α-box, β-box, γ-box 및 Enh II 등이 있고, BCP 부위에는 DR 1, hepatocyte nuclear factor (이하 HNF로 약함) III와 IV, TATA-box 및 TATA-box binding portion (이하 TBP로 약함) 등의 부위가 존재하여 여러 기능을 수행한다.8 - 16
한편 HBV에 의한 감염은 자연 회복에서부터 무증상의 만성 보유자, 만성 간염, 간경변증, 전격성 간염 및 간암 등 다양하게 나타난다. 이렇게 다양한 간손상의 원인으로는 HBV의 자체 요인, 감염된 숙주의 요인, 환경적인 요인들로 구분되기도 하며, HBV 자체의 요인으로는 감염된 바이러 스의 양, 독성 및 염기 변이 등이 논의되고 있다.8 , 17 - 19 간암, 전격성 간염의 발병시 특히 X-유전자에서 변이가 흔히 발 견되어 염기 변이가 하나의 중요한 요인으로 대두되고 있
다.2 0 - 2 2 이러한 HBV의 증식은 precore mRNA와 pregenomic
mRNA의 전사에 의하며, 이는 CP 부위에 의해 영향을 받 는다고 알려져 있다.4 ,9 , 17 또한 CP 부위의 변이는 HBeAg와 core 항원의 발현에 영향을 미쳐서 간질환의 활동도 (activity)를 변화시킬 수 있을 것으로 추론하여, BCP 부위 의 염기 변이들이 연구되어 있으며, 일부에서는 URS 부위 까지 연구되고 있다.2 3 - 2 7
이에 저자들은 만성 B형 간염 바이러스 보유자에서 X- 유전자 부위에서 염기의 변이, 그들 간의 상호 상관성 및 간질환의 심도에 따른 염기 변이 등을 관찰하고자 하였다.
대상 및 방법
1. 대 상
대상은 HBV 보유자 중 면역억제제나 조절제, 인터페론 및 항바이러스제를 사용하지 않았던 사람 중, HBsAg이 2 년 이상 양성으로 지속되었던 자로 한정하여, 이들을 3개 대상군으로 분류하였다. 제 1군은 만성 무증상 보유자 (asymtomatic carrier ; 이하 ASC로 약함)의 11명으로 간조직 검사상 특이한 병변을 찾을 수 없었던 경우 6예, 무병변 4 예, 지방간 2예와 간조직검사는 시행하지 않았지만 2년 이 상 HBsAg가 양성이면서 aminotransferase가 정상으로 유지 되었던 5예였다. 제2군은 만성 간염군으로 간조직검사에 의하여 확인된 11예와 관찰 기간 중 aminotransferase가 정 상 범주의 2배 이상으로 6개월 이상 지속되거나, amino- transferase의 상승 기간이 총 9개월 이상이었던 5예로 하였 고, 제3군은 간경변증인 경우로 조직검사에 의하여 확인된 6예와 임상증상으로 확인된 경우 17예였다(Table 1).
2 . 방 법
1) 혈청 HBV DNA추출
대상군의 HBV DNA추출은 QIAamp DNA blood Mini Kit (Qiagen , Hilden, Germany)를 이용하였다. 혈청 HBV DNA의 추출은 동사의 지침서대로 시행하였는데, 이를 약 간 변형하여 시행하였다. 이러한 변형은 HBV DNA 정량 검사(Digene hybrid capture system , Digene Co., Hamden, France)에서 200 pg/mL을 기준으로 하여, 200 μL를 사용하 였는데, 200 pg/mL 이상인 경우는 1 X PBS을 첨가하여 200 μL를 사용하였고, 10 pg/mL 이하인 경우는 혈청과 용 해제를 동비로 증량하였고, 최종 단계의 DNA 추출을 용출 액 100 μL로 사용하였다. 이중 5 μL을 중합효소연쇄반응 (PCR)의 원형으로 이용하였다.
Table 1. Age and Sex Distribution of the 3 Subgroups
Proven by biopsy Number M F Age (yr)*
Asymptomatic carrier Chronic hepatitis Cirrhosis of liver
+ - + - + -
6 5 11 5 6 17
5 3 10 4 5 12
1 2 1 1 1 5
34 .2± 12 .4 37 .1± 10 .2 47 .6± 12 .6†
* mean±SD .
† p<0 .05 vs asymtomatic carrier or chronic hepatitis .
2 5 6 The Korean Journal of Gastroenterology: Vol. 37, No. 4, 2001
2) 시발체 (prime r)의 선택 및 제조
HBV의 4가지 혈청형〔EMBO (European Molecular Biology Organizarion): adr , X046 15; ayw, Y07587 ; ayr, M57663; ady, D 12980에서 전체 X-유전자 영역을 포함하는 부위의 핵산 배열을 관찰하고자, X-유전자의 외측에서 공 통적으로 동일한 염기 배열을 갖는 곳으로 선택하여 sense 로 선택하였으며, antisense는 송 등2 8이 국내의 C유전자 부 위의 염기서열을 분석하여 변이가 가장 적은 부위를 선택 하였다. 1차 PCR에서 사용된 시발체는5'-GGC-TCC-TCT- GCC-GAT-CCA-TAC-TGC-3' (1250→1273), 5'-ATG-TTC-CGG- AGA-CTC-TAA-GGC-3' (204 1→2020)로 증폭산물은 792 염기 쌍(bp)이었고, nested PCR에 이용된 시발체는 5 -GAA- CTC-CTA-GCA-GCT-TGT-TTT-G-3 ( 1289→ 13 10)와 5 - CAG-AAG-CTC-CAA -ATT-CTT-TAT-A-3 ( 1937→ 19 16)로 최종 증폭산물은 649 bp였다.
3) HBV DNA의 증폭
Polymerase 및 dNTPs 등이 적정하게 조성되어 있는 kit
〔Premix IIIR(제노텍, 대전); PyrobestR (Takara Shuzo Co.
Shiga, Japan)=9 ;1들을 혼합하여, 증폭시 염기들의 오결합 (misincoporation)을 교정할 수 있도록 pfu 효소를 이용하였 다. HBV DNA 증폭은 총 20 μL가 되도록 대상 환자부터 분리한 DNA 검체 5μL와 각기의 시발체 20 pm ole, 상기의 혼합물 및 증류수를 가하여 Geneamp PCR system 2400 (Perkin-Elmer, Norwalk, CT, USA)을 이용하였다. 일차 PCR의 조건은 pre-denaturation을 94℃에서 5분, denatura- tion을 94℃에서 1분, annealing은 60℃에서 1분, extension 은 72℃에서 1분씩 하여 총 30회 시행하였으며, 마지막 extension은 72℃에서 5분간 시행하였다. Nested PCR시는 일차 PCR 산물 2μL와 이차 시발체 한 쌍을 넣어서 pre- denaturaion은 94℃에서 5분, denaturation 94℃에서 1분, annealing은 60℃에서 1분, extension은 72℃에서 1분씩 하 여 총 30회 시행하였으며, 마지막 extension은 72℃에서 5 분간 시행하였다. 이상의 모든 과정은 aerozol-tip 등을 사용 하면서, 오염 방지를 위한 수칙2 9을 준수하여 시행하였다.
4) 염기서열의 분석
상기에서 증폭된 2차 PCR 산물을 ethidium bromide으로 처리한 1.2% 한천겔에서 전기영동하여 650 bp의 단일 증폭 밴드를 절단하여 glass bead (Geneclean Ⅱ Kit, Boeheringer Manheim , Germ any)를 이용하여 순수한 유전자를 분리하였 다. 분리된 정제물의 일부를 채취하여 ethidium bromide로 처리한 한천겔에서 시발체들이 없는 단일 밴드임을 확인한 후, 약 50 ng의 정제된 2차 PCR 산물, 2차 PCR에 이용된 시발체 3.2 pM 및 Big dye terminatorc (Perkin-Elmer)을
이용하여 sequencing PCR을 시행하였다. 이는 제조사의 사 용지시서대로 pre-denaturation은 95℃에서 30초, denatura- tion 95℃에서 10초, annealing은 50℃에서 5초, extension은 60℃에서 4분씩 하여 총 25회 시행하였다. 증폭된 PCR 산 물은 에탄올침전법(0.3 M sodium acetate pH 5.2, 70% 에탄 올)으로 정제하여 70% 에탄올로 세척하였다. 이를 Temp- late suppression reagent (Perkin- Elmer)에 녹여, 95℃에서 5분간 denaturation시킨 후 DNA 자동분석기(ABI Prism 3 10 , Perkin-Elmer)에 넣어 분석하였다. 분석한 염기는 sense 와 antisense를 이용하여 양측 검증하였으며, 핵산번호는 EcoR 1 절단 부위를 기준으로 하여 정하였다.
5) 분석된 유전자 염기서열의 다양성
저자들이 제시하는 변이는 아직까지 한국인의 표준염기 가 정해지지 않아서, 각각의 동일한 염기서열에서 대다수 의 염기를 주 염기로, 이와 다르게 발현되는 것을 변이라 정하였다. 다양성은 모든 염기서열에서 변이의 수를 합하 여, 이를 총 염기로 나누어 100배 곱한 것이다. 염기 변이 의 다발 위치는 변이의 평균치±표준 편차를 이용하여 평 균에 대한 신뢰구간 추정시 유의수준이 p<0.00 1 이하보다 많은 곳으로 정하였다.
6) 자료 분석 및 통계 처리
측정된 자료는 평균과 표준편차로 표시하였으며, 그의 기능적인 부위에 따라 분석시 서로 중첩되는 경우가 많아, 알려진 최소의 단위로 분류하였으며, 이러한 최소의 기능 적인 단위에서도 일부가 중첩되는 경우가 있었는데, 이는 최소의 단위로 중복 사용하였다. 자료의 분석 및 통계 처 리는 분율차 검정, Student t test, ANOVA test를 사용하여 p <0 .05 이하를 유의성으로 정하였고, 0. 1<p <0 .05를 높은 경향으로 처리하였다. 제2 TATA -box 부위에서는 자료의 분석, 통계 처리 및 결과까지는 관찰된 바로 기술하였고, 고찰시에는 변이로 계측된 것이 표준염기이었고, 주 염기 가 변이이어서 이를 표준염기(M 12906)로 환산하여 논술하 였다.
결 과
1. 임 상 상 의 차 이
대상인 50명에서 질환군에 따라 연령은 무증상 HBV 보 유자와 만성 간염군에는 차이가 없었으나(34.2± 12.4 vs 37 .1± 10 .2), 간경변군에서 유의하게 높았다(47 .6± 12 .6세 p <0 .05, Table 1) .
Kim, et al. Mutations of X-gene in Chronic Hepatitis B Virus Carriers 2 5 7
2 . X - 유 전 자 염 기 서 열 의 염 기 변 이 , 다 변 이 위 치 및 추 론 되 는 아 미 노 산 의 변 이
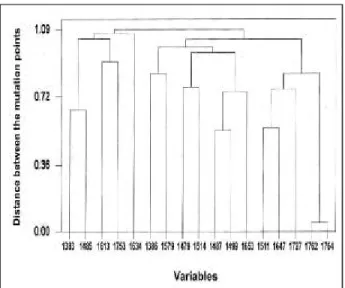
X-유전자 부위에서 염기의 변이는 총 82곳에서 272개가 발견되었다. 이들로부터 계측되는 X-유전자 부위에서의 다 양성은 약 1.17%이었으며, 대상인의 평균 변이는 5.64개이 었다. 질환군에 따른 변이 수는 ASC군에서 5.0± 1.70개, 만 성 간염군 4.75±2.04개, 및 간경변군에서 6.2 1±2.18개로 질환군에 따른 염기 변이는 간경변군에서 많은 경향이었으 나 통계학적 의의는 없었다. 또한 변이된 장소에서 평균 3.3 1±3.94개의 변이가 관찰되어, 변이의 다발 위치는 1383,
1386, 1479, 1485, 1487, 1499, 15 11, 15 14, 1579, 16 13, 163 1, 1647, 1653, 1727, 1753, 1762번과 1764번이었다(Fig . 1) . 염기 변이에 따라 추론되는 아미노산 변이는 167개의 현성 변환이 40곳에서 관찰되었으며, 현성 변환이 일어난 곳에서 평균 4.18±5.23개로 관찰되었으며, 그에 따른 아미 노산의 다양성은 2.7 1%이었다(제시하지 않음).
3 . 기 능 적 인 부 위 에 따 른 염 기 의 변 이 율
X-유전자 부위에서 최소의 기능적인 부위에 따른 변이 율(Table 2)은 DR 1, DR2 및 TBP 부위에서는 변이가 없었 으나(p<0.00 1), β-box에서 0.3%(p<0.05)로 낮거나, 첫 번째 및 두 번째의 SP 1의 부위에서는 0.6%, HNF3 0.6%, HNF4 0 .7 %로 전반적인 X-유전자 부위의 변이율보다 낮은 경향 이었으나, C/EBP에서는 2.0 %로 변이율이 높은 경향이었지 만, NRE 부위에서는 2.1%로 유의하게 높았다(p<0.05). 제2 TATA-box에 해당되는 TBP liver -specific binding TBP에서 는 4 1개의 변이가 보여, 3.15 %의 변이율을 보여, 유의하게 높았다(p<0.00 1, Table 2).
4 . 염 기 변 이 의 특 성
염기 변이가 특이적으로 다발적이고 군집성인 3부위와 소군집성인 2부위가 있었으며, 특이적으로 많은 다변이 위 치를 평균거리법으로 군집분석시 1762번(A T)과 1764번 (T A)의 변이가 가장 밀접하게 동반변이가 일어났으며, 이들과 밀접하게 동반변이가 잘 일어나는 것으로 위치는 1647번(G C)과 1727번(C T), 15 11번(G A) 등의 순이었 으며, 1762번과 1764번의 동반변이와 가장 관련성이 적었 던 것은 1383번(C A)과 1485번(C T)이었다(Fig. 1, 2).
Fig 1. Characteristics of mutation in X-gene from 50 subjects. In X gene, there were many mutation points and 17 hot-spots among them.
Table 2 . Nucleotide Mutation Rate of Variable Functional Unit in X-gene of HBV Genom e from 50 Chronic HBV Carriers Functional unit Nucleotide No . Number of mutations Mutation rate
x-gene NRE DR 1 C/EBP
HNF4 β-box HNF3 lst SP 1 2nd SP 1 TBP liver-speicific TBP
TBP DR2
1373- 1837 16 11- 1634 1590- 160 1 1643- 1658 1657- 1672 1702- 17 13 17 13- 1723 173 1- 1740 1743- 1752 1758- 1773 1786- 1794 1823- 1834
272 26 0 16 6 2 5 3 3 4 1 0 0
1.17 % 2 .08 %*
0 %† 2 .0 % 0 .75 % 0 .33 %‡ 0 .63 % 0 .6 % 0 .6 % 3.15 %† 0 %† 0 %†
NRE, negative regulatory element ; DR, direct repeat ; C/ EBP, CCAAT/enhancing binding protein ; HNF, hepatocyte nuclear factor ; TBP, TATA-box binding protein .
* p<0.05, †p <0 .00 1, ‡p <0 .0 1.
2 5 8 대한소화기학회지: 제37권 제4호 2001
5 . 기 능 적 인 부 위 에 서 표 준 염 기 (M 12 9 0 6 )로 환 산 한 경 우 의 변 이 와 질 환 군 과 의 관 계
1) 기능적인 부위들에서 염기 변이율과 질환군과의 관계 질환군에 따라 TBP liver-specific binding TBP 부위에서 한 개의 염기 변이가 있었던 예는 간경변군에서 23명중 20 예, 만성 간염군에서 12예, A SC군에서 5예로, 간경변군에 서는 ASC군에 비하여 유의하게 높았으나, C/EBP 부위 및 NRE 부위에서는 통계학적 의미는 없었다.
2) 기능적인 부위의 다변이 위치에서의 염기 변이율과 질환 군과의 관계 (Table 3)
(1) TBP live r- s pecific binding TBP 부위에서 1762 번/ 1764번의 동반변이와 질환군과의 관계
1762번/ 1764번의 동반변이시에는 간경변군에서 A SC에 비하여 변이율이 유의하게 높았으나, 만성 간염군과는 차 이는 없었으며, 만성 간염군에서는 A SC군에 비하여 높은 경향이었다.
(2) C/ EBP 부위에서 1647번과 1653번의 변이와 질 환군과의 관계
1647번에서는 질환군에 따라 유의성이 없었으며, 1653번 (C T)에서는 ASC군과 만성 간염군 간에는 차이가 없었으 나, ASC군과 간경변군 간에는 유의한 차이를 보였고, 만성 간염군과 간경변 간에도 유의한 차이를 보였다.
(3) NRE 부위에서 1613번(G A)과 163 1번(C T) 의 변이와 질환군과의 관계
16 13번에서 변이는 ASC, 만성 간염군 및 간경변군에서 각각 1, 3, 6예씩으로 질병의 심화에 따라 증가하는 경향이 었으나, 1631번에서는 0, 1, 5예씩으로 증가하여, 간경변군 에서 A SC군보다 유의하게 많았다(p =0.02). 이러한 NRE부 위에서는 16 13번과 1631번이 상호 동반변이를 보이지 않 아, NRE 부위에서 다변이 위치만을 합하면, ASC군에서 1 예, 만성 간염군 4예 및 간경변군 11예로 증가하여, 질병의 심화에 따라 증가하는 경향을 보여 ASC군보다 만성 간염 군에 비하여 높은 경향이었으며, 만성 간염군에 비하여 간 경변군에서 높은 경향을 보였다. 그리고 간경변군에서는 A SC군보다 유의하게 높은 변이율을 보였다(p =0 .004) .
고 찰
HBV 감염에 의한 간손상의 기전 중, HBV 자체의 요인 들로는 감염원인 바이러스의 양, 독성 및 유전자의 변이 형 태 등이 논의되고 있다.8 , 17 - 19
CP 부위의 변이는 HBeAg와 core 항원의 발현에 영향을 미쳐 간질환의 활동도(activity) 를 변화시킬 수 있을 것으로 추론하여, BCP 부위의 1762번 과 1764번의 염기 변이들이 연구되어 왔으며, 일부에서는 URS 부위까지 연구되어 왔다.2 3 - 2 8 ,3 0 - 3 2 하지만 이러한 염기 들은 X -유전자의 일부로, URS의 부위를 포함하여도 X -유 전자 부위의 약 45%에 지나지 않는다. 이는 X -유전자의 주 기능이 교차 활성화(trans-/or cis-activating)4 ,6 , 15이며 URS의 상부에 NRE 부위가 작용하고 있음15을 생각하면 너무나 단 순화하였다고 생각된다. 따라서 저자들은 X -유전자 부위에 서 염기 변이를 전반적으로 관찰하고, 변이 간의 상호 관련 Fig 2. Relationship between the mutation points in X-gene. The
mutations were occurred frequently at 1762 and 1764 and the 1762/ 1764 mutations were accompanied by the mutations with one of 1647, 1727 and 15 11 nucleotide.
Table 3 . Nucleotide Mutation Point and Its Number of 3 Functional Unit in X-gene of HBV Genom e from 50 Chro- nic HBV Carriers
NRE C/EBP TBP-liver specific binding TBP 16 13 163 1 1647 1653 1762 1764 ASC ( 11)
CH ( 16) LC (23)
1 4 6
0 1 5*
3 1 1
0 2 8†
5 11 19*
5 11 20*
NRE, negative regulatory elem ent; C/ EBP, CCAAT/
enhancing binding protein (included in α-box) .
* p<0.05, mutations were compared to standard sequence (M 12906) at 2nd TATA box and significant difference was noted between asymptomatic carriers and cirrhosis of liver .
†p <0 .0 1.
김지웅 외 2인. 만성 B형 간염 바이러스 보유자에서 X-유전자 부위의 염기 변이 2 5 9
성, 기능적인 부위에 따른 변이율 등을 B형 간염 바이러스 에 의한 질환군과의 상호 유관성을 살피고자 하였다.
HBV 간질환에서 유전자의 변이 형태로 질환의 심도를 논의하고자 함은 우선 HBV에서 그의 염기서열의 다양성 을 이해하여야 할 것3 3으로 생각된다. 이러한 다양성은 유 전자형에 따라 차이가 있으며, 동일한 유전자형내에서도 부위에 따라 다르며, 또한 동일한 유전자형의 유전자 부위 내에서도 그의 다양성은 차이가 있다.3 0 ,3 4 ,3 5
국내의 HBV와 같은 C-유전자형에서도 X-유전자의 다양성은 2.1-2.5 %인 것으로 보고되었다.3 4 본 연구에서 X-유전자 부위에서 그의 다양성이 1.2 %로 전술한 바에 비하여 적었다. 이는 한국인 에서 CP 부위에서 0.95%라고 한 저자들3 1의 보고와 국내의
보고들3 2 ,3 6이 유사하여, 국외 보고들과의 차이는 한국인이
단일 민족임을 고려하면 충분히 수긍될 수 있는 차이라 생 각된다.
X-유전자 부위에서의 염기 변이의 특성을 보면, 군집 분 석시 1762번과 1764번의 동반변이가 대다수의 예에서 일어 났다. 이는 기능적인 부위로 보면 제2 TATA-box로, 이러한 동반변이는 많은 예에서 확인되었다.2 3 - 2 6 이러한 동반변이 시에 α-box의 1653번의 변이나, 혹은 NRE 부위의 1631번 나 16 13번의 변이가 관찰되어, 동반변이가 많이 발생됨을 알 수 있다. 이러한 동반변이에 대한 관련성은 1653번에서 확인되었지만,0 ,3 1 ,3 2
16 13번이나 163 1번에 대해 논한 바 없 다. 이러한 동반변이도 1653번의 변이시에는 163 1번이나 16 13번의 변이는 동반된 예가 적어 마치 시소놀이와 같았 으며, 163 1번이나 16 13번의 변이시에는 상호 동반된 변이 는 하나도 없었다. 이는 BCP의 기능이 URS의 주된 영향을 받으며, NRE 부위는 이를 보정하고 있음5 , 15을 고려하면, 이 러한 동반변이를 유추할 수 있으나, 이는 다른 연구자들에 게서도 향후 확인될 수 있을 것이다.
본 연구에서 X-유전자 부위에서 최소 기능적인 부위를 단위로 하여 관찰시, DR 부위와 TBP 부위에서는 염기의 변이가 전혀 없었던 점은, 이 장소가 HBV의 특성을 보이 는 부위일 가능성을 시사하는 것으로 생각된다. 이러한 점 은 HBV에 의한 모든 간질환에서 유전체가 결손되어도, X- 유전자와 pre-c/C-유전자의 전반부가 모든 예에서 존재하 며,,3 7 만약에 C-terminal 부위-특히 제2 TATA-box의 전방- 에서 8개의 염기가 결손되면 frame-shift가 일어나 HBV의 표지자가 발현되지 않는 간염이 발생된다는 점19과 더불어 HBV가 증식시에 증폭효소(HBV polymerase)가 결합하여 전사가 시작된다는 점10 - 12 , 16을 미루어 전형적인 HBV의 간 염에 특징을 가지는 부위로 생각된다.
TBP liver-specific binding TBP 부위는 두 번째의 TATA- box에 해당되는 부위로, initiator라고도 불리우기도 하며, 여러 물질이 결합되어 작용할 수도 있는 곳인 동시에, 복사
의 시작점으로 자리하기도 한다.9 , 10 , 13 , 14 이제까지의 보고들 은 간질환의 심도(severity)와 염기 변이와의 관련된 보고들 이 주로 BCP 부위에 국한되어 왔으며, 일부에서 URS 부분 이 논의되었다.7 - 3 7 하지만 이러한 부위는 여러 단백질과 결 합할 수 있으며, 결합하는 단백질에 따라 기능적으로 다른 initiator로 인지하거나, 혹은 모든 initiator들이 하나의 인식 단백질로 상호작용하여 그의 활성도를 증강시킬 수도 있
다.9 - 1 1 따라서 X-유전자로부터 발현하는 단백질은 바이러스
및 세포내의 여러 단백질과 상호작용을 할 수 있으며, 또한 RNA와 단백질과도 상호작용할 수 있으며, HBV의 증식에 있어서 이들 간에도 상호작용이 있을 수 있고, X-단백질의 교차 활성화 작용을 하는 데는 필수적인 3구간이 존재한다 는 점6들을 근거로 NRE, URS, 및 BCP 부위까지를 조사하 였다. 하지만 아직까지 본 연구와 같이 X-유전자를 전반적 으로 관찰하고, 이들에서 상호 변이 관계 및 질병군 간의 관련성을 논한 연구가 미약하여, 기존의 보고된 바와 비교 가 가능한 부위와 통계학적 유의성이 있는 다변이 발생 위 치만을 고찰하였다.
또한 TBP-liver specific binding TBP 부위에 변이율이 많 았던 점은 이 부위가 제2 TATA -box 부위로 1762번과 1764 번 동반변이가 빈번하게 이루어지며,2 3 - 2 5 ,3 8 이러한 염기의 변이는 만성 간질환에서 많이 발현하여, 간경변군에서는 만성 간염군에 비하여 높은 경향이었으며 무증상의 보유자 에 비하여 유의하게 높아, 이는 간질환이 심할수록 변이가 심하다는 여러 보고들2 4 - 2 6 ,3 1 ,3 2 ,3 9과 일치하였다. 1762/ 1764번 의 동반 위치로부터 전방으로 진행되는 방향으로 보면 1753번에서 이러한 동반된 변이가 많았고, 이는 간경변군 에서 많았지만 통계학적 유의성은 없었다. 하지만 전격성 간염이나 급성 간염시에는 변이의 발현율이 적은 것에 비 하여 만성 간질환 환자에서 많이 보인다는 보고2 4 ,4 0와 말기 의 간질환에서 결손된 HBV가 많이 출현한다는 보고8 ,3 0가 있으며, 국내에서도 이와 유사한 보고들3 1 ,3 2이 있었다. 따라 서 그의 통계학적 유의성이 관찰된 바는 없지만, 이 부위가 TBP에 결합하는 장소임을 감안하면, 말기의 간질환과 관련 성이 깊을 것으로 생각된다.
α-box의 한 부위인 C/EBP 부위는 변이율이 높았지만, 통 계학적인 의미는 갖지 못하였다. 이는 1653번에서 다수의 염기 변이가 관찰된다는 다른 보고들2 5 ,2 7 ,3 1 ,3 2로부터 추론하 면, 변이율이 높을 것으로 추정된다. 최근 국내의 동일 환 자에서 3-4년이 지나 혈청전환되면서 1653번의 변이가 발 생됨을 보고3 2한 바 있다. 본 연구에서도 ASC군보다 간경 변군에서 유의하게 높았으며, 또한 만성 간염군에 비하여 서도 높은 경향의 빈도를 보여, 간질환이 심해질수록 이러 한 변이가 더 잘 나타나는 것을 통계학적으로 증명한 것이 라 생각된다.
2 60 The Korean Journal of Gastroenterology: Vol. 37, No. 4, 2001
NRE 부위에서도 변이율이 다른 기능적인 부위에 비하 여 변이율이 높았으며, 이들의 부위에 속하는 16 13번과 163 1번에서 간질환이 심해짐에 따라 발현 빈도가 높았다.
이는 BCP의 기능이 URS의 α-box에 의하여 주된 영향을 받으며 NRE 부위는 이를 보정하고 있음을 고려하면, 1653 번의 염기와 163 1번의 염기는 TBP-liver specific binding TBP 부위의 기능에 중요한 영향을 줄 것으로 생각되기 때 문에, 이들의 부위에서 변이율이 높을 수 있을 것으로 생각 된다. 하지만 아직은 구할 수 있는 다른 연구들이 없어 향 후 보다 많은 사례에서 연구되어야 할 것으로 생각되고, transfection을 통하여 확증되어야 할 것으로 생각된다.
X-유전자의 NRE, C/ EBP 및 TBP-liver specific binding TBP 부위에서 표준염기로 환산시, 염기의 변이가 만성 간 염군에서 ASC군보다 발현 빈도가 높은 경향이었으며, 간 경변군에서는 유의성이 있음은 만성 간질환의 심도와 관련 성이 있을 것으로 생각되고, 이는 다른 보고자들과 일치한 다. 본 연구에서 ASC군과 만성 간염군과의 연령에 차이가 없었으나, 간경변군에서 이들보다 연령이 높았다. 그렇기 때문에 국내의 대부분의 HBV에 의한 간질환이 모자수직 감염인 점을 고려할 때 이러한 변이 다발 위치의 차이가 제 2 TATA-box의 변이는 만성 간질환을 결정하는 특성이고 NRE나 C/EBP의 변이는 질병 심도를 반영하는 것인지, 질 병의 자연경과에 의한 변화인지, 혹은 바이러스 자체에 의 한 발병기전인지는 향후 연구 조사되어야 할 것으로 생각 된다.
요 약
목적 : X-유전자는 4개의 ORF 중의 하나인 B형 간염 바 이러스(HBV) 유전자이다. X-유전자 산물은 전사의 활성인 자 중의 하나이며, 여러 세포성 및 바이러스성 촉진자들을 교차활성화하며, 이곳에는 각기 다른 기능적인 부위들이 산재되어 있는데 이들로는 DR, NRE, TBP, C/EBP, TBP liver -specific binding TBP 등이 있다. 따라서 이들 부위의 염기의 변이는 HBV의 증식에 영향을 미치며, 이는 간 질 환의 심도에도 영향이 있을 것으로 생각된다. 이에 저자들 은 만성 HBV 보유자에서 X -유전자의 염기서열을 조사하 여, 발생된 변이, 변이 염기 간의 동반 및 간질환의 중증도 와 비교 분석하고자 하였다. 대상 및 방법 : 만성 B형 간염 바이러스 보유자로서 무증상 보유자 11명(ASC군), 만성 간 염 환자 16명(CH군), 간경변증 23명(LC군)의 혈청에서 DNA를 추출하였다. 이들의 HBV DNA를 염기 분석함으로 써, 염기 변이 간의 관련성 및 질환의 만성적 경과에 따라 서 비교 분석하였으며, 제2 TATA-box에서는 표준 염기서 열(M 12906)과 비교하였다. 결과 : 연령은 LC군에서 ASC군
이나 CH군보다 높았다. 염기의 변이가 다양하였고, 평균 변이의 다양성은 1.17 %이었으며, 다발인 위치가 17곳으로 확인되었다. X-유전자의 기능적인 단위에서 보면 몇 가지 의 특징이 있다. 변이가 없었던 부위는 DR, TBP 부위이었 고, 변이가 호발한 부위는 제 2 TATA-box, NRE, C/EBP 순 으로 중-후반부의 기능적인 부위에서 많이 발견되었다. 염 기 변이는 1762번과 1764번의 염기가 가장 흔하였고, 또한 동반변이도 흔하였다. 1762/ 1764번의 동반변이시에 1653번 의 변이나, 혹은 16 13번과 163 1번의 변이가 흔하였다.
1762/ 1764번의 동반변이시에 1653번의 동반변이는 1762/ 1764번의 변이가 없는 군에 비하여 많았고, 16 13번과 163 1번의 동반 변이는 없는 군에 많은 경향이었다. 이러한 변이와 간질환군을 비교 분석하면, LC군에서 1762/ 1764번, 1653번 및 163 1번의 변이가 ASC군에 비하여 많았다. 결론 : DR과 TBP 부위는 X -유전자 부위에서 가장 특징적인 부위 로 생각되며, 간질환이 심도와 제2 TATA-box, NRE, C/ EBP 부위의 변이와 유관할 것으로 생각되며, 이러한 변 이들도 상호 유관성이 있을 것으로 생각된다.
색인단어 : B형 간염 바이러스, X-유전자, 기능 부위, 염기 변이, 심도
참 고 문 헌
1. Ganem D, Varmus HE. The molecular biology of the hepatitis B viruses. Annu Rev Biochem 1987;56:65 1-693.
2. Schaller H, Fisher M. Transcriptional control of hepa- dnavirus gene expression . Curr Top Microbiol Immunol 199 1; 168:2 1-39.
3. Yuh CH, Ting LP. The genome of hepatitis B virus contains a second enhancer: cooperation of two elements within this enhancer is required for its function. J Virol 1990;64:428 1-4287.
4. Yuh CH, Ting LP. Transcriptional regulation of precore and pregenomic RNAs of hepatitis B virus. J Virol 1992;66:
4073-4084.
5. Kim YH, Kang SK, Lee YI. Functional analysis of hepatitis B virus transactivator X: implication of the leucine zipper- like region and C-terminal seven conserved amino acids in functional regions. Biochem Biophys Res Commun 1993;
197:894-903.
6. Arii M, Takada S, Koike K. Identification of three essential regions of hepatitis B virus X protein for trans-activation function . Oncogene 1992;7:397-403.
7. Uchida T, Saito T, Shinzawa H. Mutations of the X region of hepatitis B virus and their clinical implications. Pathol
Kim, et al. Mutations of X-gene in Chronic Hepatitis B Virus Carriers 2 6 1
Int 1997;47:183-193.
8. Günther S, Piwon N, Iwanska A, Schilling R, Meisel H, Will H. Type, prevalence, and significance of core promoter/ enhancer II mutations in hepatitis B viruses from immunosuppressed patients with severe liver disease. J Virol 1996;70:8318-833 1.
9. Chen IH, Huang CJ, Ting LP. Overlapping initiator and TATA box functions in the basal core promoter of hepatitis B virus. J Virol 1995;69:3647-3657.
10. Yaginuma K, Koike K. Identification of a promotor region for 3.6 kilobase mRNA of hepatitis B virus and specific cellular binding protein. J Gen Virol 1989;63:29 14-2920 11. Lopez-Cabrera M, Letovsky J, Hu KQ, Siddiqui A. Multiple
liver-specific factors bind to the hepatitis B virus core/pregenomic promotor: transactivation and repression by CCAAT/enhancer binding protein . Proc Nat Acad Sci USA
1990;87:5069-5073.
12. Yuh CH, Ting LP. C/EBP-like proteins binding to the functional box-α and box-β of the second enhancer of hepatitis B virus. Mol Cell Biol 199 1; 11:5044-5052.
13. Wang GH, Zoulim F, Leber EH, Kitson J, Seeger C. Role of RNA in enzymatic activity of the reverse transcriptase of hepatitis B viruses. J Virol 1994;68:8437-8442.
14. Wang HD, Yuh CH, Dang CV, Johnson DL. The hepatitis B virus X protein increases the cellular level of TATA- binding protein, which mediates transactivation of RNA polymerase III genes. Mol Cell Biol 1995;12:6720-6728.
15. Park GT, Yi YW, Choi CY, Rho HM. A negative re- gulatory element and its binding protein in the upstream of enhancer II of hepatitis B virus. DNA Cell Biol
1997;16:1459- 1465.
16. Fourel G, Ringeisen F, Flaj olet M, Tiollais P, Buendia MA.
Functional analysis of ground squirrel hepatitis virus enhancer II. J Virol 1998;72:1616-1622.
17. Nishizono A, Hiraga M, Kohno K, et al. Mutations in the core promoter/enhancer II regions of naturally occurring hepatitis B virus variants and analysis of the effects on transcription activities. Intervirology 1995;38:290-294.
18. Sato S, Suzuki K, Akahane Y, et al. Hepatitis B virus strains with mutations in the core promoter in patients with fulminant hepatitis. Ann Intern Med 1995;122:24 1-248.
19. Fukuda R, Ishimura N, Kushiyama Y, et al. Hepatitis B virus with X gene mutation is associated with the maj ority of serologically "silent" non-B, non-C chronic hepatitis.
Microbiol Immunol 1996;40:48 1-488.
20. Takeuchi M, Fuj imoto J, Niwamoto H, Yamamoto Y, Okamoto E. Frequent detection of hepatitis B virus X-gene
DNA in hepatocellular carcinoma and adj acent liver tissue in hepatitis B surface antigen-negative patients. Dig Dis Sci 1997;42:2264-2269.
2 1. Moriarty AM, Alexander H, Lerner RA, Thornton GB.
Antibodies to peptides detect new hepatitis B antigen : serological correlation with hepatocellular carcinoma.
Science 1985;227:429-433.
22. Kaneko M, Uchida T, Moriyama M, et al. Probable implication of mutations of the X open reading frame in the onset of fulminant hepatitis B. J Med Virol 1995;47:
204-208.
23. Okamoto H, Tsuda F, Akahane Y, et al. Hepatitis B virus with mutations in the core promoter for an e antigen- negative phenotype in carriers with antibody to e antigen . J Virol 1994;68:8 102-8 110.
24. Laskus T, Rakela J, Nowicki MJ, Persing DH. Hepatitis B virus core promoter sequence analysis in fulminant and chronic hepatitis B. Gastroenterology 1995;109:16 18- 1623.
25. Fukuda R, Nguyen XT, Ishimura N. et al. X gene and precore region mutations in the hepatitis B virus genome in persons positive for antibody to hepatitis B e antigen : comparison between asymptomatic "healthy" carriers and patients with severe chronic active hepatitis. J Infect Dis
1995; 172:119 1- 1197.
26. Moriyama K, Okamoto H, Tsuda F, Mayumi M. Reduced precore transcription and enhanced core-pregenome trans- cription of hepatitis B virus DNA after replacement of the precore-core promoter with sequences associated with e antigen-seronegative persistent infections. Virology 1996;
226:269-280.
27. Kidd-Ljunggren K, Oberg M, Kidd AH. Hepatitis B virus X gene 175 1 to 1764 mutations: implications for HBeAg status and disease. J Gen Virol 1997;78:1469- 1478.
28. 송우건, 서검석, 김용성, 김학철. 한국인에서 B형 간염 바이러 스 유전자형-핵산 배열로 관찰한 B형 간염 바이러스의 C 유 전자 부위에서-. 대한간학회지 1998;4(suppl):S22.
29. Kwok S, Higuchi R. Avoiding false positives with PCR.
Nature 1989;339:237-238.
30. Buckwold VE, Xu Z, Yen TS, Ou JH. Effects of a frequent double-nucleotide basal core promoter mutation and its putative single-nucleotide precursor mutations on hepatitis B virus gene expression and replication. J Gen Virol 1997;
78:2055-2065.
31. 김용성, 서검석, 송우건, 김학철. 만성 B형 간염 바이러스 보 유자에서 Core promoter의 다양성. 대한소화기학회지 1999;
34:632-643.
32. 김연수, 이진우, 김영석 등. 만성 B형 간질환에서 hepatitis B
2 62 대한소화기학회지: 제37권 제4호 2001
virus Enhancer II 영역의 변이-Basic core promotor 및 precore 변이와의 상관 관계. 대한소화기학회지 2000;36:
2 1-223.
33. Okamoto H, Tsuda F, Sakugawa H, et al. Typing hepatitis B virus by homology in nucleotide sequence: comparison of surface antigen subtypes. J Gen Virol 1988;69:2575-83.
34. Norder H, Courouce AM, Magnius LO. Complete genomes, phylogenetic relatedness, and structural proteins of six strains of the hepatitis B virus, four of which represent two new genotypes. Virology 1994:198:489-503.
35. Kidd-Ljunggren K, Oberg M, Kidd AH. The hepatitis B virus X gene: analysis of functional domain variation and gene phylogeny using multiple sequences. J Gen Virol.
1995;76:2 119-2 130.
36. 정성택, 신용준, 김영수, 김진홍, 조성원. B형 만성 간질환에 서 core promoter 변이의 양상. 대한소화기학회지 1998;31:
780-788.
37. Gunther S, Sommer G, Iwanska A, Will H. Heterogeneity and common features of defective hepatitis B virus genomes derived from spliced pregenomic RNA. Virology 1997;238:363-37 1.
38. Takahashi K, Aoyama K, Ohno N, et al. The precore/core promoter mutant (T 1762 A 1764) of hepatitis B virus:
clinical significance and an easy method for detection . J Gen Virol 1995;76:3 159-3164.
39. 김연수, 김성일, 황성규 등. HBeAg 양성인 만성 HBV 보유 자에서 HBV core promoter 변이의 임상적 의의. 대한소화기 학회지 1997;30:769-778.
40. Alexopoulou A, Karayiannis P, Hadziyannis SJ, et al.
Whole genome analysis of hepatitis B virus from 4 cases of fulminant hepatitis: genetic variability and its potential role in disease pathogenicity. J Viral Hepat 1996:3; 173- 181.