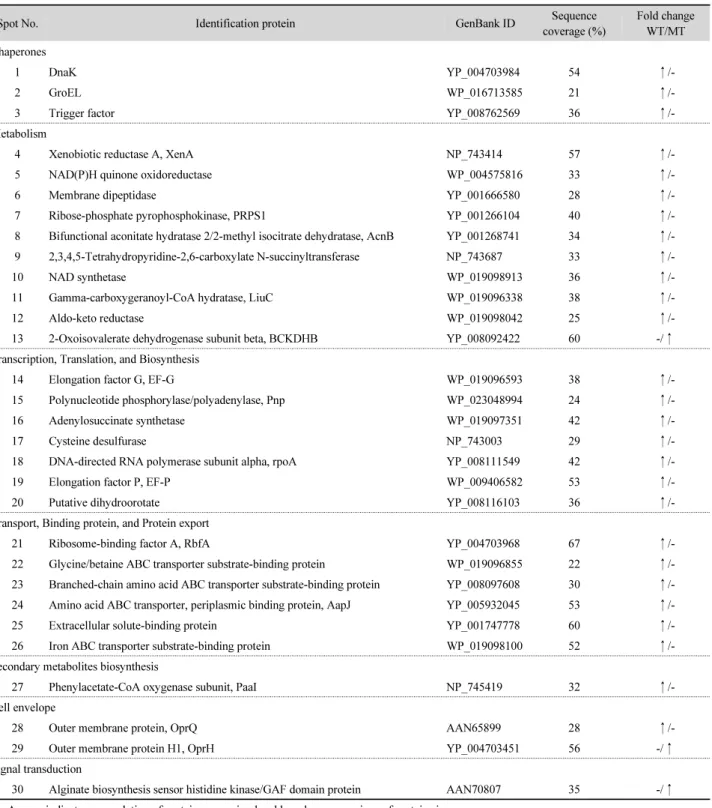
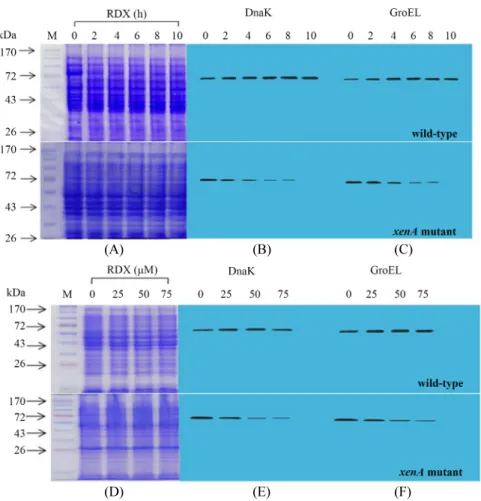
Integrative analysis of cellular responses of Pseudomonas sp. HK-6 to explosive RDX using its xenA knockout mutant
Bheong-Uk Lee
1, Moon-Seop Choi
2, Ji-Won Seok
2, and Kye-Heon Oh
2*
1
Division of Biological Sciences and Chemistry, Kosin University, Busan 49104, Republic of Korea
2
Department of Life Science and Biotechnology, Soonchunhyang University, Asan 31091, Republic of Korea
Pseudomonas sp. HK-6의 xenA 돌연변이체를 이용하여 RDX 폭약에 노출된 세포반응들의 통합적 분석
이병욱
1・ 최문섭
2・ 석지원
2・ 오계헌
2*
1