와송약침액이 Oxidant에 의한 신장세포손상에 미치는 영향
박상원
1․김철홍
2․윤현민
2․장경전
2․안창범
2․송춘호
1동의대학교 한의과대학 1경혈학교실, 2침구학교실
Beneficial effect of Orostachys japonicus A. berger herbal acupuncture on oxidant-induced cell injury in renal epithelial cell
Sang-Won Park1, Cheol-Hong Kim2, Hyoun-Min Youn2, Kyung-Jeon Jang2, Chang-Beohm Ahn2, Choon-Ho Song1
Dept. of 1Meridian & Acupoint, 2Acupuncture & Moxibustion, College of Oriental Medicine, Dongeui University Abstract
Objectives : This study was performed to determine if Orostachys japonicus A. Berger herbal acupuncture (OjB) provides the protective effect against the loss of cell viability and DNA damage induced by oxidant in renal proximal tubular cells.
Methods : The cell viability was evaluated by a MTT reduction assay and DNA damage was estimated by measuring double stranded DNA breaks in opossum kidney (OK) cells, an established proximal tubular cell line. Lipid peroxidation was determined by measuring malondialdehyde (MDA), a product of lipid peroxidation.
Results : H2O2 increased the loss of cell viability in a time-dependent manner, which were prevented by 0.1% OjB. The protective effect of OjB was dose-dependent over concentration range of 0.05-0.5%. H2O2 caused ATP depletion and DNA damage, which were prevented by OjB and the hydrogen peroxide scavenger catalase. The loss of cell viability by H2O2 was not affected by the antioxidant DPPD, but lipid peroxidation by the oxidant was completely inhibited by DPPD. Generation of superoxide and H2O2 in neutrophils activated by phorbol-12,13-dibutyrate was inhibited by OjB in a dose-dependent manner. OjB inhibited generation of H2O2 in OK cells treated with antimycin A and exerted a direct H2O2 scavenging effect. Exposure of OK cells to 1 mM tBHP caused a significant depletion of glutathione which was prevented by OjB. OjB accelerated the recovery in cells cultured for 20 hr in normal medium without oxidant following oxidative stress.
Conclusions : These results suggest that OjB exerts the protective effect against oxidant-induced cell injury and its protective effect was resulted from radical scavenging and antioxidant activities.
Key words : Orostachys japonicus A. Berger, herbal acupuncture, oxidant-induced cell injury, renal epithelial cell
⋅교신저자 : 송춘호, 부산광역시 부산진구 양정2동 산 45-1 동의대 학교 한의과대학 경혈학교실, Tel. 051-850-8643, Fax. 051-853-4036, E-mail: [email protected]
⋅투고 : 2007/02/28 심사 : 2007/03/08 채택 : 2007/03/13
Ⅰ. 緖 論
腎은 五臟六腑의 精을 받아 갈무리하며 五臟이 왕성하면 精을 瀉할 수 있어 先天의 本으로서의 중요성을 가진다
1). 腎臟은 尿의 생성, 酸과 鹽基의 평형, 滲透壓 조절 등 체 액의 항상성을 유지하며 생식, 내분비, 면역 기능감시, 중추신경계통의 일부 생리기능을 담당하고 있다
2,3).
腎臟組織細胞는 다양한 자극에 의해 反應 性酸素基(reactive oxygen species ; ROS)를 발생시킬 수 있음이 생체 실험을 통하여 입 증되고 있다
4).
ROS는 생명유지에 필수적 역할을 하지만 매우 불안정하고 활성이 강하여 생체 내에 서 과도하게 생성될 경우 絲球體腎炎이나 gentamicin에 의한 急性腎不全과 같은 腎臟 疾患의 원인이 된다
5).
生體細胞膜은 불포화지방산을 많이 含有 하므로 ROS에 의해 쉽게 脂質의 過酸化가 일어나고
6), ROS는 직접적으로 또는 脂質의 過酸化를 통해 간접적으로 細胞의 투과성을 증가시키고
7)Na
+-K
+-ATPase와 같은 필수 단백질에 영향을 미친다. 단백질, 지방, 탄수 화물, DNA, RNA는 모두 oxidant에 의해 細胞가 損傷될 수 있지만, DNA분자는 다른 어떤 細胞 보다도 더 큰 損傷을 입을 수 있 다
8).
천연식물에서 추출한 약들이 서양의학계 에서 치료제로 사용되고 있으나 急性腎臟疾 患을 豫防할 수 있는지는 아직 알려져 있지 않다.
瓦松(Orostachys japonicus A. Berger)은
氣味가 凉, 酸苦 無毒하여 淸熱解毒, 利濕, 消腫, 通經破血시키는 작용이 있으므로 牙齦 腫痛, 脣裂生瘡, 惡瘡不斂 등을 치료하며
9,10). 李
11)와 Yoon 등
12)은 瓦松이 神經細胞에서 H
2O
2로 인한 apoptosis를 豫防하며 抗癌, 免 疫效能이 있다고 하였다.
본 연구에서는 瓦松藥鍼液(OjB)이 oxi- dant에 의해 처리된 opossum kidney(OK) 細胞에서 細胞사망을 豫防할 수 있는지 실 험하여 유의한 결과를 얻었기에 보고하는 바이다.
Ⅱ. 材料 및 方法 1. 材 料
1) 試 藥
Hydrogen peroxide (H
2O
2), t-butyl h y d r o p e r o x i d e ( t B H P ) , c a t a l a s e , luciferin-luciferase, thiobarbituric acid, malonaldehyde tetraethylacetal, phorbol-12, 1 3 -dibutyrate ( P D Bu) , 5 -am ino-2 , 3-dihydro-1, 4-phthal-azinedione (luminol), NaOCl, 3-[4,5-dimethylthiazol-2-yl]-2, 5-diphenyltetrazolium bromide (MTT)는 Sigma Chemical (St. Louis, MO, U.S.A.)社 로부터, [
3H]methylthymidine는 American International (Amersham,U.K.)社로부터, N,N'-diphenyl-p-phenylenediamine (DPPD) 는 Aldrich Chemical(Milwaukee, WI, U.S.A.)社로부터 구입한 제품을 사용하였고 그 외 기타 모든 試藥은 특급품을 사용하였
다 .
2) 藥 材
실험에 사용한 瓦松은 시중에서 구입하여 정선해서 사용하였다.
2. 方 法
1) 瓦松藥鍼液 제조
483 g의 粉碎된 가공되지 않은 瓦松에 6,000 ㎖의 증류수를 가한 후 100℃에서 3시 간 동안 加熱하고 침전물을 여과하였다. 여 과한 추출물은 rotary evaporator로 減壓濃 縮하였으며 동결건조 후 78.2 g의 건조물을 얻어 냉동 보관하였다. 이를 실험에 사용하 기 직전 Hank's balanced salt solution (HBSS, Sigma Co., U.S.A.)에 용해시켜 사 용하였다.
2) 주머니쥐 OK 細胞 배양
OK 細胞는 American Type Culture Collection(Rockville, MD, U.S.A.)에서 분주 받아 75 ㎠ culture flask에서 일련의 단계를 거쳐 維持되었다(Costar, Cambridge, MA, U.S.A.). 이 細胞들은 10%의 fetal bovine serum이 포함된 Dulbeco's modified Eagle's medium 과 Ham's F12 (DMEM/F12, Sigma Chemical Co.) 培養液으로 CO
2培 養器(37℃, 5% CO
2)에서 培養하였다.
培養된 細胞의 subculture는 0.02%
EDTA-0.05% trypsin 용액을 사용하여 준비 하였고, 이 細胞들은 10%의 fetal bovine se- rum을 含有하고 있는 DMEM/F12 培養液 으로 24-well culture plate에서 培養되었다.
모든 實驗은 細胞를 培養한지 3-4日 후에 시작되었다. 細胞는 115 NaCl, 5 KCl, 25 NaHCO
3, 2 NaH
2PO
4, 1 MgSO
4, 1 CaCl
2,5 glucose(pH 7.4)를 함유하고 있는 HBSS에 서 H
2O
2로 처리되었다.
3) 細胞 生存率 測定
細胞 生存率 測定은 MTT assay를 이용 하였다. 細胞는 24-well dish에서 培養된 후 HBSS로 부드럽게 씻어 37℃ 5% CO₂培 養器에서 도표에 따라 지정된 時刻에 H
2O
2에 노출시켰다. 細胞를 씻은 후 HBSS에 0.5
㎎/㎖를 포함하고 있는 500 ㎕의 MTT용 액을 각각의 24-well dish에 加하고 위에 펼쳐진 환경조건을 제거하고 나서 60分 동 안 細胞를 培養하였다. 그리고 5分동안 3 DMSO 110 ㎕를 加한 후, 초소형 plate 눈 금을 이용해서 550 ㎚에서 吸光度를 測定하 였다(Molecular Device Corp., Menlo Park, CA, U.S.A.). Data는 control의 백분율로 자 료를 나타냈으며 이는 H
2O
2가 없는 상태에 서 실시되었다.
4) ATP 內容物 測定
細胞內의 ATP 수준은 luciferin-luciferase
분석에 의해 測定되었다. Oxidant stress에
노출된 후 細胞들은 0.5% triton X-100의
500 ㎕에 용해시켰고, 0.6M perchloric acid
100 ㎕에서 산화시킨 후 얼음위에 놓아 두었
다. 細胞의 suspension은 4 mM MgSO₄(pH
7.4)를 함유하고 있는 10 mM의 potassium
phosphate buffer를 넣고 희석시킨 후, 희석
전 이 10 ㎕의 sample에 100 ㎕의 20 ㎎/㎖
luciferin-luciferase를 가하였다. Luminometer (Micro Lumat LB96P, Berthold, Germany) 에 빛의 방출이 20 s로 나타나도록 하였으 며, 단백질용량은 細胞 suspension의 aliquot 에 의해 결정하였다.
5) DNA 단일나선구조 測定
DNA 가닥 파괴는 DNA 凝結分析에 의해 測定되었다
13). 24-well dish에서 培養된 細 胞들은 0.25 μCi/㎖[
3H]methylthymidine에 24시간 동안 있게 한 후 라벨을 붙여 놓았 다. 細胞들을 HBSS를 이용하여 철저하게 씻은 후 0.5 ㎖의 lysis buffer(10mM Tris/HCl, 10nM EDTA, 50 mM NaOH, 2% SDS, pH 12.4)와 함께 effendorf tube에 용해시킨 후 0.12M KCl 0.5 ㎖를 추가하였 다. Lysate는 65℃에서 10分間 培養 후 다시 5分間 얼음 위에서 식힌 후 응결시켰다. 이 러한 상태에서 DNA 단백질인 K-SDS 凝結 物이 형성되고 여기서 저입자의 끊어진 DNA가 방출되도록 하였다. 이 DNA는 1 0℃, 200 g에서 10分間 원심분리하여 recover된 후 200 mM HCl 1 ㎖를 함유하고 있는 액체의 섬광약병(scintillation vial)으로 옮겨졌다. 이중나선구조의 DNA에서 그대로 保存된 凝結物은 65℃, 1 ㎖의 물속에서 용 해되었다. 시험관을 1 ㎖의 물로 씻고 8 ㎖ 의 섬광액을 각각의 약병에 첨가하였으며, 남아 있는 이중 나선구조의 DNA 量을 각 각의 sample에 대해서 계산하였다.
計算 方法= d.p.m값
전체d.p.m값+위에뜨는내용물 ×100
DNA 損傷정도는 단일나선구조 DNA 對 전체나선구조 DNA(이중나선구조+단일나선 구조)로 표현되었다.
6) 脂質의 過酸化 測定
脂質의 過酸化는 Uchiyama와 Mihara의 方法
14)으로 그 産物인 malondialdehyde (MDA)의 量을 測定하여 평가하였다. 腎臟 細胞는 차가운 1.15% KCl(5% wt/vol)용액 속에서 破碎하였다. 이 組織 破碎 均質液 0.5 ㎖에 1% phosphoric acid 3 ㎖와 0.6%
의 thiobarbituric acid 용액 1 ㎖를 첨가 하여 끓는 물에서 45分 동안 가열하였다.
n-butanol 4 ㎖를 첨가하여 완전히 섞은 다 음 2,000 g에서 20分 동안 遠心分離한 후 上層液의 吸光度를 535와 520 nm에서 測定 하였다(Hewlett Packard, 8452A). MDA 값 은 단백질 1 ㎎당 pmoles로 표시하였고, 단 백질 농도는 Bradford의 方法
15)으로 測定하 였다.
7) Chemiluminescence 分析에 의한 Radical Scavenging 測定
Phorbol esters에 의해 活性化된 neutrophil속 에 있는 luminol-dependent chemiluminescence 를 사용해서 測定하였다
16).
(1) Neutrophil 준비
토끼의 neutrophil은 뉴질랜드산 수컷 흰
토끼로부터 standard dextran sedimentation
과 Grisham
17)의 方法에 따라 gradient sepa-
ration on Histopaque-1077을 사용해서 末梢
血液에서 分離하였다. 이러한 과정을 통하여 trypan blue exclusion assay에 의해 95% 생 존력이 있는 neutrophil을 生成해냈다.
(2) Protocol 분석
Neutrophil (1x10
5cells/㎖)은 다양한 농 도의 OjB와 함께 또는 없이 Kreb's Ringer-phosphate buffer 2 ㎖ 속의 luminol (0.96 ㎍/㎖)과 培養되었고 20 μM PDBu에 의해 活性化되었다. Chemiluminescence는 chemiluminescence分析器 (Biolumet LB 9505, Berthold, Germany)를 사용해서 測定 하였다.
8) H2O2 Scavenging 測定
(1) 螢光性染色을 사용한 測定
H
2O
2를 scavenge 할 수 있는 OjB의 能力 은 OK細胞內에서 mitochondrial electron transport의 방해물질인 antimycin A에 의해 처리된 細胞內에서의 H
2O
2에 대한 OjB의 效能을 測定하여 평가되어졌다. H
2O
2細胞 內 生成의 변화는 2’,7’-dichlorofluorescin di- acetate(DCFH-DA)
18)를 사용해서 測定되었 다. 非螢光性 ester는 細胞속으로 침투해서 細胞의 esterase에 의해 DCFH로 가수분해 되었다. DCFH는 細胞의 peroxidase 그리고 hydrogen peroxide, fatty acid peroxide와 같 은 ROS가 있음으로 해서 高度의 螢光合成 物質인 2’,7’-dichlorofluorescin(DCF)으로 급 속히 酸化되었다. 細胞는 35 mm tissue cul- ture petri dish에서 培養되었다. 培養媒介物 이 제거되고 trypsin-EDTA 溶液을 사용해
서 flask에서 細胞를 채취하였다. DMEM /F12를 이용해서 細胞를 두 번 씻고 나서 螢光分析을 하기위해 glucose가 含有되어 있 지 않은 HBSS에 浮遊시켰고 반응은 螢光 cuvette속에서 이루어졌다. 細胞는 3 ㎖의 glucose가 함유되어 있지 않은 HBSS와 ethanol에 있는 20 mM DCFH-DA 용액으 로부터 나온 20 μM DCFH-DA를 함유하고 있는 螢光 cuvette에서 OjB가 있거나 또는 없는 상태에서 37℃로 10분간 미리 培養했 다. 미리 培養한 이후에 細胞를 antimycin A로 처리하고 나서 최대 60분간 培養하였 다. 이 시간 동안에 螢光性의 정도는 ex- citation wave length가 485 nm 이고 em- mission wave length가 530 nm인 spectro- fluorometer (SPEX 1681, SPEX Co., U.S.A.)로 관찰되어졌다. DCF 螢光性(임의 상수단위)의 순수 증가수치는 antimycin A 첨가 전후의 값 차이를 알아내어 계산되어 졌다.
(2) Chemiluminescence를 사용한 測定
H
2O
2의 농도는 Muller의 方法
19)에 따라
chemiluminescence의 변화에 의해 測定되
어졌다. 이 分析實驗은 sodium hypochloride
(NaOCl)에 의한 luminol의 산화현상에 바탕
을 두고 있다. luminol은 NaOCl에 의해
two-electron oxidation에서 diazaquinone으로
산화된다. 이것은 더 나아가 특히 H
2O
2에 의
해 α-hydroxy-hydroperoxide를 경유해 ex-
cited aminophthalate로 변화한다. 이러한 반
응의 짧은 螢光性 신호는 최대 파장 431 nm
를 지니고 있다. 이 파장은 직선 모양으로
H
2O
2에 의존된 것으로 최소 10
-9M 범위에 까지 이른다. 50 μM NaOCl 100 ㎕를 50 μ M luminol 100 ㎕와 다양한 농도의 H
2O
2를 담고 있는 시험관에 다양한 농도의 OjB가 있거나 또는 없는 상태에서 첨가하였다.
NaOCl의 첨가는 빠른 속도로 chem- iluminescence를 증가시켰으며 이러한 변화는 60초 동안 測定되었다.
9) Glutathione(GSH) 內容物 測定
6개의 plate에서 培養한 細胞를 3%
perchloric acid 0.5 ㎖에 넣어 40℃에서 15분 간 처리한 후 이 細胞를 5분 동안 2,000 g에 서 均質化시켜 遠心分離하였다. Glutathione 含有量은 DTNB-GSSG reductase recycling 方法
20)으로 測定하였다. 전체 glutathione 含 有量을 測定하기위해 25 ㎕의 上層液을 6 mM DTNB 100 ㎕와 6.3 mM EDTA와 143 mM sodium phosphate buffer(pH 7.5) 에 용해된 NADPH 700 ㎕(0.248 mg/㎖)와 물 175 ㎕에 混合시켰다. 5-10 ㎕ GSSG reductase를 첨가함으로써 반응이 시작되었 으며 412 nm에서 absorbance의 변화 속도가 10분간 관찰되었다. GSSG를 測定하기 위해 0.04M N-ethylmaleimide(NEM) 20 ㎕와 0.1 N NaOH를 上層液 50 ㎕에 첨가해서 遊離된 GSH를 차단하였다. 20분 후에 이 샘플은 위에서 언급한 것처럼 含有되었고, 전체 glutathione과 GSSG 사이의 차이는 GSH값으로 취해졌다.
10) 統計分析
成績은 平均値±標準誤差(Mean±S.E.)로 나타내었고, 통계분석은 SPSS version 10.0 을 사용하였으며, 對照群과 實驗群과의 平均 의 차이를 Student's t-test로 檢定하여 p값 이 0.05미만일 때 有意性이 있는 것으로 판 정하였다.
Ⅲ. 成 績
1. H2O2에 의한 細胞 生存率에 미치는 OjB의 效能
細胞 生存率의 중대한 손실은 H
2O
2에 노 출시킨지 30分 후에 나타났고 최고 180分까 지 증가했다. 그러나 細胞 生存率의 손실은 0.1%의 OjB를 동시에 첨가함으로써 豫防 되었다.
OjB의 豫防效力을 알아보기 위해 細胞를 다양한 OjB 농도를 가진 0.5 mM H
2O
2에 노출시켰다. OjB 농도에 비례하여 H
2O
2로 인한 細胞死亡은 豫防되었고 뚜렷한 豫防效 果가 0.05%에서 나타났다. 자료에는 나타나 있지 않지만 OjB는 H
2O
2처리되지 않은 對 照群 細胞에서는 아무런 효력을 발휘하지 않았다(Fig. 1, 2).
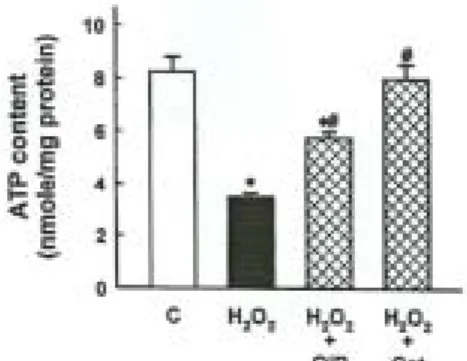
2. H2O2에 의한 ATP 枯渴에 미치는 OjB 의 效能
H
2O
2는 ATP를 對照群(3.44±0.18 vs
8.23±0.56 nmole/mg protein in control)의
약 42% 수준으로 감소시켰다. H
2O
2로 인한
ATP 枯渴현상은 0.5% OjB에 의해 豫防되
었고, H
2O
2scavenger enzyme catalase는
ATP枯渴을 豫防하였다(Fig. 3).
Fig. 1. Time course of H2O2-induced cell death in opossum kidney (OK) cells. Cells were exposed to 0.5 mM H2O2 for various times in the presence or absence of 0.1%
Orostachys japonicus A. Berger herbal
acupuncture (OjB). Cell death was eval- uated by a MTT reduction assay. Data are Mean ± S.E. of six experiments.
*p<0.05 compared with H2O2 alone.
Fig. 2. Effect of Orostachys japonicus A. Berger herbal acupuncture (OjB) on H2O2-induced cell death in opossum kidney (OK) cells.
Cells were exposed to 0.5 mM H2O2 for 60 min the presence or absence of vari- ous concentrations of OjB. Cell death was evaluated by a MTT reduction assay. Data are Mean ± S.E. of six experiments.
*p<0.05 compared with control(C).
Fig. 3. Effect of Orostachys japonicus A. Berger herbal acupuncture (OjB) and hydrogen peroxide scavenger on H2O2-induced ATP depletion in opossum kidney (OK) cells.
Cells were exposed to 0.5 mM H2O2 for 60 min the presence or absence of 0.5%
and 300 U/㎖ catalase (cat). Data are Mean ± S.E. of four experiments. *p<0.05 compared with control (C), #p<0.05 com- pared with H2O2 alone.
3. Oxidant에 의한 DNA 損傷에 미치는 OjB의 效能
0.5mM H
2O
2에 노출된 細胞는 결과적으로 DNA 損傷이 증가되었으며 이중나선 구조 의 DNA가 감소되었다(37.02±4.19 vs 86.55±5.93% in control). 이러한 변화는 0.5% OjB(60.95±4.27)를 첨가함으로 해서 豫防되었다. 이와 유사하게 H
2O
2에 의한 DNA 損傷은 catalase에 의해 豫防되었다 (Fig. 4).
4. H2O2에 의한 細胞死亡에 미치는 OjB 와 Antioxidant의 效能 比較
OjB(0.5%)와 catalase(300 U/㎖)는 0.5
mM H
2O
2에 의한 細胞 生存率의 손실을 豫
防하였다. 對照的으로 강력한 antioxidant인
DPPD 20 μM은 H
2O
2에 의한 細胞 生存率 손실에 영향을 미치지 못했다. 이는 H
2O
2로 인한 細胞死亡이 lipid peroxidation과 연관 되어 있지 않음을 나타낸 것이다. H
2O
2로 인한 細胞死亡에 DPPD의 豫防效果가 없는 것은 oxidant가 이 연구에서 사용된 농도에 서 lipid peroxidation을 유도해내지 못했거 나 아니면 DPPD가 lipid peroxidation을 抑制하지 못했기 때문으로 추정된다. 이를 확인하기 위하여 lipid peroxidation에 대한 H
2O
2와 DPPD 의 效能을 검정한 結果 H
2O
2는 lipid peroxidation을 눈에 띄게 증가 시켰으며 이것은 OjB와 DPPD에 의해 豫防 되었다(Fig. 5, 6).
Fig. 4. Effect of Orostachys japonicus A. Berger herbal acupuncture (OjB) and hydrogen peroxide scavenger on H2O2-induced DNA damage in opossum kidney (OK) cells. Cells were exposed to 0.5 mM H2O2 for 60 min the presence or ab- sence of 0.5% and 300 U/㎖ catalase (cat). Data are Mean ± S.E. of four experiments. *p<0.05 compared with con- trol (C), #p<0.05 compared with H2O2 alone.
Fig. 5. Comparison of effects of Orostachys
japonicus A. Berger herbal acupuncture(OjB), hydrogen peroxide scavenger, and antioxidant on H
2O
2-induced cell death in opossum kidney (OK) cells.
Cells were exposed to 0.5 mM H
2O
2for 60 min the presence or absence of 0.5% and 300 U/㎖ catalase (cat), and 20 μM N,N'-diphenyl-p-phenylenediamine (DPPD). Data are Mean ± S.E. of four experiments. *p<0.05 compared with control (C), #p<0.05 compared with H
2O
2alone.
Fig. 6. Effect of Orostachys japonicus A. Berger herbal acupuncture (OjB) and antioxidant on H2O2-induced lipid peroxidation in opossum kidney (OK) cells. Cells were ex- posed to 0.5 mM H2O2 for 60 min the presence or absence of 0.5% and 20 μM N,N'-diphenyl-p-phenylenediamine (DPPD).
Data are Mean ± S.E. of four experiments.
*p<0.05 compared with control (C),
#p<0.05 compared with H2O2 alone.
5. Neutrophil에서 Superoxide와 H2O2 生 成抑制에 미치는 OjB의 效能
Neutrophil을 다양한 농도의 OjB를 첨가 한 상태에서 phorbol ester의 일종인 PDBu 를 가지고 자극했을 때 chemiluminescence 强度가 급속히 증가해서 4분내에 최대정점 에 달하였으며 그 후 최저 수준으로 감소하 였다. OjB를 첨가하자 chemiluminescence의 최고수준이 낮추어졌다. OjB는 0.002∼
0.05% 농도에서 chemiluminescence최고 수 준을 抑制하였다(Fig. 7, 8).
6. H2O2 生成抑制에 미치는 OjB의 效能
Antimycin A에 依存的인 DCF螢光性은 0.01%의 OjB를 첨가함으로해서 감소되었다.
Antimycin A로 처리된 細胞속에 DCF螢光 性에 대한 OjB의 效能은 그 분량에 따라 다 르게 나타났다. DCF螢光性의 强度는 antimycin A를 첨가한 이후 30분 동안 1.92±0.45에서 4.11±1.02 arbitrary unit로 증 가했으며 이것은 0.01%의 OjB에 의해 2.24±0.54 arbitrary unit로 抑制되었다(Fig.
9, 10).
7. H
2O
2除去에 미치는 OjB의 직접적인 效能
Luminol과 다양한 농도의 H
2O
2를 함유하 고 있는 媒介物에 NaOCl을 첨가하면 chemiluminescence성향이 분량에 따라 비례 해서 증가한다. 이러한 증가현상은 OjB 첨 가에 의해 抑制되었다(Fig. 11).
Fig. 7. Effect of Orostachys japonicus A. Berger herbal acupuncture (OjB) on in phorbol 12,13-dibutyrate (PDBu) - induced pro- duction of reactive oxygen species in hu- man neutrophils. Neutrophils (1x10
5cells/
㎖) were incubated with luminol (0.96 g/
㎖) in 2 ㎖ of Kreb's Ringer-phosphate buffer in the absence or presence of OjB for 4 min. A phorbol ester PDBu (20 μM) was added and chemiluminescence was measured using a chemiluminescence analyzer.
Fig. 8. Effect of Orostachys japonicus A. Berger herbal acupuncture (OjB) on generation of reactive oxygen species in neutrophils ac- tivated by a phorbol ester. Experiments were performed as described in Fig. 1.
Data are mean ± S.E. of three
experiments. *p<0.05 compared with con-
trol (Cont).
Fig. 9. Time course of the generation of re- active oxygen species in OK cells dur- ing exposure to antimycin A (AA). Cells were treated with 20 μM AA in the presence or absence of 0.01%
Orostachys japonicus A. Berger herbal
acupuncture (OjB) and changes in DCF fluorescence were measured. Data are mean ± S.E. of three experiments.
Fig. 10. Effect of Orostachys japonicus A. Berger herbal acupuncture (OjB) on antimycin A (AA)-induced increase in DCF fluo- rescence in opossum kidney (OK) cells.
Experiments were performed as in Fig.
3 except for the addition of various con- centrations of OjB. Data are mean ± S.E. of three experiments. *p<0.05 com- pared with AA alone.
Fig. 11. H2O2 scavenging effect of Orostachys
japonicus A. Berger herbal acupuncture(OjB). 100 μl of 50 μM NaOCl was added to tube containing 100 ㎕ of 50 μM luminol and various concentrations of H2O2 in the presence or absence of OjB. Chemiluminescence was measured at 431 nm for 60 sec. Data are mean
± S.E. of four experiments.
OjB는 분량에 의존하는 방식으로 chem- iluminescence의 强度를 抑制하였다. 0.05%
에서 OjB의 效能은 catalase 500 units/㎖
(H
2O
2scavenging enzyme)와 유사하였다 (Table 1).
Table 1. H
2O
2scavenging effect of OjB.
Group Chemiluminescence Control 483.62±1535.96 OjB(0.005%) 154.92±1105.55*
OjB(0.01%) 0.27±0.14*
OjB(0.05%) 0.001±0.001*
Catalase(500units/ml) 0.002±0.001*
*p<0.05 compared with control.
8. GSH의 枯渴에 미치는 OjB의 效能
GSH 내용물은 t-BHP에 의해 현저히 감
소되었으나 0.001% OjB에 의해 豫防되었다
(Fig. 12).
9. Oxidant로 처리된 細胞에 미치는 OjB 의 效能
Oxidant에 의해 損傷을 입은 細胞의 生存 率은 OjB가 없는 상태에서는 15.63±1.25%였 지만 OjB가 있을 때는 30.29±1.54%로 회복 되었다(Fig. 13).
Fig. 12. Effect of Orostachys japonicus A. Berger herbal acupuncture (OjB) on t-butyl hy- droperoxide (tBHP)-induced depletion of glutathione (GSH) in opossum kidney (OK) cells. Cells were treated with 1 mM tBHP for 60 min in the presence or ab- sence of 0.001% OjB, and then gluta- thione content was measured. Data are mean ± S.E. of four experiments.
*p<0.05 compared with control, #p<0.05 compared with tBHP alone.
Fig. 13. Effect of Orostachys japonicus A. Berger herbal acupuncture (OjB) on the recov- ery following oxidative stress in OK cells.
Cells was treated with 0.5 mM H
2O
2for 60 min and then were cultured for 20 hr in normal medium without H
2O
2in the presence or absence of 0.1% OjB. Cell viability was evaluated by a MTT reduc- tion assay. Data are mean ± S.E. of four experiments. *p<0.05 compared with H2O2 alone.
Ⅳ. 考 察
腎은 先天之本으로 五臟六腑之精을 받아 藏하며 伎巧를 出하고, 腎主水, 主納氣, 通於 腦, 其華在髮, 開竅於耳 司二陰하며
3), 命門, 三焦와 밀접한 관계를 가지고 生殖器, 內分 泌系 및 泌尿器系統을 포괄한다
21). 水液代謝 는 吸收, 運輸, 運化를 主管하는 脾의 기능 과, 宣發肅降 通調水道를 主管하는 肺의 기 능과, 水를 主管하고 氣化作用을 하는 腎의 기능에 의하며, 脾肺腎 중에 腎이 水液代謝 의 중심적인 역할을 담당한다
22).
서양의학에서 腎臟은 체액의 量과 전해질 組成維持, 代謝로 생긴 尿酸이나 크레아틴과 같은 노폐물의 배설, 毒素 및 毒性 代謝産物 의 해독 또는 체외배설, 細胞外液의 量과 혈 압의 內分泌性 조절, 赤血球 총량 조절, 鑛 物質代謝의 內分泌性 조절, peptide 호르몬 의 분해 및 理化作用, beta-2 microglobulin 등 작은 蛋白의 理化作用 등을 담당하고 있 다
23). 인체의 수분은 체중의 약 50-70%이며 정상상태에서는 수분의 섭취량과 배설량이 비슷하여 체액의 平衡狀態를 유지하며 腎臟 은 소변을 형성함으로써 노폐물을 배설하고 酸鹽基의 평형을 조절하여 체액의 恒常性을 유지한다.
瓦松은 돈나물과(crassulaceae)에 속한 다
년생초본인 바위솔의 全草로 昨葉荷草, 屋上
無根草, 向天草 등의 異名이 있으며 氣味가
凉,酸苦 無毒하고 口中乾痛, 水穀血痢, 大腸
下血, 小便沙淋, 頭風白屑, 牙齦腫痛, 脣裂生
瘡, 湯火灼傷, 惡瘡不斂, 風狗咬傷 등을 다스
리는 효능이 있다
9). 主成分으로 大量의
oxalic acid를 함유하고 있고 血熱로 經絡이 不行하고 通하지 못하여 생기는 月經不順을 凉血 行血시켜 淸熱解毒, 止血, 利濕, 消腫시 키는 작용이 있어 吐血, 鼻衄, 血痢, 肝炎, 血疾, 熱淋, 痔瘡, 濕疹, 癰毒, 痔瘡, 湯火灼 傷 등을 다스린다
10).
사람은 나이가 들어감에 따라 抗酸化機能 은 감소하는 반면 活性酸素와 같은 ROS는 체내에 축적되어 細胞나 組織을 파괴하고 여러가지 退行性 疾患을 유발하여 생체의 기능을 약화시킨다
24). 본 실험에 사용된 H
2O
2는 ROS의 일종으로 生體內 代謝過程 中 부수적으로 생기는 活性酸素로, 細胞內顆 粒 및 cytosol, 백혈구에서도 생성되고
25), 脂 質의 過酸化反應을 진행시켜 細胞膜의 파괴, 細胞의 노화, 細胞의 괴사 그리고 DNA에 대한 細胞損傷 등을 유발시켜 생체의 기능 을 약화시키고 노화를 진행시키므로 노화의 정도와 抗老化 효과를 測定하는 기준으로 응용하기 쉬워 최근 많은 연구가 진행되고 있다
26).
ROS는 生體內에서 활성이 강하여 여러 組織에서 심한 독성을 나타내거나 過酸化脂 質의 생성을 유도하여 老化 ,發癌 그리고 心 筋梗塞, 關節炎 그리고 알츠하이머병과 같은 여러 가지 급성 및 만성 神經性疾患의 병인 으로 인정되고 있다
27). ROS는 正常細胞의 미토콘드리아에서 代謝過程 中 발생되고 있 으나 몸속에는 이들을 제거하는 효소나 물 질들을 가지고 있어 細胞속에서 발생되는 有害酸素基들이 細胞損傷을 일으키지 못하 도록 조절하고 있다. 그러나 藥物 또는 細胞 內 환경의 변화로 ROS의 발생이 증가되거
나 ROS를 제거하는 防禦作用이 약화되었을 때 細胞는 損傷을 받게될 것이고 심하면 死 亡에 이르게 된다
28). 따라서 ROS에 의한 細 胞損傷을 豫防할 수 있는 새로운 anti- oxidant 藥의 개발이 요구되어져 왔으며 細 胞損傷에 대한 방어기전을 증대시키는 藥物 의 개발은 대단한 의미를 지니게 되는 것이 다.
天然植物에서 추출한 藥은 西洋醫學에서 강력한 치료제로서의 영향력을 발휘하고 있 으며 현재 사용중인 藥들 중에는 이러한 植 物에서 抽出하여 만들어진 것이 많다. 예를 들면 steroids, cardiotonic glycosides, anti- cholinergics, analgesics, antimalarials, anti- cancer agents 등을 들 수 있다
29).
抗酸化관련 실험연구들은 單味劑, 複合處 方, 藥鍼製劑 등을 이용하여 腎臟機能, 肝機 能, 腦․神經組織 등에서의 oxidant에 의한 물질 이동장애를 방지하는 작용이 있음을 보고한 바 있다
30).
본 硏究에서 OjB는 腎臟 上皮細胞에서
oxidant인 H
2O
2로 유발되는 細胞 生存率의
손실을 뚜렷하게 豫防하였다(Fig. 1, 2). 또
한 OjB는 oxidant에 의한 ATP 枯渴現象을
有意하게 감소시켰다(Fig. 3). ATP 枯渴현
상은 oxidative stress에 대한 細胞들의 초기
반응현상임이 입증되었다
31). 비록 ATP 枯
渴이 oxidant로 인한 細胞死亡의 病因에 관
련되어 있는 것으로 주장되고 있지만
32)다
른 연구들에 의하면 oxidant에 의한 損傷은
ATP 枯渴과는 관계가 없는 것으로 밝혀졌
다
33). 그러므로 H
2O
2로 인한 細胞死亡이
OjB에 의해 豫防된다는 것이 ATP 枯渴의
豫防에 의한 것인지는 명확하지 않다.
DNA는 oxidant stress의 중요한 細胞 및 分子의 표적이다. Oxidant stress는 단일 가 닥의 損傷을 유발하고 DNA 염기 損傷 또 는 apoptosis 유발로 인한 DNA 損傷을 가 져오는 結果를 초래한다
33,34).
본 연구는 OjB가 hydrogen peroxide scavenger catalase와 유사하게 H
2O
2로 유발 되는 DNA 損傷을 효과적으로 豫防함을 입 증했다(Fig. 4). 그러나 DNA 損傷과 cell lysis 사이의 관계가 논란이 되고 있기 때문 에 본 연구 결과만으로는 OjB의 豫防效力이 DNA 損傷 豫防에 영향을 미치는지 分明하 지는 않다. DNA 損傷은 in vitro 에서는 細 胞死亡에 중요한 작용을 하고 있다는 많은 연구가 발표되었다
33,34). 하지만 in vivo에서 DNA 損傷은 정상적인 物質代謝 과정
중 충분히 생성될 수 있기 때문에 腎臟 上皮細胞
31)와 hepatocyte
35)에서 oxidative stress로 인한 細胞死亡의 주요한 매개체는 아니다.
生體膜細胞들은 不飽和脂肪酸을 많이 함 유하고 있어서 이것은 특히 oxidant에 의한 過酸化物攻擊(peroxidative attack)에 약하고 결론적으로 脂質의 過酸化가 발생하게 된다
6)