80 책임저자:황혜진, 614-714, 부산시 부산진구 엄광로 176
동의대학교 생활과학대학 식품영양학과 Tel: 051-890-1594, Fax: 051-890-2646 E-mail: [email protected]
접수일:2013년 2월 8일, 1차 수정일:2013년 2월 13일, 2차 수정일:2013년 2월 15일, 게재승인일:2013년 2월 19일
Correspondence to:Hye-Jin Hwang
Department of Food and Nutrition, College of Human Ecology, Dong-Eui University, 176, Eomgwang-ro, Busanjin-gu, Busan 614-714, Korea
Tel: +82-51-890-1594, Fax: +82-51-890-2646 E-mail: [email protected]
잎파래 메탄올 추출물의 항비만 활성
1부산대학교 식품영양학과, 동의대학교 2항노화연구소, 3블루바이오소재개발 및 실용화 지원센터,
4생활과학대학 식품영양학과, 5자연과학대학 분자생물학과, 6한의과대학 생화학교실
최은옥1,2ㆍ김향숙2,4ㆍ한민호3ㆍ박 철5ㆍ최영현2,3,6ㆍ황혜진2,3,4
Anti-obesity Activities of Methyl Alcohol Extract of Enteromorpha linza in 3T3-L1 Preadipocytes
Eun-Ok Choi1,2, Hyang Suk Kim2,4, Min Ho Han3, Cheol Park5, Yung Hyun Choi2,3,6 and Hye-Jin Hwang2,3,4
1Department of Food and Nutrition, Pusan National University, Busan 609-735, 2Anti-Aging Research Center,
3Blue-Bio Industry RIC, 4Department of Food and Nutrition, College of Human Ecology,
5Department of Molecular Biology, College of Natural Sciences, Dong-Eui University, Busan 614-714,
6Department of Biochemistry, Dong-Eui University College of Oriental Medicine, Busan 614-052, Korea
The aim of this study was to evaluate a methyl alcohol extract of Enteromorpha linza (MEEL) for its anti-obesity effects. We demonstrated the inhibitory effects of MEEL on adipogenesis by investigating the lipolytic effects of MEEL in 3T3-L1 preadipocytes. Treatment with MEEL significantly suppressed terminal differentiation of 3T3-L1 preadipocytes in a dose-dependent manner, as confirmed by a decrease in lipid droplet content observed by Oil Red O staining. The concentrations of cellular triglycerides were also reduced in 3T3-L1 cells after treatment of MEEL as compared with the controls. Treatment with 300 and 500 μg/ml of MEEL caused 67 and 19% reduction in lipid droplet content, respectively. In addition, MEEL significantly reduced the expression levels of key pro-adipogenic transcription factors including peroxisome proliferator-activated receptor γ (PPARγ), cytidine-cytidine-adinosine-adenosine- thymidine (CCAAT)/enhancer binding proteins α (C/EBPα) and C/EBPβ as compared with controls.
Taken together, these findings demonstrated that MEEL showed strong anti-obesity effects. (Cancer Prev Res 18, 80-87, 2013)
Key Words: Enteromorpha linza, 3T3-L1, Obesity, PPARγ, C/EBP
서 론
최근 해조류는 육상식물과는 달리 물질대사의 경로가 특이하고 새로운 구조를 갖는 생리활성 물질을 생산할 수 있는 소재로 각광받고 있고, 이에 포함된 식이성 섬유 소는 항산화, 항돌연변이성, 항균 및 항암 효과와 같은 중요한 기능성을 보여주고, 인체에서의 지방대사 완화
에 중요한 역할을 하는 것으로 보고되어 있다.1) 특히 해 조류에 다량으로 함유되어 있는 점질성 다당류들은 그 특성이 독특하여 생리활성이 강한 물질로 알려져 있어 서 해조류로부터의 생리활성물질 확인 및 기능성 식품 개발에 관심이 모아지고 있다.2) 이에 많은 해조류가 전 세계 다양한 지역에서 전통 약제, 식재료, 건강 보조품으 로 사용되고 있으며,3) 해조류 내에 함유되어 있는 생리 기능성 물질 탐색의 필요성에 따라 해조류의 이용가치
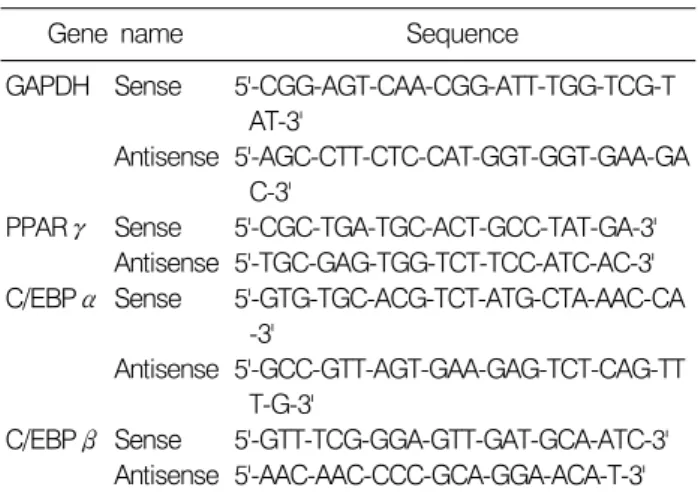
Table 1. Sequence of primers used for RT-PCR
Gene name Sequence
GAPDH Sense 5'-CGG-AGT-CAA-CGG-ATT-TGG-TCG-T AT-3'
Antisense 5'-AGC-CTT-CTC-CAT-GGT-GGT-GAA-GA C-3'
PPARγ Sense 5'-CGC-TGA-TGC-ACT-GCC-TAT-GA-3' Antisense 5'-TGC-GAG-TGG-TCT-TCC-ATC-AC-3' C/EBPα Sense 5'-GTG-TGC-ACG-TCT-ATG-CTA-AAC-CA
-3'
Antisense 5'-GCC-GTT-AGT-GAA-GAG-TCT-CAG-TT T-G-3'
C/EBPβ Sense 5'-GTT-TCG-GGA-GTT-GAT-GCA-ATC-3' Antisense 5'-AAC-AAC-CCC-GCA-GGA-ACA-T-3' 는 더욱 높아지고 있다.4)
Adipogenesis (지방세포형성)는 세포형태의 변화, 호르 몬 민감성의 변화, 유전자 발현의 변화 및 단백질 발현의 변화가 일어나는 복합적인 과정이다. 3T3-L1 세포는 지 방전구세포(preadipocyte)로서 다양한 호르몬 및 perox- isome proliferator-activated receptor γ (PPARγ), cytidine-cy- tidine-adinosine-adenosine-thymidine (CCAAT)/enhancer bind- ing proteins (C/EBPs), sterol regulatory element binding pro- teins (SREBP) 등과 같은 지방분화 유도인자에 의해 성숙 한 지방구로 분화되면서 세포내 triglyceride (TG) 축적에 관여하는 다양한 지방세포화 유전자와 효소 활성이 증 가된다.5,6) Adipogenesis 경로에서 PPARγ와 C/EBPα의 상 위에 위치하는 C/EBPβ와 C/EBPδ는 세포분열 유도물질 과 호르몬의 자극에 의해 분화초기에 빠르게 증가되어 PPARγ와 C/EBPα의 발현을 증가시키고 분화의 후기에 는 감소한다.7,8) C/EBPβ와 C/EBPδ의 발현은 insulin/dexa- methasone/3-isobutyl-1-methylxanthine (IBMX) 복합물에 의해 유도되며, 세포 내 phosphodiesterase의 활성을 억제하여 cAMP의 분해를 억제함으로써 세포 내 cAMP수준을 증가 시키며 cAMP 생성을 촉진시켜 발현을 증가시킨다.9~11) Adipogenesis의 master regulator (핵심 조절자)로 불리는 PPARγ와 C/EBPα는 서로 상호작용하여 상승효과를 가 지며 그 결과로 adiponectin (ADIPOQ), glucose transporter (GLUT) 4, fatty acid synthase (FASN) 등을 포함한 adipo- genesis의 terminal marker의 발현을 증가시킨다.12,13) C/EBP β와 C/EBPδ 이외에도 PPARγ와 C/EBPα의 상위에 위 치한 다양한 pro-adipogenic regulator (지방세포형성 유도 조절자) 및 anti-adipogenic regulator (지방세포형성 억제 조절자)들이 분화의 초기와 후기에 발현되거나 감소되 면서 adipogenesis를 조절한다.13)
본 연구에 사용된 잎파래(Enteromorpha linza (L.) J. Ag.)는 녹조식물 갈파래과에 속하는 해조류로 독특한 맛과 향 을 지녀 예로부터 널리 식용되어 왔으며, 우리나라 전 연안에 분포하며 특히 전라남도 지방에 많이 서식하고 있다.14) 편평한 엽상체로서 모양은 피침상, 신장형 또는 선상이며, 단조이고 하부에 더러 조금의 가지를 분출하 는 수가 있고 몸의 하부는 매우 가늘고 가장자리는 주름 이 있거나 또는 평탄하다. 보통 길이는 10∼20 cm이고 넓게 된 것은 막의 대부분이 유착해서 막혀있고 양 가장 자리만 유착하지 않고 남아있다. 색은 선녹색이고 질은 엷은 막질이며 11월경에서 5, 6월까지 번무한다.15,16) 잎 파래는 칼륨 성분이 풍부하고 파래의 추출물은 세포의 독성에 대한 항산화효과, 면역 증강효과, 암세포 성장억 제에 대해서 보고되어 있다.17) Choi17)의 연구에 의하면
잎파래 당단백질은 에탄올로 유도된 쥐의 위 손상을 보 호하는 효과가 있었으며, Kim과 Han18)은 잎파래 추출물 이 식품부패미생물에 대한 높은 항균성을 지니고 있다 고 보고하였다.
현재 다양한 해조류의 기능성에 대한 연구가 활발히 수행되고 있으나 다른 해조류에 비해서 잎파래의 기능 성에 대한 연구는 아직 부족한 실정이다. 따라서 본 연구 에서는 잎파래 추출물의 항비만 효능을 알아보기 위해 3T3-L1 지방전구세포에서 잎파래 메탄올 추출물에 의한 항비만 효과 및 지방세포 분화에 관여하는 PPARγ, C/EBPα 및 C/EBPβ 등과 같은 지방세포화 전사인자들 의 발현에 어떠한 변화가 유발되었는지를 조사하였다.
재료 및 방법 1. 시료
본 연구에 사용된 잎파래 메탄올 추출물(methyl alcohol extract of E. linza, MEEL)은 제주 유용생물자원추출물은행 (Jeju Bio-Resource Extract Bank, Jeju, Korea)에서 구입하여 이용하였다.
2. 실험재료
본 실험에서 mRNA 발현 양 분석을 위하여 Bioneer (Taejeon, Korea)에서 구입한 primer 염기서열은 Table 1에 나타내었으며, 단백질 분석을 위하여 사용된 PPARγ, C/EBPα, C/EBPβ 및 actin 항체는 Santa Cruz Biotechnol- ogy Inc. (Santa Cruz, CA, USA) 및 Cell Signaling Technology (Beverly, MA, USA)에서 구입하였다. Immunoblotting을 위 해 2차 항체로 사용된 peroxidase-labeled donkey anti-rabbit 및 peroxidase-labeled sheep anti-mouse immunoglobulin은
Amersham Life Science Corp. (Arlington Heights, IL, USA)에 서 구입하였다. 또한 3T3-L1 지방전구세포의 분화를 위 하여 사용된 insulin, dexamethasone 및 IBMX와 지방세포 내 TG 생성을 확인하기 위하여 사용된 Oil Red O는 Sigma-Aldrich Chemical Co. (St. Luis, MO, USA)에서 구입하 였다.
3. 세포배양
실험에 사용된 3T3-L1 지방전구세포는 90%의 Dulbecco’s Modified EaDCRT Media (DMEM, Gibco-BRL, Grand Island, NY, USA), 10%의 bovine calf serum (BCS, Gibco-BRL) 및 1%
의 penicillin 및 streptomycin (Gibco-BRL)이 포함된 배지를 사용하여 37oC, 5% CO2 조건 하에서 배양하였다. 세포수 의 증식에 따른 과밀도 현상을 해소하기 위하여 성장배 지의 교환을 매 48시간마다 실시하여 적정수의 세포를 유지하였다.
4. MTT assay
각 물질들의 세포독성 측정과 더불어 실험에 사용될 농도 범위 결정을 위하여 MTT assay를 하였다. 이를 위하 여 세포 배양용 6 well plate에 3T3-L1 지방전구세포를 분 주하여 confluent 상태까지 배양한 후 MEEL을 100∼600 μg/ml의 농도로 처리하였다. 72시간 후 희석된 0.5 mg/ml 농도의 tetrazolium bromide salt (MTT, Amresco, Solon, OH, USA)를 200 μl씩 분주하고 2시간 동안 CO2
incubator에서 배양시킨 다음 배지와 MTT 시약을 깨끗하 게 제거하였다. DMSO를 1 ml 씩 분주하여 well에 생성된 formazan을 모두 녹인 후 96 well plate에 200 μl씩 옮겨서 ELISA reader (Molecular Devices, Sunnyvale, CA, USA)로 540 nm에서 흡광도를 측정하였다. 각 세포에 대한 독성은 각 각의 대조군의 평균 흡광도 값을 구하여 평균 흡광도 값 에 대한 백분율로 나타내었다.
5. 지방세포 분화유도
3T3-L1 지방전구세포를 세포 배양용 6 well plate에서 confluent 상태까지 배양한 후 10 μg/ml insulin, 10 μM dexamethasone 및 0.5 mM 3-isobutyl-1-methylxanthine (IBMX)이 포함된 분화배지로 교환하여 지방전구세포의 분화를 유도하였다. 2일간 분화유도 후 매 2일마다 10 μg/ml insulin이 포함된 배지로 교환하여 지방전구세포 의 분화를 촉진하였다.
6. Oil Red O staining
지방전구세포 분화 시에 형성된 지방의 함량을 확인
하기 위하여 Oil Red O 염색을 실시하였다. 준비된 3T3-L1 지방전구세포를 PBS로 세척한 후 3.7% formalin으 로 10분간 고정하고 60% isopropanol을 이용하여 세척한 다음 Oil Red O solution을 처리하여 실온에서 20분간 염 색하였다. 염색 후 Oil Red O solution을 제거하고 PBS로 4회 세척한 다음 염색된 세포를 위상차 현미경을 이용하 여 관찰하였다. 또한 Oil Red O 염색 후 정량적 분석을 위하여 100% isopropanol을 이용하여 지방을 추출한 후 96 well plate에 200 μl씩 옮겨서 ELISA reader로 500 nm에 서 흡광도를 측정하였고 대조군의 흡광도 값에 대한 백 분율로 나타내었다.
7. Reverse transcriptase polymerase chain re- action (RT-PCR)에 의한 mRNA 분석
MEEL을 처리하면서 분화시킨 3T3-L1 지방전구세포의 배지를 제거한 후 TRIzol reagent (Invitrogen Co., Carlsbad, CA, USA) 1 ml을 첨가하여 4oC에서 1시간 동안 반응시키 고 200 μl의 chloroform을 첨가하여 4oC에서 30분 동안 반응시켰다. 14,000 rpm, 4oC에서 15분간 원심분리 시켜 400 μl의 상층액을 분리하여 동량의 isopropanol을 첨가 하여 4oC에서 30분 동안 반응시킨 후 14,000 rpm, 4oC에 서 30분간 원심분리시켜 total RNA를 분리하였다. 분리 된 RNA를 DEPC water를 이용하여 녹이고 정량한 후, Oligo dT primer와 AMV reverse transcriptase를 이용하여 RNA에서 cDNA를 합성하였다. 합성된 cDNA를 polymer- ase chain reaction (PCR)의 방법으로 증폭하였다. 각 PCR 산물들의 양적 차이를 확인하기 위하여 1X TAE buffer로 1.5% agarose gel을 만들고 well 당 각각의 primer에 해당하 는 PCR 산물에 DNA gel loading solution을 섞어서 loading 한 후 100 V에서 전기영동을 하였다. 전기영동으로 DNA 분리가 끝난 gel을 ethidium bromide (EtBr)을 이용하여 염 색한 후 UV 하에서 확인하였으며, housekeeping 유전자인 glyceraldehyde-3-phosphate dehydrogenase (GAPDH)를 in- ternal control로 사용하였다.
8. Western blot analysis에 의한 단백질 발현 분석
MEEL을 처리하면서 분화시킨 3T3-L1 지방전구세포에 적당량의 lysis buffer [25 mM Tris-Cl (pH 7.5), 250 mM NaCl, 5 mM EDTA, 1% NP-40, 1 mM phenymethylsulfonyl fluoride (PMSF), 5 mM dithiothreitol (DTT)]를 첨가하여 4oC 에서 1시간 동안 반응시킨 후, 14,000 rpm으로 30분간 원 심분리하여 상층액에 있는 총 단백질을 분리하였다. 상 층액의 단백질 농도는 Bio-Rad 단백질 정량 시약(Bio-Rad, Hercules, CA, USA)과 그 사용방법에 따라 정량한 다음
Fig. 1. Effect of MEEL on the cell viability in 3T3-L1 mouse preadipocytes. 3T3-L1 cells were cultured with various concentrations of MEEL for 72 h. Data were expressed as percentage of control. The cell viability was measured by the metabolic-dye-based MTT assay. Each point represents the mean±SD of three independent experiments.
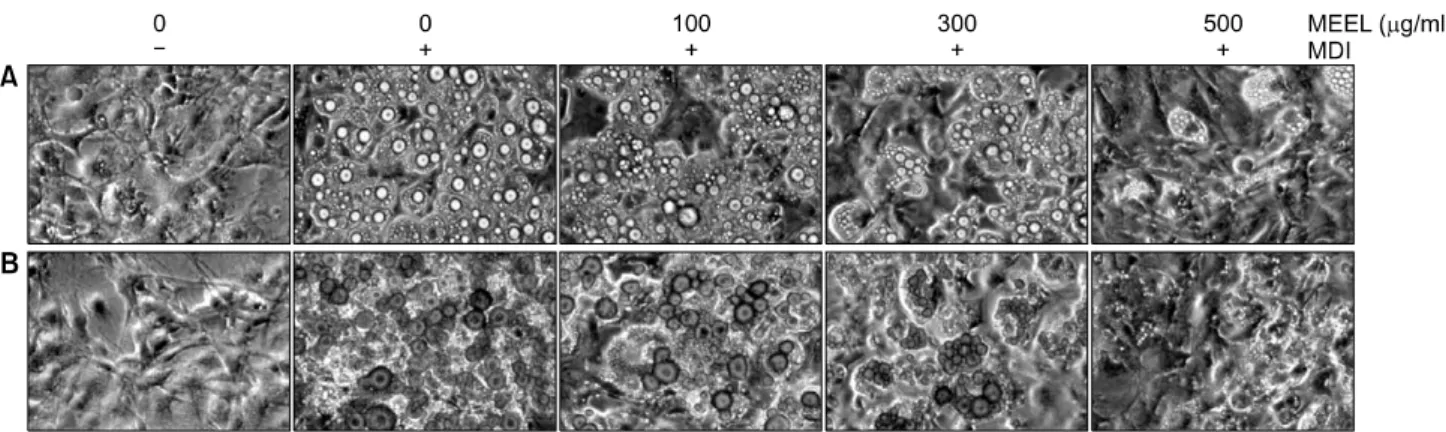
Fig. 2. Effects of MEEL on the morphological changes and lipid droplet accumulation of differentiated 3T3-L1 mouse preadipocytes.
(A) Differentiation of confluent 3T3-L1 mouse preadipocytes was initiated with MDI (0.5 mM 3-isobutyl-1-methylxanthine, 10 μM dexamethasone, and 10 μg/ml insulin) and maintained DMEM-5% FBS medium in the absence or presence of MEEL. After day 8, differentiating 3T3-L1 cells were visualized by light microscopy (magnification, ×200). (B) After day 8, cells were fixed and stained with Oil Red O to visualize lipid droplets by light microscopy (magnification, ×200).
동량의 Laemmli sample buffer (Bio-Rad)를 섞어서 sample을 만들었다. 동량의 sample을 sodium dodecyl sulfate (SDS) -polyacrylamide gel을 이용하여 전기영동으로 분리한 후, PVDF membrane (Bio-Rad)으로 electroblotting에 의해 전이 시켰다. 분리된 단백질이 전이된 PVDF membrane을 5%
skim milk를 처리하여 비특이적인 단백질들에 대한 blocking을 실시하고 1차 항체를 처리하여 상온에서 2시 간 이상 또는 4oC에서 over-night 시킨 다음 PBS-T로 세척 (10분간 4번)하고 처리된 1차 항체에 맞는 2차 항체 (PBS-T로 1:1,000으로 희석하여 사용)를 사용하여 상온 에서 1시간 정도 반응시켰다. 반응이 끝난 후 암실에서
Enhanced Chemiluminoesence (ECL) solution (Amersham Life Science Corp.)을 적용시킨 다음 X-ray film에 감광시켜 특 정 단백질의 발현양을 분석하였다.
9. Statistical analysis
모든 실험 결과는 SPSS ver. 18.0 (SPSS Inc., Chicago, IL, USA) 통계 프로그램을 이용하여 평균(mean)±표준편차 (SD)로 나타냈다. 각 실험군의 분석 항목별 통계의 유의 성은 Student t-test와 Duncan's multiple range test를 이용하 여 p<0.05 수준에서 검증하였다.
결 과
1. MEEL이 3T3-L1 세포 생존율에 미치는 영향
3T3-L1 지방세포에서 MEEL의 세포독성 여부를 조사 하기 위해서 100∼600 μg/ml의 농도에서 MTT assay 실 험을 수행하였다. MEEL을 처리하였을 때 대조군에 비해 세포 증식의 변화 정도를 퍼센트로 계산하여 MEEL이 세 포독성을 나타내는지 조사하였다. MEEL의 농도에 따른 3T3-L1 세포의 성장에 미치는 영향을 MTT assay로 분석 한 결과는 Fig. 1에 나타낸 바와 같이, 100∼500 μg/ml의 농도에서는 80% 이상의 비슷한 수준의 세포생존율을 보 였다. 따라서 생존율 및 증식에 큰 영향을 미치지 않는 500 μg/ml 농도까지를 실험 조건으로 설정하였다.
2. MEEL이 3T3-L1 세포의 분화억제에 미치는 영향 MEEL의 처리가 3T3-L1 지방전구세포의 지방세포 분 화 및 TG 축적에 미치는 영향을 확인하기 위하여 MEEL
Fig. 3. Inhibitory effect of MEEL on triglyceride accumulation of differentiated 3T3-L1 mouse preadipocytes. Triglyceride contents were determined by Oil Red O staining after treatment of the absence or presence of MEEL. The rates of triglyceride contents were measured at λ=500 nm wavelength by the ELISA reader. Data are expressed as the means±SD of three separate experiments.
Fig. 5. Effect of MEEL on the transcriptional levels of adipogenic transcription factors protein and mRNA expression in differentiated 3T3-L1 mouse preadipocytes. Differentiation of confluent 3T3-L1 mouse preadipocytes was incubated with the absence or presence of MEEL for 8 day after initiated with MDI. Total RNAs were isolated and reverse-transcribed. The resulting cDNAs were subjected to PCR with indicated primers and the reaction products were subjected to electrophoresis in 1.5% agarose gel and visualized by EtBr staining. GAPDH was used as an internal control.
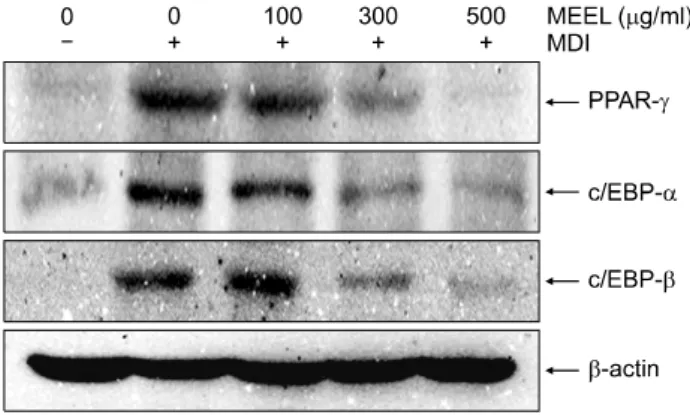
Fig. 4. Effect of MEEL on the translational levels of adipogenic transcription factors protein and mRNA expression in differentiated 3T3-L1 mouse preadipocytes. Differentiation of confluent 3T3-L1 mouse preadipocytes was incubated with the absence or presence of MEEL for 8 day after initiated with MDI. The cells were lysed and cellular proteins were separated by SDS-polyacrylamide gels and transferred onto PVDF membranes. The membranes were probed with the indicated antibodies. Proteins were visualized using an ECL detection system. Actin was used as an internal control.
을 적정농도로 처리하여 지방전구세포에서 성숙지방세 포로 분화를 유도하였다. 분화를 유도한 후 Oil Red O 염색 전후로 구분하여 지방구 생성정도를 위상차 현미 경으로 관찰하였다. Fig. 2에서 볼 수 있듯이 분화 유도를 하지 않은 세포에서와는 달리 MEEL을 처리하지 않고 분 화를 유도하였을 경우에 세포질 내 지방구의 형성이 활
발하게 유발되는 것으로 관찰되었기에, MEEL에 의한 지 방구의 형성의 억제 여부를 조사하였다. Fig. 3에 나타낸 결과에서 알 수 있듯이 중성지방의 양은 MEEL을 처리하 지 않은 대조군에 비하여 MEEL을 100 μg/ml 처리했을 경우 85%, 300 μg/ml 및 500 μg/ml 처리군에서는 각각 67% 및 19%로 나타나, MEEL이 3T3-L1 지방전구세포에 서 지방세포로의 분화억제에 효과가 높았음을 알 수 있 었다.
3. MEEL이 지방 합성 관련 효소의 mRNA 및 단백질 발현량에 미치는 영향
지방전구세포에 IBMX 복합물을 처리하면 분화초기 의 전사인자인 C/EBPβ를 활성화시키고 단계적으로 PPARγ 및 C/EBPα가 활성화되어 지방세포로의 분화가 이루어지게 된다. 이러한 과정에서 MEEL이 어떠한 영향 을 미치는지를 mRNA 및 단백질 수준에서 확인하였다.
Fig. 4와 5에 나타난 바와 같이 MDI를 처리하여 분화를 유도하였을 경우, PPARγ, C/EBPα 및 C/EBPβ의 발현 이 현저하게 증가하는 것으로 나타났으며, 이러한 분화 유도 과정에서 MEEL을 처리하였을 경우 PPARγ, C/EBP α 및 C/EBPβ의 발현이 mRNA 및 단백질 수준에서 농 도 의존적으로 감소하였음을 확인하였다. 그 중 특히 500 μg/ml의 농도에서 PPARγ, C/EBPα 및 C/EBPβ의 발현이 현저하게 감소하였음을 알 수 있었다. 이상의 결 과를 살펴볼 때 MEEL은 3T3-L1 지방전구세포에서 전사
초기와 후기를 유도하는 지방세포화 전사인자인 PPAR γ, C/EBPα 및 C/EBPβ의 발현을 억제함으로서 전사 초 기와 후기의 지방구 생성을 감소시켜 지방세포로의 분 화를 억제시킬 수 있을 것으로 추정된다.
고 찰
본 연구에서는 3T3-L1 지방전구세포에서 MEEL의 처 리가 지방세포의 성장과 지방축적 억제에 미치는 영향 을 관찰하였고, 지방 형성 관련 단백질 발현에 대한 영향 을 확인하기 위해 PPARγ, C/EBPα 및 C/EBPβ의 발현 변화를 조사하였다. 3T3-L1 지방세포의 성장에 MEEL이 어떠한 영향을 미치는지 조사하기 위해서 MTT assay를 실시한 결과, 500 μg/ml 농도까지는 80% 이상의 생존율 을 보였으나 600 μg/ml의 농도에서는 지방세포의 성장 이 급격하게 저하되어 500 μg/ml의 농도까지를 실험조 건으로 설정하였다(Fig. 1). 본 연구에 사용된 농도는 Kim 등19)의 연구에서 미역에서의 50 μg/ml 농도의 MTT 결 과와 Choi 등20)의 연구에서 톳에서의 200 μg/ml 농도의 MTT 결과와 비교했을 때 다소 높은 농도로 설정되어 잎 파래의 독성이 그리 높지 않은 것으로 추정된다.
지방세포 내의 지방구(lipid droplet)는 지질의 대사 및 조절에 중요한 역할을 하는 것으로 알려져 있으며, 지방 구 내에 주로 존재하고 있는 TG의 분해와 이로 인한 glycerol의 유출은 세포 내 지방의 축적을 조절하는 중요 한 기전으로 여겨져 왔다.21) 따라서 3T3-L1 지방전구세 포가 지방세포로의 분화과정에 나타나는 지방구 생성에 MEEL이 어떠한 영향을 미치는지를 확인하였다. 지방전 구세포를 지방세포로 분화 시키게 되면 세포 내에 지방 이 축적되게 되는데 대부분이 중성지방이기 때문에 분 화된 3T3-L1세포는 중성지방이 축적된다. 본 실험에 사 용한 Oil Red O 용액은 중성지방과 cholesterol ester만이 염색되기 때문에 이 실험을 통해 MEEL이 지방세포로의 분화와 지방축적을 억제하는지에 대해 알 수 있다. 실험 결과 분화유도를 하지 않은 세포군과 대조적으로 MEEL 을 처리하지 않고 분화를 유도하였을 경우 세포질 내 지 방구의 형성이 활발하게 유발되는 것으로 관찰되었다.
그에 비해 MEEL을 처리했을 경우 지방구의 형성이 처리 농도에 의존적으로 억제되는 것이 확인되었으며, 세포 내 축적된 TG량도 MEEL 처리 시 농도 의존적으로 억제 되었음을 확인하였다(Fig. 2, 3). Kim 등19)의 연구에서 미 역 에탄올 추출물이 TG 축적에 미치는 영향을 살펴본 결과 50 μg/ml 농도에서 지방축적이 40% 정도로 감소 하는 것으로 나타났고, Choi 등20)의 연구에서 톳의 열수
분획물은 200 μg/ml 농도에서 지방축적을 50% 이상 억 제한다고 보고한 바 있다. 따라서 본 연구에서 사용된 MEEL의 농도가 다소 높게 설정되었으나 세포성장에 영 향을 크게 미치지 않는 범위 내에서는 다른 해조류에 비 해 지방형성 억제효과가 뛰어난 것으로 추정된다.
지방전구세포에 IBMX 복합물을 처리하게 되면 지방 세포로 분화하게 되는데 분화과정에 있어서 많은 전사 인자들이 필요하며 분화됨에 따라 지방세포에 특이적인 유전자들이 발현된다. 3T3-L1 지방전구세포가 지방세포 로 분화되는 과정은 C/EBPβ와 C/EBPδ를 중심으로 조 절되는 초기 분화와 PPARγ와 C/EBPα를 중심으로 조 절되는 후기 분화로 나누어진다. Mitogen (세포분열 유도 물질)과 호르몬의 자극으로 초기 분화가 시작되면 다양 한 인자들에 의해 C/EBPβ와 C/EBPδ의 발현은 상승되
고,22,23) C/EBPβ와 C/EBPδ는 서로 협동적으로 혹은 단
독으로 PPARγ와 C/EBPα의 발현을 조절한다.24~27) Adi- pogenesis의 핵심 조절자로 불리는 PPARγ와 C/EBPα는 서 로 상호작용하여 상승효과를 가지며 그 결과로 ADIPOQ, GLUT4, FASN 등을 포함한 adipogenesis의 terminal marker의 발현을 증가시킨다.12,13) 이렇게 분화된 세포는 지방구 생 성 및 세포의 크기 증가 등과 같은 형태적 특징과 더불 어 특이적인 유전자의 발현을 유발함으로서 지방세포의 특징을 지니게 된다. 따라서 지방세포화 전사인자인 PPARγ, C/EBPα 및 C/EBPβ의 발현에 어떠한 영향을 미치는지 mRNA 및 단백질 수준에서 확인하였다. MDI 를 처리하지 않았을 경우와 대조적으로 MDI를 처리하 여 분화유도를 하였을 경우 PPARγ, C/EBPα 및 C/EBP β의 발현이 현저하게 증가하는 것으로 나타났으며, 이 러한 분화유도 과정에서 MEEL을 처리하였을 경우 PPAR γ, C/EBPα 및 C/EBPβ의 발현이 단백질 및 mRNA 수준 에서 농도 의존적으로 감소하였음을 확인하였다(Fig. 4, 5). C/EBPβ는 분화초기를 조절하는 전사인자이고 PPAR γ및 C/EBPα는 분화후기를 조절하는 전사인자인 점을 고려할 때, MEEL은 3T3-L1 지방세포에서 전사초기와 후 기에 모두 영향을 미처 단백질과 mRNA 수준에서 지방 구 생성을 감소시키고 지방세포로의 분화를 억제시킬 수 있을 것으로 추정된다. 따라서 MEEL은 3T3-L1지방세 포의 분화와 성장, 지방축적을 억제하며 lipid 생성에 관 여하는 효소의 활성 및 유전자 발현 억제효과를 나타내 므로 향후 비만 기능성 소재로서의 활용이 가능할 것이 라고 사료된다.
결 론
3T3-L1 지방전구세포에서 세포성장에 큰 영향을 없는 MEEL의 처리 조건에서, MEEL의 처리 농도 의존적으로 지방구의 형성 및 TG 생성이 유의적으로 억제되었다.
또한 MEEL은 IBMX 복합물 처리에 의해 증가되는 PPARγ, C/EBPα 및 C/EBPβ의 발현을 효과적으로 차단시켰다.
본 연구에서 관찰된 3T3-L1 지방전구세포에서 MEEL의 지방세포로의 분화 억제 및 지방 축적과 지방 생성에 관 여하는 유전자 발현 억제 효과는 잎파래 추출물이 향후 항비만 기능성 소재로의 개발 및 활용성이 매우 우수할 것임을 보여주는 결과이다.
감사의 글
이 연구는 지식경제부ㆍ부산광역시 지원 지역혁신센 터사업(RIC 08-06-07) 동의대학교 블루바이오 소재 개발 및 실용화 지원센터의 지원으로 이루어졌습니다.
참 고 문 헌
1) Stroheker T, Cabaton N, Berges R, Lamothe V. Influence of dietary soy isoflavones on the accessory sexorgans of the Wistar rat. Food Chem Toxicol 41, 1175-1183, 2003.
2) Lee HR. Antioxidant and antitumor activites of ethanol extracts from Hizikia fusiformis. M.S. Thesis, Department of Biotechnology, Graduate School of Dong-A National University, Busan, Korea, 2007.
3) Yoon SJ. Measurement of HaCaT viability by seaweed Undaria finnatifida extracts. M.S. Thesis, Department of Biotechnology, Graduate School of Pukyoung National University, Busan, Korea, 2006.
4) Choi EM, Suh KS, Kim YS, Choue RW, Koo SJ. Soybean ethanol extract increase the function of osteoblastic MC3T3-E1 cells. Phytochemistry 56, 733-739, 2001.
5) Morrison RF, Farmer SR. Hormonal signaling and transcrip- tional control of adipocyte differentiation. J Nutr 130, 3116S- 2121S, 2000.
6) Ntambi JM, Kim YC. Adipocyte differentiation and gene expression. J Nutr 130, 3122S-3126S, 2000.
7) Christy RJ, Kaestner KH, Geiman DE, Lane MD. CCAAT/
enhancer binding protein gene promoter: binding of nuclear factors during differentiation of 3T3-L1 preadipocytes. Proc Natl Acad Sci USA 88, 2593-2597, 1991.
8) Hamm JK, Park BH, Farmer SR. A role for C/EBPbeta in regulating peroxisome proliferator-activated receptor gamma activity during adipogenesis in 3T3-L1 preadipocytes. J Biol
Chem 276, 18464-18471, 2001.
9) Yeh WC, Bierer BE, Mcknight SL. Rapamycin inhibits clonal expansion and adipogenic differentiation of 3T3-L1 cells. Proc Natl Acad Sci 92, 11086-11090, 1995.
10) Tontonoz P, Hu E, Graves RA, Budavari AI, Spiegelman BM.
mPPAR gamma 2: tissue-specific regulator of an adipocyte enhancer. Genes 8, 1224-1234, 1994.
11) Darlington GJ, Ross SE, Macdougald OA. The role of C/EBP gene in adipocyte differentiation. J Biol Chem 273, 30057- 30060, 1998.
12) Morrison RF, Farmer SR. Hormonal signaling and transcrip- tional control of adipocyte differentiation. J Nutr 130, 3116S- 3121S, 2000.
13) Rosen ED, Macdougald OA. Adipocyte differentiation from the inside out. Nat Rev Mol Cell Biol 7, 885-896, 2006.
14) Hong JS, Kwon YJ, Kim YH, Kim MK, Park IW, Kang KH. Fatty acid composition of Miyeok (Undaria pinnatifida) and Pare (Enteromorpha compressa). Kor J Food Nutr 20, 376- 380, 1991.
15) Kim KY, Ahn YS, Lee IK. Growth and morphology of Enteromorpha Linza (L.) J. Ag. And E. prolifera (Muller) J. Ag.
(Ulvales, Chlorophyceae). Kor J Phycol 6, 31-45, 1991.
16) Lee JK, Jung HM, Kim SY. 1,8-Dihydroxynaphthalene (DHN)-melanin biosynthesis inhibitors increase erythritol pro- duction in Torula coralline, and DHN-melanin inhibits erythrose reductase. Appl Envir Microbiol 69, 3427-3434, 2003.
17) Choi JW. Protective effect of Enteromorpha linza extracted glycoprotein of EtOH-induced gastric mucosal damage in IEC-6 cell and S.D. rat. M.S. Thesis, Department of Food Science and Nutrition, Graduate School of Pukyoung National University, Busan, Korea, 2012.
18) Kim SJ, Han YS. Effect of green laver on the extention of shelf-life of muk (starch jelly). Kor J Soc Food Sci 14, 119-123, 1998.
19) Kim HJ, Kang CH, Kim SK. Anti-adipogenic effect of Undaria pinnatifida extracts by ethanol in 3T3-L1 adipocytes.
J Life Sci 22, 1052-1056, 2012.
20) Choi EO, Kim HS, Han MH, Choi YH, Kim BW, Hwang JA, Hwang HJ. Effects of Hizikia fusiforme extracts on adipocyte differentiation and adipogenesis in 3T3-L1 preadi- pocytes. J Life Sci 22, 1399-1406, 2012.
21) Frayn KN, Karpe F, Fielding BA, Macdonald LA, Coppack SW. Integrative physiology of human adipose tissue. Int J Obes Relat Metab Disord 27, 875-888, 2003.
22) Chen Z, Torrens JI, Anand A, Spiegelman BM, Friedman JM.
Krox20 stimulates adipogenesis via C/EBPbeta-dependent and independent mechanisms. Cell Metab 1, 93-106, 2005.
23) Zhang JW, Klemm DJ, Vinson C, Lane MD. Role of CREB in transcriptional regulation of CCAAT/enhancer-binding proein beta gene during adipogenesis. J Biol Chem 279, 4471- 4478, 2004.
24) Cao Z, Umek RM, Mcknight SL. Regulated expression of three C/EBP isoforms during adipose conversion of 3T3-L1
cells. Genes Dev 5, 1538-1552, 1991.
25) Wu Z, Bucher NL, Farmer SR. Induction of peroxisome proliferator-activated receptor gamma during the conversion of 3T3 fibroblasts into adipocytes is mediated by C/EBPbeta, C/EBPdelta, and glucocorticoids. Mol Cell Biol 16, 4128-4136, 1996.
26) Wu Z, Xie Y, Bucher NL, Farmer SR. Conditional ectopic
expression of C/EBP beta in NIH-3T3 cells induces PPAR gamma and stimulates adipogenesis. Genes Dev 9, 2350-2363, 1995.
27) Yeh WC, Cao Z, Classon M, Mcknight SL. Cascade regulation of terminal adipocyte differentiation by three members of the C/EBP family of leucine zipper proteins. Genes Dev 9, 168- 181, 1995.