호냉성 해양세균 Shewanella sp. L93로부터 Eicosapentaenoic Acid 생산 및 정제를 위한 최적화 조건
모상준1·홍혜원2·방지헌3·조기웅3*
1명지대학교 생명과학정보학부, 2한국의약품시험연구소, 3안양대학교 해양생명공학과 Received : May 31, 2011 / Revised : August 4, 2011/ Accepted : August 5, 2011
Optimal Condition for Eicosapentaenoic Acid Production and Purification from Psychrophillic Marine Baterium Shewanella sp. L93. Mo, SangJoon1, Hye Won Hong2, Jiheon Bang3, and Ki Woong Cho3*.
1Division of Bioscience and Bioinformatics, Myongji University, Yongin 449-728, Korea, 2Korea Drug Test Labora- tory, Seoul 130-864, Korea, 3Department of Marine Biotechnology, Anyang University, Incheon 417-883, Korea
− To obtain eicosapentaenoic acid (EPA)-producing bacteria, some 600 strains of bacteria were isolated from Antarctic sediment and marine organisms during the summer expedition of 1999-2000 and 7 EPA-producing bacteria were obtained through screening with TLC and GC. A strain designated as L93 showed the highest EPA production, which was gram-negative, rod-shaped bacterium. L93 strain was identified as Shewanella sp., from the sequence analysis of 16S rDNA. Optimal conditions temperature and pH for the growth and EPA production were about 4oC and pH 7. In addition, its production was optimized by 50%(w/v) sea salt. We establish the optimal production system to produce about 320 mg per liter by using this optimal EPA produc- tion conditions. EPA-methyl ester was purified from cultured L93 strain to a purity of higher than 97% and typical purification yield is greater than 72% of the input amount via urea complexation and HPLC.
Key words: Eicosapentaenoic acid, psychrophillic, marine bacterium, Shewanella sp. L93
서 론
1970년대 초반 에스키모인들을 역학 조사하던 덴마크 Dyerberg 그룹은 에스키모인들이 고지방 식생활임에도 불구 하고 동맥경화, 뇌경색, 심근경색 등의 순환계질환의 발병률 이 인근 유럽인들에 비해 매우 낮다는 흥미롭고도 의외의 사 실을 발견하였다[1, 6]. 고지방 식생활 패턴으로 인해 심장 질환에 의한 사망률이 가장 높았던 서구인들은 이 정반대의 현상에 주목하였고, 1980년대 초 에스키모인들의 현저히 낮 은 심장질환 발병률이 생선의 어유에 많이 포함된 n-3 계열 고도불포화(오메가-3) 지방산으로 불리는 특별한 지방산이 그 원인임을 밝혀내었다[5, 7]. 이에 따라 어유로부터 오메 가-3 계열 고도 불포화 지방산들을 추출, 정제하여 의약 및 건강식품으로 개발하려는 시도가 선진 각국에서 이루어져 왔다.
Eicosapentaenoic acid(EPA)는 대표적 오메가-3 지방산으 로 혈액내 지질감소, 혈전 용해, 관상동맥, 심장질환의 억제, 암세포의 선택적 파괴, 류머치스성 관절염 치료 등의 효능
을 보이고 있는 것으로 보고되었다[6-8]. 등 푸른 생선은 EPA의 함량이 10% 이상으로 비교적 높고, 화학적 합성보다 는 손쉽다는 장점, 또 어류가 전통적으로 많이 이용되어 온 식품이라는 점 등에서 EPA 생산의 주된 원료로 사용되어 왔 다. 그러나 어유에서의 생산에는 다음과 같은 문제점들이 있 는데, 우선 어종에 따른 어획시기에 크게 영향을 받으므로 원료 공급의 안정성이 결여되어 있고 어획 장소로부터 EPA 생산을 위한 장소로 이동이 필요하므로 화학적으로 불안정 한 EPA의 함량유지를 위해 냉동 등의 특수 보관 및 이동 설 비가 필요하며 어체 자체의 지방을 포함한 EPA 이외의 불 순물이 다량 포함되어 있어 EPA의 분리, 정제에 많은 어려 움이 있다[15]. 이러한 이유로 등푸른생선의 장내에 공생하 고 있는 장내미생물로부터 EPA를 생산하고자 많은 연구가 있었으며, 이중 일본의 Sagami 화학 연구소에서 등푸른생선 장내미생물 20,000 주 이상에서 EPA 생산 균주 500주를 획 득하였고, 이 중 최초로 EPA를 균주 1 g 당 최고 300 mg까 지 생산하는 SCRC-2738(Shewanella putrefaciens)을 분리하 여 보고하였다[16]. 그 이후 많은 연구자들이 EPA를 생산하 는 다양한 해양 미생물을 분리하여 보고하고 있다 [10, 11, 14]. 미생물을 이용한 EPA 생산은 어유에서의 추출에 비해 fermentation 등의 생물공학적 공정을 통해 공급원을 안정시 킬 수 있으며 이에 따라 대량생산이 가능한 이점이 있다. 또
*Corresponding author
Tel: +82-32-930-6030, Fax: +82-32-930-6215 E-mail: [email protected]
한 Yazawa 등에 의해 분리된 SCRC-2738 균주 등의 경우 와 같이 EPA 이외의 여타 고도 불포화지방산이 매우 적게 함유되어 있어 미생물을 이용한 EPA 생산방법은 어유에서 의 추출에 비해 분리추출 공정이 손쉽고 경제적인 장점도 있 다[12]. 이외에도 특히 미생물로부터의 EPA 생산은 짧은 배 양기간과 비교적 손쉬운 배양조건 확립도 큰 장점이라 하겠 으며 균주 개량 및 생산조건 개선을 통한 발전의 여지가 많 이 남아있다. 위에서 언급한 내용을 바탕으로 본 연구는 1999 년부터 2000년까지 남극하계 연구기간 중 채집된 시료로부 터 EPA 생산 균주를 수집 확보하였고, 이중 가장 생산능력 이 뛰어난 L93 균주를 분리하였고, 균주의 EPA 생산 특성 및 분리 정제 방법을 확립하였다.
재료 및 방법 균주의 분리 및 보존
본 실험에서 사용한 균주는 1999년과 2000년 남극하계 연 구기간 중 채집된 명명되지 않은 남극 토양과 해양생물에서 분리하였다. 해양생물에서 채취한 시료는 SCUBA로 채집된 즉시 2-3 g의 시료를 취하여 멸균된 가위와 칼로 잘게 자른 다음 멸균된 해수와 함께 멸균된 비닐 bag에 넣어 즙을 짜 그 즙을 ZoBell 2216E(Difco) 한천 배지에 0.1 ml 씩 분주 하고 도말 하였다. 남극 저서 퇴적물은 SCUBA를 이용하여 깊이 32 m의 수심에서 채집되었으며, 채집된 해역은 한국해 양연구소 남극 세종기지 앞 Marian cove에서 얻어졌다. 채 집 당시 수온은 0.5~0.8oC 였다. 채집된 시료는 기지로 옮겨 져 ZoBell 2216E 배지를 기본으로 한 plate에서 각 균주가 순수 분리되었으며, 분리된 균주는 야외 Igloo chamber(임야 실험용 간이 실험실, 평균 온도 2-5oC)에서 10 일간 배양되 었다. 형성된 균체 집락을 그 집락 형태 특성(색깔, 집락 형 태 등)에 따라 새로운 고형배지에 접종하여 동일 형태의 순 수 집락을 형성하도록 배양한 후 형성된 각 집락들을 각각 백금이를 사용하여 3단계 희석도말법으로 순수 분리하였다.
순수 분리, 배양된 해양 세균은 멸균된 1.5 ml microcen- trifuge tube에 1 ml씩 넣고 13,000 rpm에서 2분간 원심분리 후 상층액을 따라버린 다음 멸균된 해수로 두 번 세척 한 후 20% glycerol(v/v) 용액에 현탁 시킨 다음 -70oC 에서 보관 하였다.
EPA 함유 여부 검색
n-Hexane, acetyl chloride, chloroform, methanol, ethanol, diethyl ether, Butyl hydroxy toluene(BHT) 들은 Merck (Darmstadt, Germany)의 제품을 사용했으며 fatty acid 들과 fatty acid methyl ester 표준 시료는 Sigma-Aldrich(St.
Louis, MO, USA)의 제품을 사용하였다. 본 실험에 사용한 모든 용매에는 항산화제로서 50-100 mg/l의 BHT를 첨가하 였으며 지방산의 methylation에 사용된 methanolic HCl의
제조는 acetyl chloride 30 ml을 4oC 식힌 메틸알콜 300 ml 에 천천히 부으면서 섞어 형성된 HCl이 메틸알콜에 용해된 용액을 제조하였다. ZoBell 2216E 고체 배지에서 3일간 배 양한 균체 약 50 mg 가량을 접종용 loop를 사용하여 micro- centrifuge tube(2.2 ml)에 들어 있는 0.3 ml의 methanolic HCl에 옮긴 후 5초간 vortex로 흔들어 현탁 시킨다. 현탁 된 시료를 55oC 항온조에서 2시간 동안 반응시켜 methylation 을 완결한 후 0.4 ml 5% NaCl 용액과 1.5 ml n-hexane을 가한 후 30초간 vortex로 흔들어 형성된 fatty acid methyl ester를 n-hexane 층으로 추출한다. 깨끗한 층 분리를 위하여 약 1분간 15,000 rpm에서 원심분리한 후 n-hexane 층을 pasteur pipette 을 사용하여 새로운 1.5 ml microcentrifuge tube로 옮긴 후 이를 Speed-Vac(Vision, Korea)을 사용하여 시료 부피를 약 0.1 ml로 감압 농축 시킨 후 농축된 n- hexane 추출액 내의 EPA 검색을 일차로 박막 크로마토그라 피(Thin Layer Chromatography, TLC)를 사용하여 수행하였 다. 사용된 박막은 Silica TLC Plate(G-254, Merck, Darm- stadt, Germany)를 사용하여 위에서 준비된 fatty acid methyl ester 시료 약 10 µl를 점적하여 n-hexane과 diethyl ether의 9:1 혼합액을 전개용매로 사용하여 30분간 전개시켰다. 결과 검색은 전개 종료 후 박막 판을 말린 후 요오드(I2) 증기에 의한 지질의 착색법에 의해 검출하였다[3].
Analysis of gas chromatography
EPA 이외의 여타 고도불포화 지방산들도 본 TLC 조건하 에서는 동일 위치에 반점이 나타나므로 TLC를 사용한 일차 검색의 결과 EPA methyl ester에 해당되는 위치에 전개 반점 이 나타난 20여 종의 시료를 대상으로 gas chromatography (GC)를 사용하여 최종 확인하였다. EPA의 함유 여부는 Sigma-Aldrich(St. Louis, MO, USA)에서 구입한 EPA를 위 에서 언급한 methylation 과정을 거쳐 최종 확인을 위한 시 료로 사용하였다.
지방산 조성 분석은 Hewlett-Packard HP 5890 series II plus를 사용하였고 정량 결과는 chromate s/w를 이용하여 peak의 면적을 계산하였다. Column은 Omegawax 320(0.32
×30 m)을 사용하였고 injector 온도는 250oC, detector(FID) 는 260oC, oven 온도는 200oC, initial column head pressure (psi)는 9, split-ration는 25의 조건으로 수행하였다[3, 4].
16S ribosomal DNA 유전자 염기서열을 이용한 계통분류 Genomic DNA는 Wizard genomic DNA purification kit(Promega, USA)를 이용해 추출한 후 16S rDNA 유전서 열 분석에 사용하는 universal primer인 27F(5'-AGAGTT- TGATCATGGCTCAG-3')와 342R(5'-CACGGATCCCCA- CTGCTGCSYCCCGTAG-3') primer를 사용하여 PCR 증폭 하였다[9]. 16S ribosomal DNA 유전자를 PCR 온도조건은 96oC에서 5분간 예비가열 후, 96oC 1분간 변성, 50oC 1분
annealing, 72oC 1분간의 중합반응 과정을 25회 반복하고, 마지막으로 72oC에서 10분간 처리한 후 반응을 중단했다. 증 폭된 PCR 산물은 QIAquick PCR purification kit(Quiagen, Valencia, CA, USA)을 이용하여 정제하고, 정제된 PCR산물 은 ABLPRISM 3700 DNA Analyzer를 이용하여 염기서열 을 분석하였다. 그 결과는 BLASTN 프로그램을 이용하여 GenBank의 ribosomal DNA 유전자 서열과 비교하였으며, Phylogenetic tree 작성은 Clustal X와 The National Center for Biotechnology Information(NCBI, http://www.ncbi.nlm.
nih.gov/)에서 제공하는 Advanced BlastSearch 프로그램을 통하여 GenBank에 보고된 유사 균주와의 염기서열을 비교 하여 계통분류학적 유연관계를 분석한 후, MEGA v4.0을 이 용하여 Tamura-Nei distance model과 neighbor-joining method에 의해 계통도를 만들었다.
EPA 생산 및 배양 특성
균주의 성장과 EPA 최적 생산 조건을 찾기 위해 초기 균 주 접종량을 600 nm에서 흡광도가 0.1이 되게 접종하였다.
최적 온도는 5, 10, 15, 20oC에서 5일간 배양하며 확인하였 고, 최적 pH는 초기 pH를 5, 6, 7, 8, 9로 각각 다르게 적 정 된 배지에 접종하여 확인하였다. 또한, 최적 염도 요구도 를 찾기 위해 sea salt(염전 소금)를 사용해서 10, 50, 100, 200%(w/v)로 제조한 해수를 사용 배양 배지를 제조하여, 균 주를 위에서 언급한 방법대로 접종한 후 4oC, pH 6 에서 5 일간 배양하여 확인하였다.
EPA 분리 정제
EPA를 분리정제를 위해 urea 냉침전법을 사용하였다. 우 선, Sea Water Complete(S.W.C) 배지(0.5 %(w/v) tryptone, 0.3 %(w/v) yeast extract, 0.3 %(w/v) glycerol, 50 %(w/v) sea salt, pH 7.0)에 L93 균주를 접종하여 4oC에서 배양한 후
균체를 원심분리하여 회수하였다. 회수한 균체를 동결건조한 후 건조된 균체 1 g을 30 ml의 Folch solution (chloroform : methano = 2 : 1)으로 3회 추출하고 농축하였다. 농축한 추 출물을 위에서 언급한 방법으로 fatty acid methyl ester로 변 형시켰다. Urea 1.5 g이 들어 있는 10 ml 용기에 methanol 1 ml을 가하여 섞은 후 L93 균주에서 추출한 0.1 g의 fatty acid methyl ester를 hexane에 녹여 그 중 4 ml 가하였다.
이 용액을 상온에서 4~5시간 천천히 흔들어 주면서 방치하 여 urea 표면에서 용해와 결정화가 일어나게 한 후 영하 20oC에서 하룻밤을 방치하여 결정화를 완결하였다. 다음날 이를 여과하여 결정을 제거하고 미리 영하 20도로 식혀놓은 hexane 10 ml로 결정을 세척한 후 이를 다시 걸러 sodium sulfate로 수분을 제거한 후 질소 가스를 이용하여 1 ml까지 농축시켰다. 이 농축액의 지방산 조성 및 EPA의 농축도를 GC를 이용하여 분석하였다[4]. 최종 정제는 HPLC는 SpectraPhysics의 P-1000 pump를 사용하였고 column은 Tosoh Co.에서 구입한 ODS-120 reverse phase column(5 µm, 4.6×150 mm)을 사용하였다. 이동상은 methanol(100%, (v/v))을 사용하였고, 유속은 1 ml/min, 검출은 206 nm에서 확인하였다[13]. 분리 정제 과정마다 매번 EPA standard을 사용하여 분리 정제 순도와 수율 확인을 하였다.
결과 및 고찰
남극 하계연구기간 중 남극 토양과 해양생물에서 분리한 균주를 그 집락 형태나 색등에 따라 약 600여 종을 분리하 였다. EPA 생산 세균을 검색하기 위해 TLC를 사용하여 1 차 검색한 결과 20여 종의 균주에서 EPA가 생산되는 것을 확인하였고, 이 균주들의 EPA 생산여부를 최종 확인하기 GC를 사용하여 7개의 EPA 생산 균주를 찾았다. 이들 중 EPA의 함량이 가장 높은 L93번을 대상으로 본 연구를 수행
Fig. 1. Phylogenetic tree generated via the neighbor-joining method showing the phylogenetic relationships of the isolated Shewanella sp. L93 strain among Shewanella species.
하였다. L93 균주는 그램 음성으로 현미경 사진에서 전형적 인 간균 형태를 보였고, ZoBell 고체 배지에서 배양한 균주 는 오렌지색의 콜로니를 형성하였다. 16S rDNA 염기서열을 이용하여 균주를 동정하기 위해 재료 및 방법에 언급한 primer 를 사용하여 PCR을 행하였고, L93의 16S rDNA의 전체 염 기서열을 결정하였다. 그 결과, 다양한 Shewanella 속 미생물 중에서도 Shewanella gelidimarina와 99%의 높은 상동성을 나타내어, 분리된 균주의 16S rDNA의 염기서열은 Shewanella 속으로 조사되었다. 이에 분리된 균주를 Shewanella sp. L93 라 명명하였다(Fig. 1).
남극 저온성 세균 L93은 20oC에서는 거의 성장을 보이지 않았다. 초기 균주 접종량을 600 nm에서 흡광도 0.1로 조정 하여 접종한 상태에서 5일까지 균주의 성장은 전혀 증가하 지 않았으며, EPA 함량도 초기의 7.8%에서 지속적으로 감 소하여 5일째에는 0.1% 수준으로 감소하여 세포가 분열하 여 성장하는 대신 고온에 적응한 상태로 지방산 조성만을 변 화시키며 생존하고 있는 것으로 나타났다(Fig. 2). 반면에 불 과 5oC 차이인 15oC에서는 매우 활발한 균주 성장을 보였 다. 배양 후 3일째부터 급격히 성장하여 초기 접종량의 27 배까지 성장하고 있으나, 역시 고온의 영향으로 cellular EPA 의 함량은 8.2% 수준으로 증가하는데 그치고 있다(Fig. 2).
10oC에서 배양을 할 경우 균주의 성장은 4일째에서 초기 접 종량의 38배까지 증가하여 가장 좋은 성장을 보였으나, EPA 함량의 경우 11.1%의 함량을 보였다(Fig. 2). 그러나 4oC에 서 배양 할 경우 균주의 성장은 10oC에서 배양할 때와 비슷 한 성장을 보이나, EPA의 함량의 경우 13.5%의 함량을 보 여 최적 EPA 생산 온도로 밝혀졌다(Fig 2). 따라서, 본 균주 의 최적 배양 온도는 4oC부터 10oC 사이인 것으로 밝혀졌 고, 이 온도는 남극 세종기지 근처의 연중 최고 온도에 해당 된다. 그러나, S. gelidimarina의 최적 배양 온도는 15-17oC 이며, 성장 최대 온도가 23oC이다[2]. 이런 특징은 본 연구
에서 분리된 L93 균주와 서로 다른 균주임을 보여주고 있다.
본 실험을 통하여 L93 균주는 내냉성균주(psychrotolerant) 가 아니고 호냉성균주(psychophillic) 임을 확인하였다. 호냉 성균주의 경우 저온에서 성장을 위하여 세포막의 구성 지방 산의 불포화도를 증가시켜 세포막의 유동성을 증가시키려는 경향 때문으로 알려져 있는데, 이러한 현상이 본 균주들이 EPA를 생산하는 이유를 부분적이나마 설명해주고 있다. 즉 본 균주들은 EPA를 거의 유일한 고도 불포화지방산으로 함 유하고 있는데 EPA의 생리적인 기능이 최소한 세포막의 유 동성을 변화시켜 외부 조건에 맞추어 유지하려는 목적에 사 용됨을 알 수 있다.
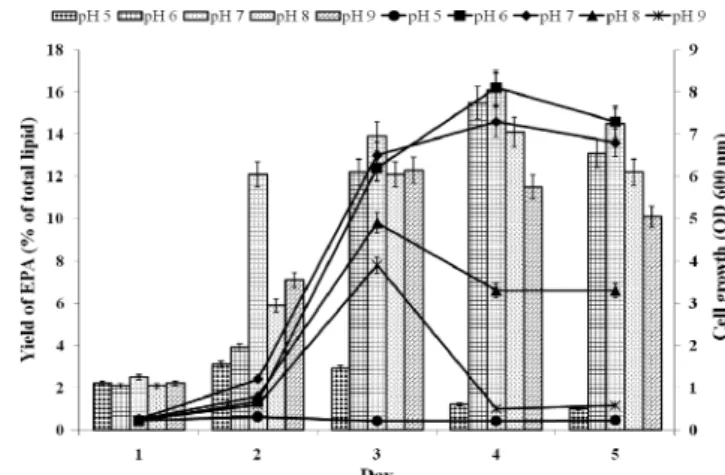
균주의 최적 초기 pH를 알아보기 위해 L93 균주를 4oC 에서 배양하며, 각각 초기 pH가 5에서 9까지 다르게 적정한 액체 배양 배지를 사용하여 최적 초기 pH를 확인하였다. 초 기 pH 5에서는 거의 균체의 성장이 나타나지 않았으며, EPA 함량 역시 초기 접종 상태를 유지하고 있어 이 조건에서는 균체가 성장하지 않고 단지 배양 온도가 낮으므로 생존만을 유지하고 있는 것으로 보였다(Fig. 3). 초기 pH 6에서 가장 빠른 균주 성장을 보였다. 배양 4일째 균주 성장이 초기 접 종량의 81배에 도달하였으며, 이후로는 감소하는 경향을 보 이고, EPA 함량은 최대 15.5%에 도달하였다(Fig. 3). pH 7 에서의 균주 성장은 pH 6과 거의 유사하며, EPA의 함량은 16.1%로 최고를 나타냈다, pH 8과 9에서의 균주 성장은 pH 6과 7에 비해 급격히 둔화되었다. EPA 함량 역시 낮아 각각 14.1%(4일째)와 12.5%(3일째)로 감소하고 있다(Fig. 3).
본 연구에서 분리된 EPA생산 균주들은 해양동물과 해양 침 적토에서 분리된 해양 세균으로서 해수 염도의 영향에 따른 EPA 생산량의 관계를 알아보았다. 해수의 농도는 sea salt를 사용하여 실험을 행하였다. L93 균주는 해수 농도 50% 수준 에서 가장 높은 성장을 보이고 있으며, EPA 함량도 15.1%로 가장 높았다(Fig. 4). 100% 해수에서는 성장과 EPA 함량에서
Fig. 2. Effect of temperature on the production of EPA by Shewanella sp. L93. EPA production: a bar graph; cell growth: a line graph.
Fig. 3. Effect of pH on the production of EPA by Shewanella sp. L93. EPA production: a bar graph; cell growth: a line graph.
50%에 비해 다소 낮은 경향을 보이고 있고, 10%와 200%의 경우 균주 성장 및 EPA 함량이 모두 현저하게 낮아지는 것을 확인하였다(Fig. 4). 위의 실험을 통해 확립된 L93 균주의 최 적 EPA 생산조건을 이용해 L93 균주의 EPA 생산량을 확인 한 결과 균주배양액 1 L 당 EPA를 320 mg 생산할 수 있었고 이 결과는 SCRC-2738과 비슷한 생산량을 나타내는 것으로 확인되었다. 이로써 본 연구를 통해 독자적인 EPA 생산 균주 를 확보하였고, 확보된 균주를 이용하여 좀 더 다양한 실험을 할 수 있는 토대를 마련하였다.
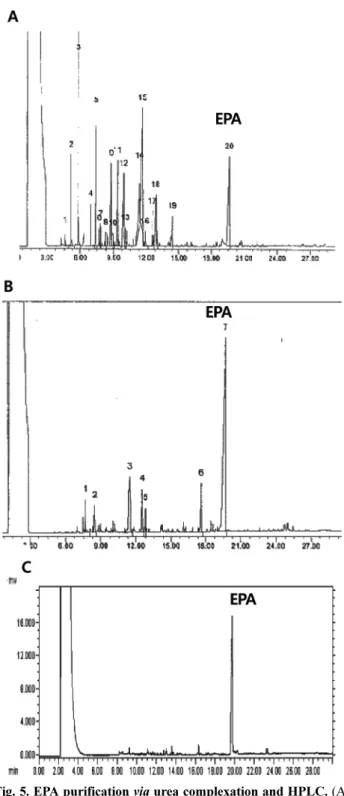
EPA 분리정제 최적화 조건 확립을 위해 위의 실험 결과 에 나타난 EPA 최적 생산 배양 조건을 사용하여 L93 균주 를 배양하였다. 배양된 균주를 재료 및 방법 내용에 나온 지 방산 추출 방법을 통하여 추출된 전체 지방산을 추출하고 GC를 사용하여 분석을 하였다(Fig 5A). Urea 냉 침전 방법 을 이용하여 1차 분리 정제를 시도하여 초기 전체 지방산 함량에 비교하여 EPA 함량을 60%로 증가시켰으며(Fig 5B), 이렇게 증가된 시료를 최종적으로 HPLC를 사용하여 정제 하여 95% 정제된 72.6 mg의 EPA를 분리하여 약 분리 정 제 수율 72% 이상인 EPA 정제 시스템을 확립하였다(Fig.
5C). 분리 정제 때마다 GC 및 HPLC의 retention time이 변 하기 때문에 EPA standard을 사용하여 목적 화합물의 분리 정제를 매 단계마다 확인하였다. 다만, Fig. 5에는 EPA standard peak를 매번 명기하지 않았다. EPA와 같은 여러 개 의 cis-형 이중결합을 포함하는 불포화 지방산의 경우 urea 냉탕침전 법을 이용하면 고도 불포화 지방산을 포화 지방산 이나 낮은 불포화도의 지방산으로부터 경제적으로 분리 정 제가 가능하다는 것을 확인하였다.
요 약
Eicosapentaenoic acid 생산 세균을 얻기 위해 1999~2000
년 하계연구 기간 중에 남극 생물 및 침적토를 사용하여 600 주의 균주를 분리하였고 TLC와 GC를 사용하여 오메가-3 고도불포화 지방산 EPA를 생산하는 미생물 7 주를 성공적 으로 분리하였으며, 이중 EPA 생산이 가장 높은 L93 균주 를 선발하였다. 16S rDNA의 염기서열 분석을 통하여 Fig. 4. Effect of sea salt concentration on the production of
EPA by Shewanella sp. L93. EPA production: a bar graph; cell growth: a line graph.
Fig. 5. EPA purification via urea complexation and HPLC. (A) Total fatty acid ethyl ester analysis of Shewanella sp. L93 by gas chromatography. (B) GC chromatogram of Fatty acid methyl ester after urea complexation. (C) GC chromatogram of purified EPA methyl eater after HPLC purification.
Shewanella 속으로 조사되었으며, 이에 분리된 균주를 Shewanella sp. L93라 명명하였다. EPA를 생산 최적 배양온 도 4oC 이며, 초기 pH 7에서 최적 EPA 함량을 보였다. 아 울러 염 농도는 50 %(w/v)에서 생산이 최대였다. EPA 최적 생산 조건을 이용하여 리터당 320 mg을 생산할 수 있는 생 산 시스템을 확립하였다. Urea 침전법과 HPLC을 이용하여 수율 72% 이상의 97% 순도를 가진 EPA를 정제할 수 있는 분리 정제 시스템 또한 본 연구를 통하여 확립하였다.
REFERENCES
1. Bang, H. O. and J. Dyerberg. 1972. Plasma lipids and lipoproteins in Gleenlandic west coast Eakimos. Acta Med.
Scand. 192: 85-94.
2. Bowman, J. P., S. A. McCammon, D. S. Nichols, J. H.
Skerratt, S. M. Rea, P. D. Nichols, and T. A. McMeekin.
1997. Shewanella gelidimarina sp. nov. and Shewanella frigidimarina sp. nov., novel Antarctic species with the ability to produce eicosapentaenoic acid (20:5 omega 3) and grow anaerobically by dissimilatory Fe(III) reduction. Int. J.
Syst. Bacteriol. 47: 1040-1047.
3. Cho, K. W. and S. J. Mo. 1999. Screening and characteri- zation of eicosapentaenoic acid-producing marine bacteria.
Biotechnol. Lett. 21: 215-218.
4. Christie, W. W. 1982. Lipid Analysis, Pergamon Press.
London.
5. Connor, S. L. and W. E. Conner. 1997. Are fish oils bene- fical in the prevention and treatment of coronary artery disease? Am. J. Clin. Nutr. 66 (suppl): 1020-1031.
6. Dyerberg, J. and H. O. Bang. 1979. Lipid metabolism, atherogenesis, and haemostasis in Eskimos: the role of the prostaglandin-3 family. Haemostasis 8: 227-233.
7. Dyeberg, J., H. O. Bang, E. Stoffersen, S. Moncada, and J.
R. Vane. 1978. Eicosapentaenoic acid and prevention of
thrombosis and atherosclerosis? Lancet ii: 117-119.
8. Kelly, J. F. 1991. The metabolic role of n - 3 polyunsaturated fatty acids: relationship to human disease. Comp. Biochem.
Physiol. A Comp. Physiol. 98: 581-585.
9. Lane, D. J. 1991. Nucleic acid techniques in bacterial syste- matic, pp. 115-175. John Wiley and Sons, New York, U.S.A.
10. Lee, S. J., P. S. Seo, C. H. Kim, O. Kwon, B. K. Hur, and J.
W. Seo. 2009. Isolation and characterization of the eicosa- pentaenoic acid biosynthesis gene cluster from Shewanella sp. BR-2. J. Microbiol. Biotechnol. 19: 881-887.
11. Lee, W. H., K. W. Cho, S. Y. Park, K. S. Shin, D. S. Lee, S.
K. Hwang, S. J. Seo, J. M. Kim, S. Y. Ghim, B. H. Song, S.
H. Lee, and J. G. Kim. 2008. Identification of psychrophile Shewanella sp. KMG427 as an eicosapentaenoic acid pro- ducer. J. Microbiol. Biotechnol. 18: 1869-1873.
12. Lopoz Alonso, D., E. Molina Grima, J. A. Sunclez Perez, J.
L. Gracia Sanchez, and F. Gracia Camacho. 1992. Fatty acid variation among different isolates of a single strain of Isochrysis galbana. Phytochemistry 31: 3901-3904.
13. Pettinella, C., S. H. Lee, F. Cipollone, and I. A. Blair. 2007.
Targeted quantitative analysis of fatty acids in atherosclerotic plaques by high sensitivity liquid chromatography/tandem mass spectrometry. J. Chromatogr. B 850: 168-176.
14. Salunkhe, D., N. Tiwari, S. Walujksr, and R. Bhadekar.
2011. Halomonas sp. nov., an EPA-producing mesophilic marine isolate from the Indian Ocean. Pol. J. Microbiol. 60:
73-78.
15. Taketama, H., K. Iwamoto, S. Hata, H. Takno, and T.
Matsunaga. 1996. DHA enrichment of rotifers: A simple two-step culture using the unicellula algae chlorella regularis and Isochrysis galbana. J. Mar. Biotechnol. 3: 244-247.
16. Yazawa, K., K. Araki, N. Okazaki, K. Watanabe, C. Ishikawa, A. Inoue, N. Numao, and K. Kondo. 1988. Production of eicosapentaenoic acid by marine bacteria. J. Biochem. 103:
5-7.