Roles of miR-128 in Myogenic Differentiation and Insulin Signaling in Rat L6 Myoblasts
Myung-Ju Oh
2, So-Hyeon Kim
1, Ji-Hyun Kim
1and Byung H. Jhun
1*
1College of Nanoscience and Nanotechnology, Pusan National University, Busan 46241, Korea
2Ministry of Food and Drug Safety, Chungbuk 28159, Korea
Received August 19, 2020 /Revised September 4, 2020 /Accepted September 7, 2020
Skeletal muscle differentiation or myogenesis is important to maintain muscle mass and metabolic homeostasis. Muscle-specific microRNAs (miRNAs) are known to play a critical role in skeletal myo- genic differentiation. In this study, we examined the expression profiling of miRNAs during myogenic differentiation in rat L6 myoblasts using rat miRNA microarrays. We identified the upregulated ex- pression of miR-128 as well as several well-known myogenic miRNAs, including miR-1, miR-133b, and miR-206. We additionally confirmed the increased expression of miR-128 observed on microarray through quantitative real-time PCR (qRT-PCR), which showed similarly upregulated expression of both primary miR-128 and mature miR-128, consistent with the microarray findings. Furthermore, transfection of miR-128 into rat L6 myoblasts induced gene expression of myogenic markers such as muscle creatine kinase (MCK), myogenin, and myosin heavy chain (MHC). Protein expression of MHC was increased as well. Inhibition of miR-128 by inhibitory peptide nucleic acids (PNAs) blocked the expression of those myogenic markers. In addition, the transfection of miR-128 into rat L6 myo- blasts enhanced the phosphorylation of Erk and Akt proteins stimulated by insulin, while simulta- neously reversing the inhibited phosphorylation of Erk and Akt due to insulin resistance. These find- ings suggest that miR-128 may play important roles in myogenic differentiation and insulin signaling.
Key words : Insulin signaling, L6 myoblasts, microRNA, miR-128, muscle differentiation
*Corresponding author
*Tel : +82-51-510-5117, Fax : +82-51-514-2358
*E-mail : [email protected]
This is an Open-Access article distributed under the terms of the Creative Commons Attribution Non-Commercial License (http://creativecommons.org/licenses/by-nc/3.0) which permits unrestricted non-commercial use, distribution, and reproduction in any medium, provided the original work is properly cited.
Introduction
MicroRNAs (miRNAs or miRs) belong to non-coding RNAs composed of approximately 22 nucleotide sequences [2, 7]. miRNAs have been shown to be an important coor- dinator in a wide spectrum of biological processes and dis- eases including myogenesis and various musculoskeletal disorders [13, 23]. miRNAs are first synthesized into the pri- mary miRNAs by RNA polymerase II in the nucleus, then cleaved into 60-70 nucleotide precursor miRNAs by RNase III Drosa and its cofactor, DiGeorge syndrome chromosomal region 8 (DGCR8), which are transported into the cytosol by exportin 5. There, the precursor miRNAs are digested by RNase III Dicer and assembled with the argonaute (AGO) protein, resulting in mature miRNAs with 22 nucleotides.
AGO proteins loaded with an individual mature miRNA are
referred to as the RNA-induced silencing complex (RISC).
The binding of these mature miRNAs in the RISC to the complementary target mRNA induces the degradation of their cognate mRNA resulting in the reduction of protein translation [8].
Skeletal muscle consists of approximately 40% of human body mass and continuously regenerates through myo- genesis. Myoblast differentiation from mesodermal stem cells is a highly regulated process in myogenesis [4]. The formation of myocytes and final myofibers from myoblasts are controlled by myogenic regulatory transcription factors such as MyoD, Myf5, myognein and myogenic regulatory factor 4 (MRF4) [3]. Each myogenic transcription factor coor- dinates a specific process within myoblast differentiation.
The final steps are regulated by MRF4 and myosin heavy chain (MHC), which are responsible for muscle fiber for- mation and function.
It is of importance to identify myogenic miRNAs and un-
derstand functional roles of miRNAs in skeletal muscle de-
velopment and musculoskeletal disorders. Several miRNAs
are shown to be exclusively and preferentially expressed in
skeletal and cardiac muscles: miR-1, miR-133a, miR-133b,
miR-206, miR-208a, miR-208b and miR-499 [13]. Among
these, miR-206 and miR-208a are specifically expressed in skeletal muscle and cardiac muscle, respectively. Other miRNAs that have shown to be expressed not only in mus- cles but also other tissues including various cancer cells in- clude: miR-24, miR-29, miR-125, miR-181, miR-214, miR- 221/222, miR-322/424, miR-486, miR-503 and miR-675 [9, 25, 31]. These myogenic miRNAs are currently under intense investigation to understand their roles in musculoskeletal physiology and pathophysiology.
Skeletal muscle is an important target of glucose homeo- stasis. During myogenesis, insulin receptor expression is in- creased and insulin responsiveness is also upregulated [21].
Some miRNAs regulate the signal transduction pathway of insulin [18]. Impaired insulin signaling in skeletal muscle has been shown to be associated with insulin resistance and metabolic syndromes, including type II diabetes [10]. Several miRNAs including miR-27a, miR-106a and miR-30d are known to be involved in muscle-related insulin resistance [32, 34].
In this study, we examined expression of miRNAs during myogenic differentiation and found that miR-128 is upregu- lated during myogenic differentiation in rat-L6 myoblasts.
We also observed the up-regulation of well-known myo- genic miRNAs, such as miR-1, miR-133b and miR-206. The transfection of miR-128 induced the myogenic differentiation of rat L6 myoblasts whereas the inhibition of miR-128 by inhibitory PNA-128 blocked the differentiation. We also found that miR-128 is involved in insulin signaling and in- sulin resistance. These results suggest that miR-128 plays a role in myogenic differentiation and insulin signaling.
Materials and Methods
Materials
Antibodies against phospho-Akt (Ser-473), Akt, phospho- Erk1/2 (Thr202/Tyr204), Erk1/2, phospho-S6K (Thr-389), S6K, IRS-1, IRS-2 and GAPDH were obtained from Cell Signaling Technology, Inc. (USA). miRNAs were purchased from Bioneer (Korea). PNAs, inhibitory miRNAs, against miR-1 and miR-128 were from PANAGENE (Korea). Antibody against MHC protein was obtained from Developmental Studies Hybridoma Bank (USA). Horseradish peroxidase conjugated anti-rabbit or anti-mouse secondary antibodies and Oligofectamine were obtained from GE Healthcare Life Sciences (UK). Alexa Fluor 488 goat anti-mouse antibody, Hoechst 33342, mirVana miRNA Isolation Kit, Superscript
III and random primer, Platunum SYBR Green qPCR Super Mix-UDG and NCode miRNA first-strand cDNA synthesis and qRT-PCR Kits were from Thermo Fisher Scientific (USA).
All other reagents were from Sigma-Aldrich (USA).
Cell culture and RNA extraction
Rat L6 myoblasts were directly obtained from American Type Culture Collection (ATCC, USA), maintained and dif- ferentiated as described previously [15]. Briefly, rat L6 myo- blasts were maintained in growth medium (GM) and differ- entiated by serum-deprived differentiation medium (DM).
Total RNA was extracted from rat L6 myoblasts at the in- dicated day by using the mirVana miRNA Isolation Kit ac- cording to the manufacturer’s protocol. RNA concentration was quantified with a Nano Drop spectrophotometer, and RNA integrity was evaluated using Agilent 2100 Bioanalyzer (Agilent, USA).
Analysis of miRNA microarray
For miRNA microarray analysis, 100 ng of total RNA sam- ples were labeled with pCp-Cy3 and hybridized to 8x15k Agilent Rat miRNA microarrays (Agilent, USA) for 20 hr at 55℃ according to the manufacturer's instructions. The ar- ray was scanned using the Agilent microarray scanner (G2565 AA). The total signals were extracted as "GeneView"
data files using Agilent Feature Extraction software.
Differentially expressed miRNAs (DEMirs) were identified by applying linear model with “limma” package in Bioconductor.
Analysis of primary and mature miRNAs by quan- titative real-time PCR
For analysis of primary miRNAs expression, 1 μg of total RNA was used as the template for first-strand cDNA syn- thesis with Superscript III and random primer according to the manufacturer’s protocol. Then, qRT-PCR was performed with the 7500 Real-Time PCR System (Applied Biosystems) with Platunum SYBR Green qPCR SuperMix-UDG. Rat U6 was used for normalization, and relative amounts of the tar- get were calculated using the comparative Ct method (2
-DDCt).
For analysis of mature miRNAs expression, qRT-PCR was performed using NCode miRNA first-strand cDNA syn- thesis and qRT-PCR Kits according to the manufacture’s protocol. PCR cycling conditions were 95℃ for 15 min and 45 cycles (95℃ for 10 s, 55℃ for 15 s, and 72℃ for 30 s).
The primer sequences used for qRT-PCR are listed in Table
Table 1. Primers used for qRT-PCR
Gene name Primer sequence (5’-3’)
Rno-miR-1
Mature TGGAATGTAAAGAAGTGTGTAT
Precursor F: TGCCTACTCAGAGCACATACTTCT
R: CACTTCTTTACATTCCATAGCATTC
Rno-miR-128
Mature TCACAGTGAACCGGTCTCTTT
Precursor F: GGGCCGTAGCACTGTCTGA
R: AAGCAGCTGAAAAAGAGACCG
Rno-miR-133b
Mature TTTGGTCCCCTTCAACCAGCTG
Precursor F: TAAAATGGAACCAAATCGCCT
R: CTGGTTGAAGGGGACCAAAT
Rno-miR-206
Mature TGGAATGTAAGGAAGTGTGTGG
Precursor F: CCAGGCCACATGCTTCTTTA
R: TTCCTTACATTCCATAGCGCA
MCK F: GAGTCCTACACGGTCTTCAAG
R: CAGGAAGTGGTCATCAATGAG
MHC F: GAAGGCCAAGAAGGCCATC
R: CTCGCCTCGTGTTTCG
Myogenin F: GCAGTGCCATCCAGTACATTGAGC
R: GGAAGGTGACAGACATATCCTCCAC
1. The mature miRNA-universal reverse primer supplied in
the NCode miRNA qRT-PCR kit was used for analysis of mature miRNAs.
Transfection of miRNAs/peptide nucleic acids (PNAs) and analysis of myogenic markers by quantitative real-time PCR
L6 myoblasts were transfected with 0.02 μM of either dou- ble-stranded mature miRNAs such as miR-1, miR-128 and control miR-125b-5p or inhibitory PNA against miR-1, miR- 128 and control miR-125p-5p for 4 hr using Oligofectamine according to the manufacturer’s protocol. The sequences of oligonucleotides used in this study were miR-1 (5¢-uggaau- guaaagaaguguguau-3¢), miR-128 (5¢-ucacagugaaccggucucuuu- 3¢) and control miR-125b-5p (5¢-ccuacgccaccaauuucgu-3¢) and PNAs containing antisense sequence against miR-1, miR-128 and control miR-125b-5p. The miRNAs- or PNAs- transfected L6 myoblasts were incubated in GM for 1 day and further differentiated with DM for 3 days. The trans- fected L6 cells were harvested and performed for qRT-PCR as described previously [19] . The primer sequences used for Art-PCR are listed in Table 1. Data were analyzed by a comparative cycle threshold method in Bio-Rad iQ5 software.
Immunoblotting and immunostaining of MHC pro- tein expression
L6 myoblasts transfected with either miRNAs or inhibitory PNAs were incubated in the same condition as that for qRT- PCR, lysed and immunoblotted with specific antibodies against MHC, p-Erk, Erk, p-Akt, Akt, IRS-1, IRS-2 and GAPDH. The band intensity was quantified as previously de- scribed [15]. For immunostaining of MHC protein, the cells were cultured on acid washed coverslips and transfected with miRNAs and PNAs as described above. The transfected cells were immunostained with anti-MHC antibody and im- aged with a confocal microscope (Carl Zeiss, Jena, Germany) as previously described [15]. The number of cells stained with anti-MHC antibody was quantified for every 1,000 cells per coverslip.
Insulin stimulation and induction of insulin resistance
For the stimulation of L6 myoblasts by insulin, L6 cells
were transfected with miRNA as above, differentiated for
3 days, starved with serum withdrawal for 2 hr, and then
stimulated with insulin (100 ng/ml) for 5 min. Whole cell
lysates were subject to immunoblotting analysis with anti-
bodies against p-Erk, Erk, p-Akt, Akt, and GAPDH. For the
induction of insulin resistance, the miRNA-transfected L6
myoblasts were incubated in the presence of high insulin
(1 μg/ml) and high glucose (25 mM) for 3 days and the cell
lysates were immunoblotted with specific antibodies. The band intensity was quantified as previously described [15].
Statistical analysis
Results are obtained from three independent experiments.
Data are expressed as means ± standard deviation (SD). The results were statistically analyzed by one-way ANOVA method and the statistical significance was determined by the Fischer’s Protected LSD post-hoc test. P-values <0.05 was considered statistically significant.
Results
Expression profiling of miRNA during differentiation of rat L6 myoblasts
Expression profiling of miRNA during muscle differ- entiation in mouse C2C12 myoblasts has been reported [5].
However, the expression profiling in rat L6 myoblasts has yet to be reported. To examine the miRNA expression in rat L6 myoblasts, we first established the differentiation con- dition of L6 cells by examining the expression of MHC as a differentiation marker as previously reported by us [15].
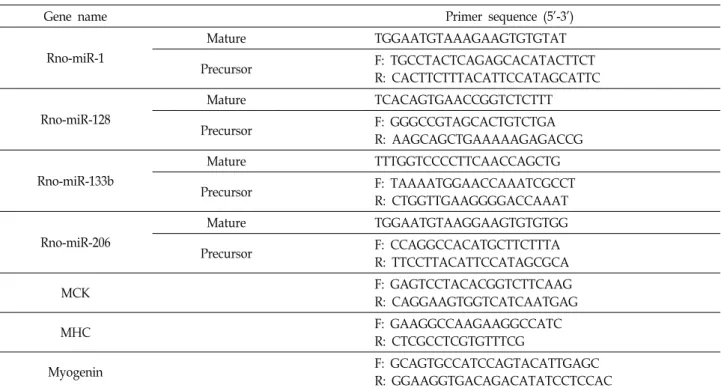
As shown in Fig. 1A, L6 cells gradually differentiated and formed multinucleated myocytes with increasing expression of MHC protein, a typical myogenic differentiation marker.
Next, we examined the expression level of miRNAs dur- ing differentiation of rat L6 myoblasts at day 0, 1, 3 and 6 using a rat miRNA array. We found that some of miRNAs were upregulated or downregulated during differentiated L6 myoblasts (Fig. 1B, upper panel). Among upregulated miRNAs, we specifically plotted the expression of miR-1, miR-128, miR-133b and miR-206 according to the time course of differentiation. These four miRNAs were significantly in- creased during differentiation (Fig. 1B).
In order to confirm the upregulated expression of miR-1, miR-128, miR-133b and miR-206, we examined the expre- ssion level of both primary miRNA and mature miRNAs of these miRNAs on day 0 and day 6 after differentiation by qRT-PCR (Fig. 1C). We used miR-125b-5p as the control miRNA, as it showed constant expression on the array dur- ing differentiation. Similar to array results, we observed the upregulation of the expression of these four miRNAs in the level of both primary miRNA and mature miRNA during differentiation. The array and qRT-PCR results showed that the expression of miR-1, miR-128, miR-206 and miR-133b was significantly increased during the myogenic differ-
entiation of rat L6 myoblasts. It has been reported that miR-1, miR-133b and miR-206 were increased during differ- entiation of mouse C2C12 myoblasts [5]. However, the levels of miR-128 expression during myogenic differentiation has not been characterized.
Induction of myogenic differentiation by miR-128 in rat L6 myoblasts
Next, we investigated function roles of miR-128 and miR-1 in muscle differentiation using rat L6 myoblasts. To do this, we transfected miR-128 and examined the induction of myogenic markers such as muscle creatine kinase (MCK), myogenin, and myosin heavy chain (MHC). As shown in Fig. 2A, transfecting miR-128 significantly induced gene ex- pression of MCK, myogenin and MHC as quantified by qRT-PCR. Similarly, we confirmed this induction of MHC protein expression using Western blot analysis (Fig. 2B) and immunostaining analysis (Fig. 2C). We also observed similar myogenic effects of miR-1, with no significant difference be- tween the effect of miR-128 and miR-1 on the induction of MHC expression. We used miR-125b-5p as the control miRNA. These results suggest that miR-128 has the ability to induce myogenic differentiation in rat L6 myoblasts.
Inhibition of myogenic differentiation by inhibitory PNA-128 in rat L6 myoblasts
In order to confirm the functional role of miR-128 in myo- genic differentiation, we then utilized the inhibitory peptide nucleic acid (PNA) against miR-128. PNA is an artificial oli- gonucleotide with a peptide backbone that has specific bind- ing affinity to its target miRNAs, which confers a highly effective, long-lasting inhibition of miRNAs [24]. Transfec- tion of inhibitory PNA-128 reduced the expression of mRNA of MHC, but not those of myogenin and MCK (Fig. 3A).
The transfected PNA-128 significantly suppressed the ex- pression of MHC protein in both western blot analysis (Fig.
3B) and immunostaining analysis (Fig. 3C). We also ob- served similar inhibitory effects of PNA-1 and there was no significant difference of the effect between PNA-128 and PNA-1. We used PNA-125b-5p as the control PNA. Taken together, these results suggest that miR-128 plays an im- portant role in myogenic differentiation in rat L6 myoblasts.
Functional role of miR-128 in insulin signaling and insulin resistance
Skeletal muscle is an important target of insulin action
A B
C
Fig. 1. Expression of microRNAs during myogenic differentiation in L6 cells. (A) L6 myoblasts were induced to differentiate for the indicated days. The differentiated myocytes were identified by immunostaining myosin heavy chain (MHC) (green) as a differentiation marker. Nuclei (blue) were stained with Hoechst 33342. (B) Heat map of miRNA expression during L6 differentiation. Total RNAs from differentiated L6 myoblasts on day 0, 1, 3, 6 were prepared and subjected to miRNA micro- array analysis on the Agilent rat miRNA array. Differentially-expressed miRNAs expressed in undifferentiated (GM) and differentiated (DM) L6 cells are shown in a heat map representation. Upregulated miRNAs and downregulated miRNAs were labeled with red and green, respectively. Expression patterns of miR-11, miR-128, miR-206 and miR-133b were compared at the bottom (three independent replicates, *p<0.05). Results are obtained from three independent microarray experiments.
Values from undifferentiated L6 cells are set to 1. Bars, the mean result ± SD. *p<0.05 relative to control undifferentiated L6 cells. (C) qRT-PCR analysis of the upregulated miRNAs. To validate the result of miRNA microarray, qRT-PCR analysis of mature miRNAs (upper panel) and primary miRNAs (lower panel) was performed with control undifferentiated and differentiated L6 cells on day 6. Rat U6 small nuclear RNA was used for normalization. Results are obtained from three independent experiments and are expressed as the fold increase related to control undifferentiated L6 cells. Bars, the mean result ± SD. *p<0.05 relative to control undifferentiated L6 cells.
to regulate the level of blood glucose. Rat L6 myoblasts are commonly used as a model cell for examining insulin actions and insulin resistance. We examined functional role of miR-
128 in insulin actions. To do these, we transfected miR-128
into rat L6 myoblasts, allowed 3 days for differentiation, and
then examined their effects on insulin actions such as activa-
A B C
Fig. 2. Induction of myogenic differentiation by miR-128 in rat L6 myoblasts. L6 myoblasts were transfected with double-stranded mature miRNAs of either miR-128, miR-1 or control miR-125b-5p. The cells were further incubated for 3 days. (A) Total RNA was prepared from the transfected cells and RT-PCR was performed using primers specific for muscle creatine kinase (MCK), myogenin, myosin heavy chain (MHC) and GAPDH. Band intensities were quantified using ImageJ software, and the relative intensities of three independent experiments are presented as mean ± SD. Values from L6 cells transfected with control miR-125b-5p are set to 1. *p<0.05 relative to control miRNA-transfected L6 cells. (B) The cells were lysed and immuno- blotted with anti-MHC antibody. Band intensities were quantified as in (A). Results are obtained from three independent experiments. Values from L6 cells transfected with control miR-125b-5p are set to 1. (C) The cells transfected with miRNAs were fixed on day 0 and day 3, and immunostained with anti-MHC antibodies (green) and Hoechst dye (blue). The 1,000 L6 cells stained with Hoechst were counted and in the same field of the microscope, the number of cells immunostained with anti-MHC antibody were also counted. Values of cells immunostained with anti-MHC antibody are obtained from three independent experiments. Bars, the mean result ± SD. *p<0.05 relative to control cells transfected with miR-125b-5p.
tion of downstream signaling proteins, such as Erk and Akt.
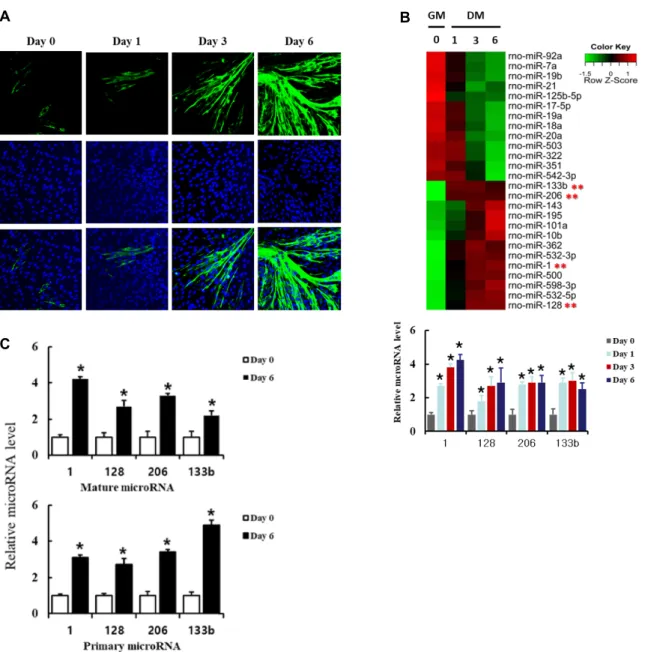
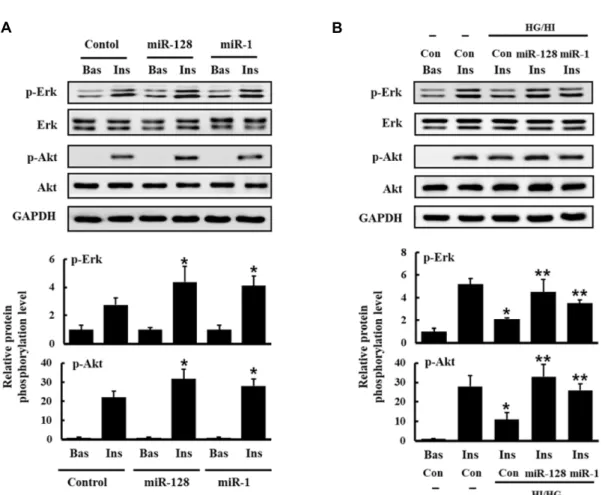
As shown in figure 4A, the transfected miR-128 significantly enhanced insulin-stimulated phosphorylation of both Erk and Akt proteins, suggesting that miR-128 potentiates in- sulin actions.
Next, we examined the effect of miR-128 on insulin resistance. Insulin resistance, a defect in insulin actions, is a key factor in type II diabetes and metabolic syndromes.
To do this, insulin resistance in rat L6 myoblasts were in- duced by treatment with high concentrations of glucose and insulin. L6 myoblasts transfected with miR-128 were differ- entiated in the presence or absence of both high glucose (25 mM) and high insulin (1 μg/ml) for 3 days. Insulin actions were examined as described above. Insulin-stimulated acti- vation of Erk and Akt proteins was significantly suppressed by treatment with both high glucose and high insulin, while the transfection of miR-128 restored insulin-stimulated acti-
vation of both Erk and Akt proteins (Fig. 4B). We also ob- served similar effects with miR-1 transfection and there was no significant difference in the restoration of Erk and Akt activation between miR-128 and miR-1. Taken together, these results suggest that miR-128 is involved in both insulin signaling and insulin resistance. We used miR-125b-5p as the control miRNA.
Inhibition of expression of IRS-1 protein by miR- 128 in rat L6 myoblasts
In order to understand the molecular mechanism under-
lying the regulation of myogenic differentiation and insulin
action, we used TargetScan to predict potential targets of
miR-128. Among possible target genes, we found that IRS-1
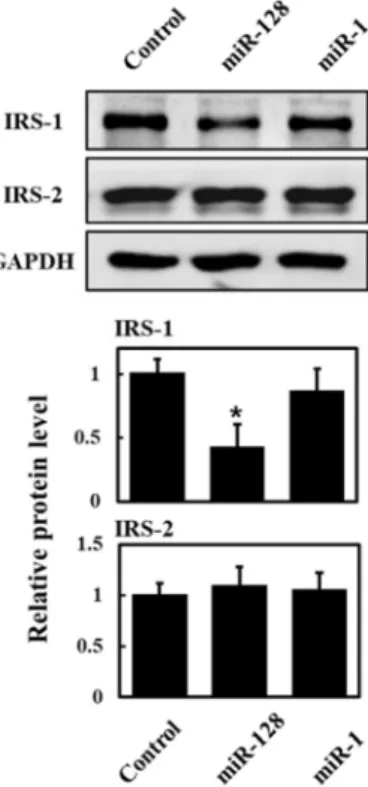
has a binding site for miR-128. As shown in Fig. 5, over-
expression of miR-128 suppressed the expression of IRS-1
protein, but not IRS-2 protein. In comparison, miR-1 did not
A B C
Fig. 3. Inhibition of myogenic differentiation by inhibitory PNA-128 in rat L6 myoblasts. Undifferentiated L6 myoblasts in 50%
confluence were transfected with inhibitory PNA-128, PNA-1 and control PNA-125b-5p for 1 day, and then the cells were induced to differentiate by differentiation medium for 3 days. The effects of inhibitory PNA-128 and miR-1 were examined by RT-PCR (A), immunoblotting (B) and immunostaining (C) as in Fig. 2. Values are obtained from three independent experiments. Bars, the mean result ± SD. *p<0.05 relative to control cells transfected with PNA-125b-5p.
affect the expression of IRS-1 protein and IRS-2 protein.
These results suggest that miR-128 may not only regulate myogenic differentiation, but also insulin action in rat L6 myoblasts. Further investigation of the underlying molecular mechanism of IRS-1 regulation through miR-128 is war- ranted.
Discussion
Accumulating evidence suggests that miRNAs are im- portant players in myogenic processes [13]. A number of miRNAs are shown to be specifically expressed in cardiac and/or skeletal muscle: miR-1, miR-133a, miR-133b, miR- 206, miR-208a, miR-208b, miR-486 and miR-499. In the pres- ent study, we observed the upregulation of a subset of these miRNAs, miR-1, miR-133b and miR-206, during myogenic differentiation of rat L6 myoblasts using miRNA array fol- lowed by qRT-PCR techniques. We additionally found in- creased expression of miR-128 in rat L6 myoblasts. The ex- pression pattern of miR-128 both in the form of primary and mature miRNA was similar to those of other myogenic miRNAs, such as miR-1, miR-133b and miR-206. We also
found that miR-128 itself has an ability to induce myogenic differentiation when overexpressed. Furthermore, we dem- onstrated that peptide-derived inhibitory PNA-128, which blocks the long-lasting action of the endogenous miRNA equivalent to miR-128, suppresses myogenic differentiation.
These findings strongly suggest that miR-128 plays a myo- genic role in the differentiation of rat L6 myoblasts.
Previous studies reported upregulation of miR-1, miR-133
and miR-206 during skeletal muscle development [5, 13], but
not that of miR-128. These studies utilized mouse-originated
C2C12 myoblasts in order to investigate the expression
profiling and functional roles of miRNAs. We made use of
rat-originated L6 myoblasts for the study of miRNA array
and additionally found the induction of miR-128 during
differentiation. Such discrepancy may arise from differences
in cell lines and differentiation conditions. Although mouse
C2C12 and rat L6 myoblasts are both frequently used for
investigating myogenesis and insulin-regulated metabolic
actions, there are notable differences between these cell lines
[1]. The comparison of transcriptomic and metabolic profiles
between rat L6 myoblasts and mouse C2C12 myoblasts
showed significant heterogeneity. Particularly, L6 myoblasts
A B
Fig. 4. Expression of miR-128 regulates insulin signaling and insulin resistance in L6 cells. (A) L6 myoblasts were transfected with miR-128, miR-1 and control miR-125b-5p, and then differentiated for 3 days. The cells were serum-starved for 4 hr and stimulated with 100 ng/ml of insulin for 5 min. The cells were then lysed and immunoblotted with antibodies against p-Akt, Akt, p-Erk, Erk and GAPDH. Band intensities were quantified using ImageJ software, and the relative intensities of three independent experiments are presented as mean ± SD. Values from L6 cells transfected with control miR-125b-5p are set to 1. *p<0.05 relative to control miR-125b-5p-transfected and insulin-stimulated L6 cells. (B) To examine a role of miR-128 in insulin resistance, L6 myoblasts were transfected with miR-128, miR-1 and control miR-125b-5p, and then differentiated for 3 days in the presence of high glucose (25 mM) and high insulin (1 μg/ml). The serum-starved cells were treated with insulin, immunoblotted and analyzed as above. Values from L6 cells transfected with control miR-125b-5p are set to 1. *p<0.05 relative to control miR-125b-5p-transfected and insulin-stimulated L6 cells. **p<0.05 relative to control miR-125b-5p-transfected L6 cells stimulated with insulin in the presence of high glucose and high insulin.
are more suitable for studies of insulin-regulated glucose transport and metabolism, while C2C12 myoblasts are more appropriate for studies of exercise and stress response. These cell-specific responses are dependent on transcriptomic dif- ferences between C2C12 and L6 myoblasts. We previously reported such a discrepancy in the behavior between mouse C2C12 myoblasts and rat L6 myoblasts in the regulation of myogenic differentiation by β
2-adrogenic receptor [15]. These previous findings may explain the unique upregulation of miR-128 expression and induction of myogenic differ- entiation by miR-128 in rat L6 myoblasts. Further studies are needed to elucidate the source of these discrepancies.
It has been reported that miR-128 is a key regulator in
tumorigenesis and differentiation of stem cells. Expression
of miR-128 was downregulated in several cancers including
prostate cancer, glioma, head and neck squamous carcino-
ma, non-small cell lung cancer, and colorectal carcinoma [6,
12, 14, 30]. Increased expression of miR-128 suppressed pro-
liferation of pancreatic cancer cells, colorectal cancer and
breast cancer [11, 16]. The level of blood circulating miR-128
was increased in glioma and lung cancer [17, 20]. In addi-
tion, miR-128 is known to play a regulatory role in the differ-
entiation of stem cells, including the differentiation of hu-
man hair follicle mesenchymal stem cells into smooth mus-
cle cells [29], bone marrow stem cells into neurons [30] and
osteoblasts [28], and neuronal stem cells into neuron [33].
Fig. 5. Transfection of miR-128 suppresses expression of IRS-1 protein in L6 cells. L6 myoblasts were transfected with miR-128, miR-1 and control miR-125b-5p, and then dif- ferentiated for 3 days. The cells were lysed and immuno- blotted with antibodies against IRS-1, IRS-1 and GAPDH.
Band intensities were quantified using ImageJ software, and the relative intensities of three independent experi- ments are presented as mean ± SD. Values from L6 cells transfected with control miR-125b-5p are set to 1. *p<0.05 relative to control microRNA-transfected L6 cells.
Nonetheless, the exact functional role of miR-128 in myo- genic differentiation is not well understood.
A number of miRNAs are known to be involved in insulin signaling and insulin resistance [18]. Insulin receptor and its signaling proteins including IRS-1/2, PTEN, PI3-kinase, Akt and others are known targets of miRNAs. Recently, miRNAs have become a potential therapeutic target to treat type II diabetes, a condition characterized by insulin resist- ance and impaired insulin signaling [22]. We also observed functional roles of miR-128 in insulin signaling and insulin resistance. miR-128 transfection potentiated insulin-induced phosphorylation of Erk protein and Akt protein and recov- ered insulin resistance, which was modeled by conditions of high insulin and high glucose.
Our results demonstrated that overexpression of miR-128 reduced the expression of IRS-1, not that of IRS-2, in L6 myoblasts. The IRS proteins are a family of cytoplasmic adaptor proteins that transmit signals from the insulin re-
ceptor and insulin-like growth factor (IGF)-1 receptor to reg- ulate numerous cellular responses [27]. IRS proteins have six members, IRS-1 through IRS-6, and each IRS protein has four isoforms, which have tissue-specific patterns of ex- pression and unique/interchangeable functions. IRS-1 and IRS-2 are ubiquitously expressed, whereas IRS-3 is specifi- cally expressed in the liver and IRS-4 in the brain. IRS-1 tends to play a more important role in growth and pro- liferation. IRS-2 protein has been shown to substitute IRS-1- mediated subcellular signaling of insulin and IGF-1 in the knock-out mouse of IRS-1 [27]. Recently, it has been reported that IRS-1 is a direct target of miR-128 that results in the suppression of proliferation of colorectal carcinoma [30]. Our observation may suggest that suppression of the growth sig- nal of IRS-1 by miR-128 downregulates signals leading to cell proliferation and instead induces differentiation of L6 myoblasts. It is also possible that miR-128 may switch the metabolic signal of IRS-1 to IRS-2, leading to the potentiation of insulin signaling to activate Erk and Akt and induce the recovery of insulin resistance. Target gene analysis of miR- 128 including IRS-1 and its subcellular function is currently under investigation.
In summary, we have shown that miR-128 induces and regulates myogenic differentiation in rat L6 cells myoblasts.
Furthermore, we demonstrated that miR-128 plays a role in insulin signaling and insulin resistance. Future research is needed to further characterize the molecular mechanisms underlying the role of miR-128 in myogenic differentiation and insulin actions.
Acknowledgement
This work was supported by a 2-Year Research Grant of Pusan National University.
The Conflict of Interest Statement
The authors declare that they have no conflicts of interest with the contents of this article.
References
1. Abdelmoez, A. M., Sardón Puig, L., Smith, J. A., Gabriel, B.
M., Savikj, M., Dollet, L., Chibalin, A. V., Krook, A., Zierath, J. R. and Pillon, N. J. 2020. Comparative profiling of skeletal muscle models reveals heterogeneity of transcriptome and metabolism. Am. J. Physiol. Cell Physiol. 318, C615-C626.
2. Bartel, D. P. 2004. MicroRNAs: genomics, biogenesis, mech- anism, and function. Cell 116, 281-297.
3. Cervelli, M., Leonetti, A., Duranti, G., Sabatini, S., Ceci, R.
and Mariottini, P. 2018. Skeletal muscle pathophysiology: The emerging role of spermine oxidase and spermidine. Med.
Sci. 6, 14.
4. Chal, J. and Pourquié, O. 2017. Making muscle: skeletal myogenesis in vivo and in vitro. Development 144, 2104-2122.
5. Chen, J. F., Mandel, E. M., Thomson, J. M., Wu, Q., Callis, T. E., Hammond, S. M., Conlon, F. L. and Wang, D. Z. 2006.
The role of microRNA-1 and microRNA-133 in skeletal mus- cle proliferation and differentiation. Nat. Genet. 38, 228-233.
6. Cui, J., Zhao, Y., Sethi, P., Li, Y., Mahta, A., Culicchia, F.
and Lukiw, W. 2010. Micro-RNA-128 (miRNA-128) down- regulation in glioblastoma targets ARP5 (ANGPTL6), Bmi-1 and E2F-3a, key regulators of brain cell proliferation. J.
Neurooncol. 98, 297-304.
7. Ebert, M. S. and Sharp, P. A. 2012. Roles for microRNAs in conferring robustness to biological processes. Cell 149, 515-524.
8. Filipowicz, W., Bhattacharyya, S. N. and Sonenberg, N. 2008.
Mechanisms of post-transcriptional regulation by microRNAs:
are the answers in sight? Nat. Rev. Genet. 9, 102-114.
9. Ge, Y. and Chen, J. 2011. MicroRNAs in skeletal myogenesis.
Cell Cycle 10, 441-448.
10. Greene, N. P., Brown, J. L., Rosa-Caldwell, M. E., Lee, D.
E., Blackwell, T. A. and Washington, T. A. 2018. Skeletal muscle insulin resistance as a precursor to Diabetes: Beyond glucoregulation. Curr. Diabetes Rev. 14, 113-128.
11. Han, H., Wang, L., Xu, J. and Wang, A. 2018. miR-128 in- duces pancreas cancer cell apoptosis by targeting MDM4.
Exp. Ther. Med. 15, 5017-5022.
12. Hauser, B., Zhao, Y., Pang, X., Ling, Z., Myers, E., Wang, P., Califano, J. and Gu, X. 2015. Functions of miRNA-128 on the regulation of head and neck squamous cell carcinoma growth and apoptosis. PLoS One 10, e0116321.
13. Horak, M., Novak, J. and Bienertova-Vasku, J. 2016. Muscle- specific microRNAs in skeletal muscle development. Dev.
Biol. 410, 1-13.
14. Khan, A. P., Poisson, L. M., Bhat, V. B., Fermin, D., Zhao, R., Kalyana-Sundaram, S., Michailidis, G., Nesvizhskii, A.
I., Omenn, G. S. and Chinnaiyan, A. M. 2010. Quantitative proteomic profiling of prostate cancer reveals a role for miR- 128 in prostate cancer. Mol. Cell. Proteomics 9, 298-312.
15. Kim, S. H., Yi, S. J., Lee, H., Kim, J. H., Oh, M. J., Song, E. J., Kim, K. and Jhun, B. H. 2019. β2-Adrenergic receptor (β2-AR) agonist formoterol suppresses differentiation of L6 myogenic cells by blocking PI3K-AKT pathway. Anim. Cells Syst. 23, 18-25.
16. Lian, B., Yang, D., Liu, Y., Shi, G., Li, J., Yan, X., Jin, K., Liu, X., Zhao, J. and Shang, W. 2018. miR-128 targets the SIRT1/ROS/DR5 pathway to sensitize colorectal cancer to TRAIL-induced apoptosis. Cell. Physiol. Biochem. 49, 2151- 2162.
17. Liang, R. F., Li, M., Yang, Y., Wang, X., Mao, Q. and Liu, Y. H. 2017. Circulating miR-128 as a potential diagnostic
biomarker for glioma. Clin. Neurol. Neurosurg. 160, 88-91.
18. Nigi, L., Grieco, G. E., Ventriglia, G., Brusco, N., Mancarella, F., Formichi, C., Dotta, F. and Sebastiani, G. 2018. Micro RNAs as regulators of insulin signaling: research updates and potential therapeutic perspectives in type 2 diabetes.
Int. J. Mol. Sci. 19, 3705.
19. Oh, M. J., Yi, S. J., Kim, H. S., Kim, J. H., Jeong, Y. H., van Agthoven, T. and Jhun, B. H. 2013. Functional roles of BCAR3 in the signaling pathways of insulin leading to DNA synthesis, membrane ruffling and GLUT4 translocation. Bio- chem. Biophys. Res. Commun. 441, 911-916.
20. Pan, J., Zhou, C., Zhao, X., He, J., Tian, H., Shen, W., Han, Y., Chen, J., Fang, S. and Meng, X. 2018. A two-miRNA sig- nature (miR-33a-5p and miR-128-3p) in whole blood as po- tential biomarker for early diagnosis of lung cancer. Sci. Rep.
8, 1-12.
21. Petersen, M. C. and Shulman, G. I. 2018. Mechanisms of in- sulin action and insulin resistance. Physiol. Rev. 98, 2133- 2223.
22. Regazzi, R. 2018. MicroRNAs as therapeutic targets for the treatment of diabetes mellitus and its complications. Expert Opin. Ther. Targets 22, 153-160.
23. Rupaimoole, R. and Slack, F. J. 2017. MicroRNA therapeutics:
towards a new era for the management of cancer and other diseases. Nat. Rev. Drug Discov. 16, 203.
24. Saarbach, J., Sabale, P. M. and Winssinger, N. 2019. Peptide nucleic acid (PNA) and its applications in chemical biology, diagnostics, and therapeutics. Curr. Opin. Chem. Biol. 52, 112- 124.
25. Sharma, M., Juvvuna, P. K., Kukreti, H. and McFarlane, C.
2014. Mega roles of microRNAs in regulation of skeletal muscle health and disease. Front. Physiol. 5, 239.
26. Sun, X. J., Wang, L. M., Zhang, Y., Yenush, L., Myers, M.
G. Jr., Glasheen, E., Lane, W. S., Pierce, J. H. and White, M. F. 2011. Role of IRS-2 in insulin and cytokine signalling.
Nature 377, 173-177.
27. Shaw, L. M. 2011. The insulin receptor substrate (IRS) pro- teins: at the intersection of metabolism and cancer. Cell Cycle 10, 1750-1756.
28. Wang, C., Qiao, X., Zhang, Z. and Li, C. 2020. MiR-128 pro- motes osteogenic differentiation of bone marrow mesen- chymal stem cells in rat by targeting DKK2. Biosci. Rep. 40, BSR20182121.
29. Wang, Z., Pang, L., Zhao, H., Song, L., Wang, Y., Sun, Q., Guo, C., Wang, B., Qin, X. and Pan, A. 2016. miR-128 regu- lates differentiation of hair follicle mesenchymal stem cells into smooth muscle cells by targeting SMAD2. Acta His- tochem. 118, 393-400.
30. Wu, L., Shi, B., Huang, K. and Fan, G. 2015. MicroRNA-128 suppresses cell growth and metastasis in colorectal carcino- ma by targeting IRS1. Oncol. Rep. 34, 2797-2805.
31. Xia, L., Song, M., Sun, M., Chen, W. and Yang, C. 2019.
miR-486 promotes Capan-2 pancreatic cancer cells pro- liferation by targeting PTEN. Front. Genet. 10, 541.
32. Yu, Y., Du, H., Wei, S., Feng, L., Li, J., Yao, F., Zhang, M., Hatch, G. M. and Chen, L. 2018. Adipocyte-derived exoso-
초록:쥐L6 근원세포에서 miR-128의 근육세포 분화와 인슐린신호에서의 역할
오명주
2․김소현
1․김지현
1․전병학
1*
(1
부산대학교 인제메카트로닉스공학과,
2식품안전처
)골격근의 분화 또는 근육 분화는 근육량과 신진대사 항상성을 유지하기 위해 중요하다. 근육 특이적 microRNAs (miRNAs)는 골격근 분화에 중요한 역할을 한다. 본 연구에서는 rat miRNAs 마이크로어레이를 사용하여 rat L6 근아세포의 근육 분화 과정에서의 miRNAs 발현 양상을 조사했다. 우리는 miR-128의 발현 증가를 발견했고, 동시 에 이미 알려진 근육 분화 조절 miRNAs인 miR-1, miR-133b와 mi-206의 발현 증가를 확인했다. 이 microarray 결과를 확인하기위해 우리는 Quantitative RT-PCR 기술을 사용하였고, microarray 결과와 유사하게 발현 초기 mRNAs와 발현 후 성숙 miRNAs에서 모두 miR-128의 발현 증가를 확인했다. 또한 Rat L6 근아세포로의 miR-128 발현 향상은 muscle creatine kinase (MCK), myogenin, myosin heavy chain (MHC)와 같은 근육분화 표지 유전 자 발현을 유발했고, 또한 MHC의 단백질 발현을 증가시켰다. 억제 PNAs를 사용한 miR-128의 작용 억제는 이러 한 근육 분화 표지 유전자들의 발현을 차단했다. 또한, miR-128 발현 향상은 Erk와 Akt 단백질의 인슐린 자극에 의한 인산화를 증가시켰고, 고인슐린혈증과 고혈당증으로 인해 유도된 인슐린 저항성으로 인한 Erk와 Akt의 억제 된 인산화를 회복했다. 이러한 발견은 miR-128이 근육분화와 인슐린 작용에 중요한 역할을 할 수 있다는 것을 시사한다.
mal miR-27a induces insulin resistance in skeletal muscle through repression of PPARγ. Theranostics 8, 2171.
33. Zhang, W., Kim, P. J., Chen, Z., Lokman, H., Qiu, L., Zhang, K., Rozen, S. G., Tan, E. K., Je, H. S. and Zeng, L. 2016.
MiRNA-128 regulates the proliferation and neurogenesis of neural precursors by targeting PCM1 in the developing
cortex. Elife 5, e11324.
34. Zhou, T., Meng, X., Che, H., Shen, N., Xiao, D., Song, X., Liang, M., Fu, X., Ju, J. and Li, Y. 2016. Regulation of insulin resistance by multiple MiRNAs via targeting the GLUT4 sig- nalling pathway. Cell. Physiol. Biochem. 38, 2063-2078.