Performance Assessment of Three Turfgrass Species, in Three Different Soil Types, and their Responses to Water Deficit in Reinforced Cells, Growing in the Urban Environment
10
0
0
전체 글
(2)
(3)
(4)
(5)
(6)
(7)
(8)
(9)
(10)
수치
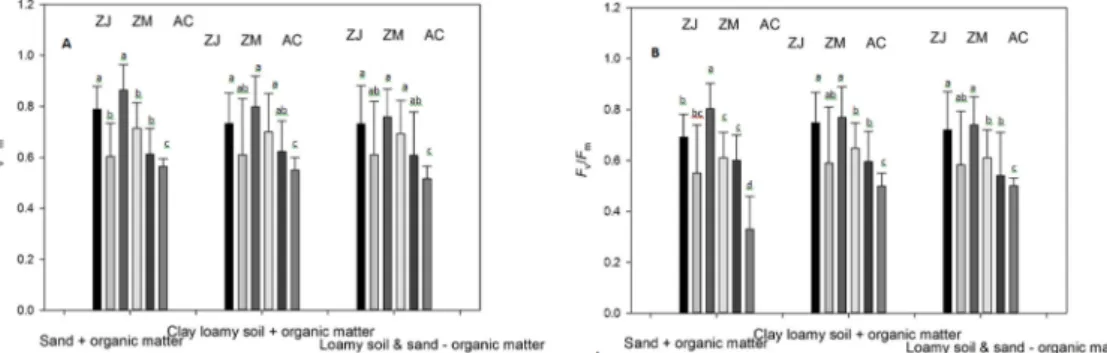
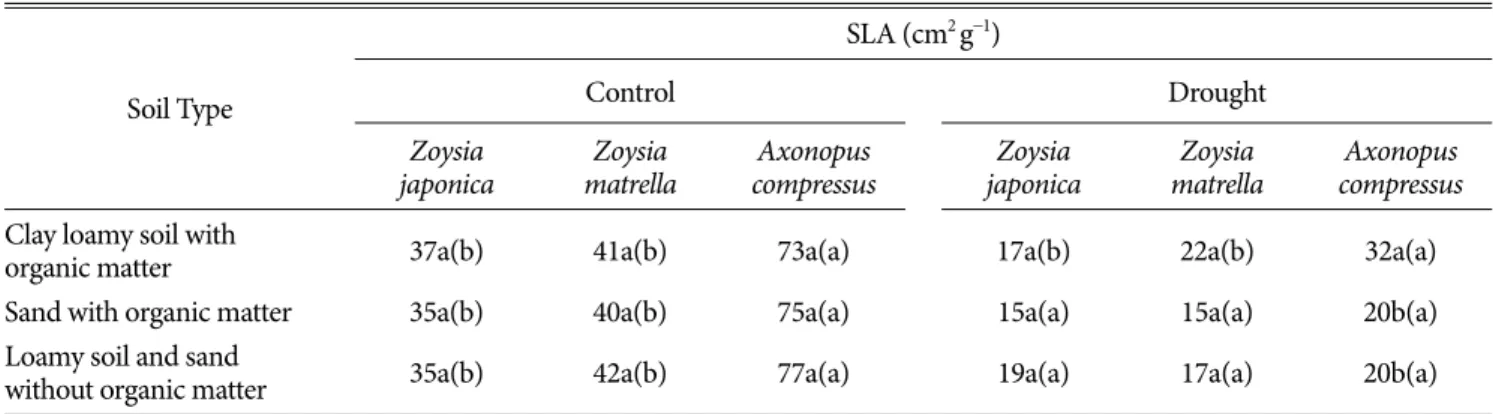

관련 문서