Received: August 3, 2018 Revised: December 5, 2018 Accepted:January 5, 2019
OPEN ACCESS
HORTICULTURAL SCIENCE and TECHNOLOGY 37(2):279-289, 2019
URL: http://www.kjhst.org pISSN : 1226-8763 eISSN : 2465-8588
This is an Open Access article distributed under the terms of the Creative Commons Attribution Non-Commercial License which permits unrestricted non-commercial use, distribution, and reproduction in any medium, provided the original work is properly cited.
Copyrightⓒ2019 Korean Society for Horticultural Science.
This work was supported by the Basic Research Program through the National Research Foundation of Korea (NRF) funded by the Ministry of Education, Science and Technology (2012- 0004486) and supported by the National Research Foundation of Korea (NRF) and the Center for Women In Science, Engineering and Technology (WISET) Grant funded by the Ministry of Science and ICT under the Program for Returners into R&D.
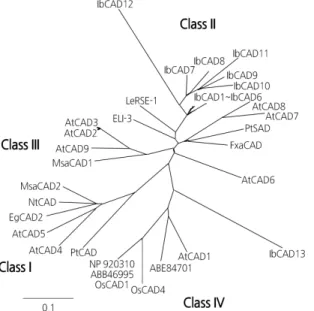
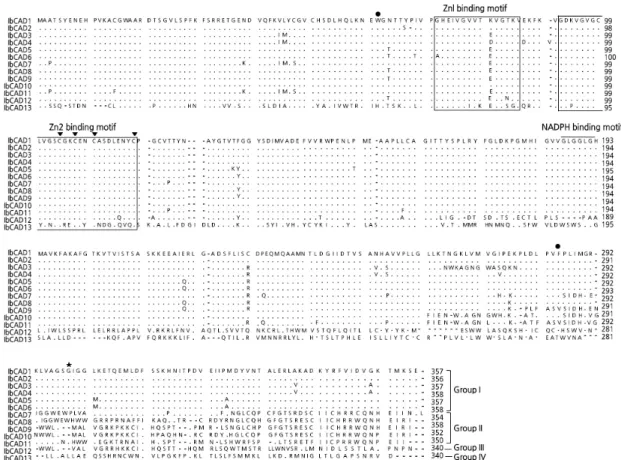
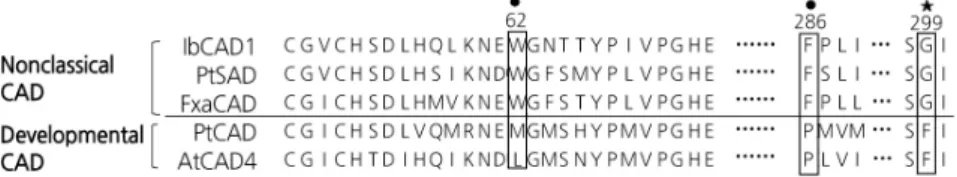
Molecular Characterization and Expression of the Cinnamyl Alcohol Dehydrogenase Gene Family in Sweet Potato (Ipomoea batatas) under
Environmental Stresses
Young-Hwa Kim 1,2 and Gyung-Hye Huh 1,2,3*
1
Ubiquitous healthcare research center, Inje University
2
Institute of Digital Anti-Aging Healthcare, Graduate School of Inje University
3