알긴산 분해 Methylobacterium sp. HJM27 균주의 분리 및 특성
김옥주1·이동근1,2·이성목1·이석준1·도형주1·박혜진2·김안드레2·이재화1,2·하종명1,2*
1신라대학교 대학원 생명공학과, 2신라대학교 의생명과학대학 제약공학과
Isolation and Characteristics of Alginate-Degrading Methylobacterium sp. HJM27. Kim, Ok Ju1, Dong- Geun Lee1,2, Sung-Mok Lee1, Suck June Lee1, Hyung Joo Do1, Hye Jin Park2, Andre Kim2, Jae-Hwa Lee1,2, and Jong-Myung Ha1,2*. 1Department of Bioscience and Biotechnology, Graduate School, Silla Univer- sity, 2Department of Pharmaceutical Engineering, College of Medical & Life Science, Silla University, Busan 617- 736, Korea − This study was aimed to screen bacteria of high alginate-degrading activity, to select the nitrogen source and concentration of NaCl and sodium alginate for the production of alginate-degrading enzyme, and to determine reaction conditions of enzyme. A novel alginate-degrading bacterium was isolated from abalone (Haliotis discus hannai) and named Methylobacterium sp. HJM27 by 16S rDNA sequence analysis. The opti- mum culture conditions for the production of alginate-degrading enzyme were 1.0% sodium alginate, 0.5%
peptone, 0.3% yeast extract, 1.5% NaCl, 25oC and 48 hours incubation time. The raw enzyme showed the highest activity at 25oC and pH 9, and produced 1.217 g - reducing sugar per liter in 0.8% (w/v) sodium algi- nate for 30 minutes. Methylobacterium sp. HJM27 and its alginate-degrading enzyme would be useful for the production of bioenergy and biofunctional oligosaccharides from seaweed.
Key words: Alginate, alginate-degrading enzyme, brown algae, Methylobacterium sp., marine organism, seaweed
서 론
알긴산(alginic acid)은 D-mannuronic acid와 L-guluronic acid가 α-1,4 결합 또는 β-1,4 결합으로 이루어진 hetero형 다당류로 주로 갈조류의 세포벽을 구성한다[3, 6]. 또한 알긴 산은 비만 억제효과, 장의 연동운동 촉진을 통한 변비 치유, 항콜레스테롤 억제 작용, 체내의 중금속 흡수와 제거, 유해 물질의 독성을 억제하는 효과[15] 등의 다양한 생리활성으 로 기능성 식품소재로 활용이 가능할 뿐만 아니라 상처를 보 호하는 창상피복재 및 지혈[14] 등의 생리활성효과를 가지 고 있어 의약품 소재로도 응용될 수 있다. 최근 알긴산 유래 올리고당의 항균/항암 작용, 면역 증강, 항콜레스테롤 효과, 항응고 등 다양한 생체조절 기능에 대한 보고들이 있어 알 긴산을 이용한 올리고당 제조에 관한 연구가 있었다[5, 10].
해조류에서 알긴산 등의 유효성분을 분리하기 위한 물리 화학적 방법으로 첫째 고온고압, 고압 혹은 방사선으로 연 화시킨 뒤 분리하는 방법과 둘째 산과 알칼리 처리를 이용 하는 방법[25] 등이 보고되고 있으나 해조류 유래의 탄수화 물 대부분이 산이나 알칼리에 비교적 안정하여 유효성분의 분리에 어려움이 있다. 따라서 최근에는 기능성 해조 올리 고당 생산에 해조류 다당의 화학적 분해보다는 효소적 제조
에 대한 연구가 진행되고 있다[7, 11]. 알긴산 분해효소인 alginate lyase[EC 4.2.2.3]는 알긴산의 β-1,4-glycosidic 결합 을 분해하여 4,5-unsaturated nonreducing 말단을 갖는 산물 을 만든다[27]. 알긴산 분해효소는 mannuronate가 많은 알 긴산을 분해하는 poly(M) lyase [EC 4.2.2.3 (1→4)-β-D- mannuronan lyase]와, guluronate가 많은 알긴산을 분해하는 poly(G) lyase[EC 4.2.2.11 (1→4)-α-L-guluronan lyase] 등 이 있다.
산업적 이용이 가능한 알긴산 올리고당의 생산을 위해서 는 알긴산 분해효소가 필요하며, 이들 알긴산 분해효소를 분 비하는 미생물의 탐색을 위한 연구가 진행되어 왔다[22]. 알 긴산 분해효소는 성게, 전복, 불가사리, 소라 등 해조류 섭식 동물이나[7] Streptomyces sp.[12], Vibrio sp.[9, 15], Bacillus sp.[21, 22], Enterobacter sp.[8]와 Klebsiella aerogenses, Alginomonas alginica, Flabovacterium sp. 등에서 확인되었 으며, 이외에도 해수와 해저 토양, 섭식동물들의 분변 등으 로부터 알긴산 분해능이 우수한 균주들을 찾아내고 있다 [2, 19, 20, 23, 24]. 국내에서는 해조류에서 알긴산 분해력이 뛰어난 미생물 균주를 분리하여 생육특성을 연구한 바도 있 다[8]. 또한 균주가 생산하는 균체외 알긴산 분해효소에 대 한 정제 및 특성을 보고한 적도 있다[9].
한편 최근 화석연료에 의한 환경오염과 원유 등 자원고 갈과 같은 사회 문제로 인해 재생 가능한 에너지에 대한 관 심이 고조되고 있으며, 자연계의 생물량을 이용한 바이오에 너지 생산 관련 연구들이 주목 받고 있다[13, 16]. 특히 옥
*Corresponding author
Tel: 82-51-999-5467, Fax: 82-51-999-5636 E-mail: [email protected]
수수 등 식량자원을 이용한 바이오에너지 생산으로 인한 식 량가격 상승 등의 문제점을 극복하기 위한 대안으로 해조 류가 주목을 받고 있으며[17, 18] 해조류를 이용한 바이오 에너지 생산을 위해 알긴산 분해능이 뛰어난 분해효소가 요 구되고 있다.
따라서 본 연구에서는 해조류를 이용한 바이오에너지 개 발과 식의약소재로 활용가능한 알긴산 올리고당을 효소적 으로 제조하기 위한 기초 연구로서 알긴산 분해능이 우수 한 새로운 미생물 균주를 해조류 섭식동물에서 선별하고, 선별된 균주의 성장과 알긴산 분해 특성을 알아보고자 하 였다.
재료 및 방법 알긴산 분해 세균 분리원 및 전처리
알긴산 분해 세균의 분리를 위한 시료는 2009년 5월에 부 산광역시 기장군 기장읍 대변항과 경상남도 통영에서 구입 한 해양 동물인 전복(Haliotis discus hannai), 소라(Batillus cornutus), 해삼(Stichopus japonicus), 멍게(Halocynthia roretzi), 개불(Urechis unicinctus)이였다. 이들 해양 동물의 내장을 잘게 잘라서 멸균된 희석액(NaCl 2.5 g, KH2PO4 0.1 g, FeSO4·7H2O 0.05 g, KCl 0.05 g, NH4Cl 0.1 g, 증류수 1 l, pH 7)을 이용하여 연속 희석하였다.
알긴산 분해 세균의 분리
알긴산 분해 세균의 분리를 위하여 다층평판배지를 이용 하였다. 다층평판배지의 하층배지는 NaCl 2.5%(w/v), KH2PO4 0.1%(w/v), FeSO4·7H2O 0.05%(w/v), KCl 0.05%(w/v), NH4Cl 0.1%(w/v), agar 2.0%(w/v), pH 7이었 고, 상층배지는 sodium alginate 1.0%(w/v), agar 2.0%(w/
v), pH 7로 하였다[9, 10]. 연속희석으로 1000배 희석된 시 료 0.1 mL를 다층평판배지 위에 도말한 후 25oC에서 2~3 일간 배양하였다. 형성된 세균군체(colony)들 중에서 군체 크 기가 1 mm 이상인 균주들은 백금이를 이용하여 다층평판 배지에서 순수 분리하였다.
분리 균주의 알긴산 분해능 확인
순수 분리된 각 균주를 PYS 배지(peptone 0.5%(w/v), yeast extract 0.3%(w/v), sodium alginate 1.0%(w/v), NaCl 1.5%(w/v), pH 7)에 접종하고 25oC에서 150 rpm 조건으로 24시간 진탕 배양한 후에, 알긴산 분해능의 1차적 확인은 plate assay로 하였다. Plate assay는 PS 한천배지(peptone 0.5%(w/v), sodium alginate 0.8%(w/v), agar 1.5%(w/v))에 배양한 균주 30 µL를 점적하고 25oC에서 2일간 정치 배양 한 후 균주가 성장한 배지에 10% cetylpyridinium chloride monohydrate(CPC)를 배지가 잠길 정도로 투여하였고, 10분 후 알긴산 분해효소의 활성을 확인하였다[4].
분리 균주의 알긴산 분해능 측정
알긴산 분해효소 활성이 확인된 분리 균주를 PYS 배지에 접종하여 진탕배양하면서(25oC, 150 rpm) 배양액의 원심분 리(14,000×g, 4oC, 5 min) 상층액으로 제조한 조효소액이 나 타내는 환원당 생성능을 시간별로 측정하여 분석하였다. 효 소반응 산물인 환원당 측정에는 DNS법을 이용하였다[15].
Sodium alginate(0.8%, w/v)가 포함된 기질용액 1.0 mL에 조효소액 500 µL를 혼합한 후 30분 동안 항온조에서(150 rpm, 25oC) 반응 시킨 후 반응액을 원심분리(14,000×g, 4oC, 5 min)하였다. 원심 분리된 상층액 500 µL 에 DNS용액 2 mL을 가한 후 10분간 가열하였고 540 nm에서 흡광도를 측 정하였다. Mannuronic acid를 이용하여 작성한 표준 검량선 을 이용하여 생성된 환원당을 정량하였으며 환원당 생성능 을 알긴산 분해효소 활성과 비례하는 것으로 간주하였다. 효 소 1 unit는 1분에 1 µmole의 환원당을 생산하는 효소량으 로 정의하였고, 각 실험조건에서 최대 활성을 보인 것을 100%로 간주하고 다른 것들을 백분율로 비교하는 상대 활 성으로 나타내었다.
배양조건별 균주의 성장 및 알긴산 분해효소 활성 측정 알긴산 농도, 질소원 종류, yeast extract 첨가 유무, NaCl 농도의 영향(0~3%), 배양 온도(20~40oC), 배양 시간 등에 따른 알긴산 분해효소의 활성을 조사하기 위하여 PYS 배지 를 기본으로 하여 물질을 첨가하거나 온도 등을 달리하여 균 주를 배양하면서 DNS법으로 효소활성을 측정하였다.
분리균주의 동정
분해능이 우수한 균주는 16S rDNA의 염기서열을 결정하 기 위해 (주)마크로젠에 의뢰하여 16S rDNA의 염기서열을 구하였고 NCBI의 blast search를 통하여 동정하였다.
결과 및 고찰 알긴산 분해효소 생산 균주 분리 및 동정
갈조류를 섭식하는 5종의 해양동물 내장시료를 이용하였 을 때 약 5만 개의 콜로니가 분리배지 위에서 성장하는 것 이 확인되었다. 소라에서 가장 많은 수의 콜로니 성장이 확 인되었고 크기가 1 mm 이상인 콜로니를 알긴산 분해능이 우수한 균주로 간주하고 분리하였다. 분리된 균주의 알긴산 분해능을 plate assay로 1차 확인하고 DNS법으로 2차 측정 한 결과, 27종의 콜로니가 알긴산을 분해하는 활성이 높은 것으로 판단되었다. 전복에서 분리한 균주가 알긴산 분해 효 소 생산균 중 가장 활성이 높은 균주로 파악되어 이후 그 균 주로 연구를 진행하였다.
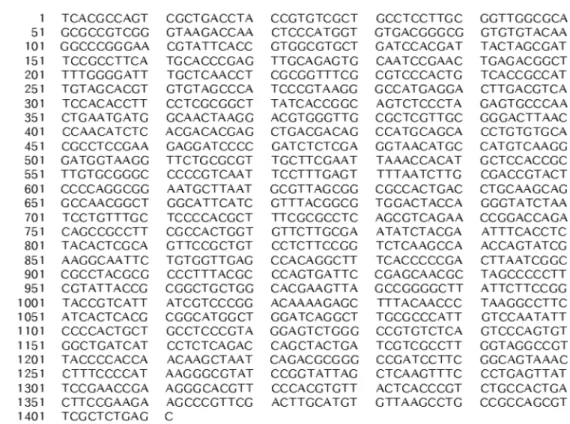
분리한 균주의 16S rDNA 염기서열 1411개를 결정하고 (Fig. 1, NCBI accession #; HM243761) blast search로 분 석한 결과 Alphaproteobacteria에 속하는 Methylobacterium
sp. 13635J(NCBI accession #; EU741082)와 99%의 유사 성을 보였다. Fig. 2는 순수분리한 Methylobacterium sp.
HJM27 균주의 16S rDNA 염기서열을 neighbor-joining
method로 나타낸 계통수(phylogenetic tree)이다. Methylo- bacterium 속 세균들과 아주 높은 연관관계를 보이는 것을 알 수 있어 분리한 HJM27 균주가 Methylobacterium 속 세 Fig. 1. 16S rDNA nucleotide sequence of Methylobacterium sp. HJM27.
Fig. 2. Phylogenetic tree generated via the neighbor-joining method showing the phylogenetic relationships of the isolated Methylo- bacterium sp. HJM27 strain based on the analysis of ca. 1150 bases of aligned 16S rDNA sequences. Bootstrap values generated from 1000 replicates are shown at each node. Values in < > are accession numbers at NCBI database and Methanobacterium uliginosum in the Domain Archaea was utilized as the outgroup. The scale bar represents 0.1 change per nucleotide.
균임을 확인할 수 있었다. 일반적으로 16S rDNA의 유사성 이 97% 이상이면 동일한 종(species)으로 분류하므로[1] 분 리한 균주를 최종적으로 Methylobacterium sp. HJM27로 명 명하였다.
질소원에 따른 균주증식과 알긴산 분해효소 활성
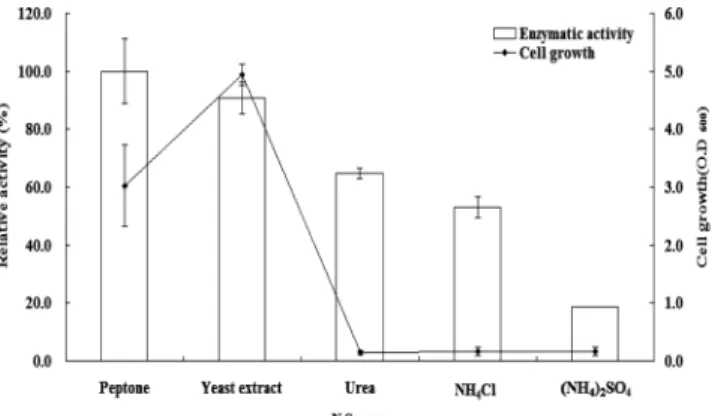
Fig. 3는 Methylobacterium sp. HJM27 균주가 서로 다른 질소원에서 나타내는 알긴산 분해효소의 활성(bar)과 세포성 장(-■-)을 나타내는 그림이다. 세포성장은 yeast extract(0.3%, w/v)에서 가장 좋았으며 효소활성은 peptone(0.5%, w/v)이 오차범위에서 가장 우수한 것으로 나타났다. 효소활성을 세 포성장으로 나눈 백분율은 NH4Cl(39.3%), urea(32.9%), (NH4)2SO4(16.8%), peptone(5.7%), yeast extract(2.1%)로 나타났지만 무기물 질소원들은 아주 낮은 효소 활성값을 보 였다. 이러한 결과는 B. lichenformis의 경우 peptone이 yeast extract에 비해 세포성장과 효소활성에서 우수하였으며 NH4Cl, urea, (NH4)2SO4등은 세포성장과 효소활성이 없었 던 보고[21]와 차이를 보였으며 Enterobacter와 Vibrio sp.의 경우 yeast extract에 비해 peptone에서 분해활성이 높았던 보고[8]와 유사성을 보인 것으로 나타났다.
Yeast extract 첨가 유무와 배양시간에 따른 세포성장과 알 긴산 분해효소 활성
Fig. 3과 같이 세포성장에는 yeast extract가 가장 우수하 였고 알긴산 분해효소 활성에는 peptone이 우수하여 peptone 이 함유된 배지에 yeast extract를 첨가하여 이들이 효소활성 등에 어떠한 영향을 미치는가에 관하여 조사하였다. Fig. 4 는 배지 성분인 peptone과 sodium alginate에 yeast extract 의 첨가 유무에 따른 세포성장과 알긴산 분해활성을 나타낸 것이다. 전반적으로 Methylobacterium sp. HJM27 균주는 peptone과 sodium alginate만 첨가된 PS 배지보다 peptone
과 sodium alginate 그리고 yeast extract가 첨가된 PSY 배 지에서 세포성장 및 알긴산 분해활성이 2배 이상 큰 것으로 나타났다. 0일째 알긴산 분해 활성이 비교적 높은 이유는 1 차 배양액을 2차 배양에 이용한 계대배양의 결과로 판단된 다. B. licheniformis AL-577 균주는 배양 144~150시간에서 균체 성장 및 조효소액의 알긴산 분해 활성이 최대였고[21], Streptomyces sp. MET 0515의 경우 배양 72시간째가 효소 활성이 최대라는 보고[12]와 비교하여 볼 때 Methylobac- terium sp. HJM27 균주는 생장과 최대효소활성이 빠른 시 간에 이루어지므로 응용 가능성이 높다고 판단할 수 있었다.
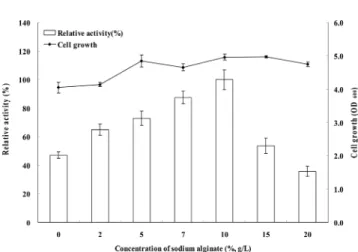
알긴산 농도에 따른 균주증식과 알긴산 분해효소 활성 Fig. 5는 Methylobacterium sp. HJM27 균주가 서로 다른 알긴산 농도에서 보이는 세포성장(bar)과 효소활성(-■-)을 나 타내는 결과이다. 전반적으로 세포성장은 알긴산 농도가 0.5%
이상이면 큰 차이를 보이지 않았다. 효소활성은 알긴산 농 도가 1.0%까지는 유의하게 증가하다가 1.5%에서 급격한 감 소하였다. 이는 알긴산 농도가 높아짐에 따라 배지의 점도 가 높아지고 일정 수준에 이르러서는 분비되는 효소의 양을 오히려 저해하는 요소로 작용할 수 있다는 것을 의미하였다 [21]. 알긴산이 전혀 없는 경우에도 활성을 보여 알긴산 분 해효소는 알긴산 유무에 상관없이 항시적으로 발현되는 것 으로 판단되었다. 알긴산의 농도가 1.0%일 때 알긴산 분해 능이 가장 높았지만, 효소활성을 세포성장으로 나눈 값은 0.7%와 1.0%가 오차범위내에서 1.0%가 우수한 것으로 나 타났다. 이러한 결과는 B. licheniformis가 2.0% 알긴산에서 효소활성과 세포성장이 최대였고[20] 1.0% 까지만 균주의 성장과 효소활성이 증가한 Yonemoto 등[26]의 보고와 차이 Fig. 3. Cell growth (bar) and the relative activity (line) of raw
enzyme at different N source. Cells were cultured in sodium alg- inate 1.0%(w/v), NaCl 1.5%(w/v) and each N source 0.5%(w/v) except yeast extract 0.3%(w/v) and cultured at 25oC and 150 rpm.
Relative activity are shown as percentages of the enzyme activity (100%, 625 U) observed at 25oC for 30 minutes.
Fig. 4. Cell growth (bar) and the relative activity (line) of raw enzyme of Methylobacterium sp. HJM27 with (■) or without (□) 0.3% yeast extract with PS medium (peptone 0.5%(w/v), sodium alginate 1.0%(w/v), NaCl 1.5%(w/v), pH 7). Culture conditions were 25oC and 150 rpm. Relative activity are shown as percentages of the enzyme activity (100%, 2090 U) observed at 25oC for 30 minutes.
를 보이는 것이었다. 이후 연구에서는 알긴산 농도를 1.0%
로 하였다.
반응온도에 따른 알긴산 분해효소 활성
Fig. 6은 조효소액이 반응 온도별로 나타내는 알긴산 분 해 활성을 나타내는 것이다. 20~40oC에서 효소활성을 비교 하니 25oC에서 가장 높았다. 한편 30oC 이상의 온도에서는 알긴산 분해 활성이 급격히 낮아지는 것을 확인하였다. 보 고된 다른 균주가 나타내는 효소활성에 대한 최적 온도와 비교하면 해양에서 분리한 Vibrio와 Enterobacter 속 세균 은 25oC와 30oC였으며[8] B. licheniformis는 30oC였고[21]
Streptomyces sp. MET 0515의 경우 최적 활성 온도가 70oC 였다[12].
알긴산 분해활성에 대한 pH의 영향
Fig. 7은 알긴산 분해활성에 영향을 미치는 pH의 결과를 나타낸 것이다. 완충용액은 200 mM sodium acetate(pH 3~6), 40 mM sodium phosphate(pH 6~8), 100 mM glycine- NaOH(pH 8~11) 완충용액을 사용하였다. 알칼리성 영역인 pH 9에서 분해 활성이 최대였다. 300 mM GTA(3,3- dimethylglutarid acid, Tris(hydroxymethyl) aminomethane, 2-amino-2-methyl-1,3-propanediol, pH 3~10) 완충용액에서 도 유사한 결과를 얻었다(data not shown). 알긴산 분해 효 소의 활성에 대한 pH의 영향은 균주별로 차이를 보였는데 Streptomyces sp. MET 0515 균주 유래 효소는 pH 7.5에서 가장 높은 알긴산 분해활성을 나타냈으며[12] B. lichenifomis AL-755 균주 유래 효소는 pH 6에서 가장 높은 활성을 나타 내어[22] 본 연구 결과와 차이를 보였다.
균주 성장 및 알긴산 분해효소 활성에 대한 NaCl 영향 NaCl 농도를 달리한 배지에서 균체의 성장 및 알긴산 분 해 활성을 살펴본 결과를 Fig. 8에 나타내었다. 분리균주는 NaCl의 농도가 0%에서 3.0%까지 성장과 알긴산 분해 활성 이 확인되었고 NaCl 2.5%에 비해 3.0%일 때 성장이 급격 히 저하되었으므로 호염성이 아닌 내염성(halotolerant) 균주 로 판단되었다.
특이한 점은 NaCl 농도가 1.0%와 2.5%에서 성장의 쌍고 점을 보였다는 점인데 반복 실험에서도 유사한 경향을 보였 다. 이는 균주가 성장하면서 floc을 형성하는 것에 의한 영 향으로 판단되었다. 또한 환원당 생성능은 세포를 제거한 조 효소액을 사용하여 세포들이 floc을 형성하여도 결과에 크게 Fig. 5. The effect of alginate concentration on cell growth (line)
and activity (bar) of raw enzyme of Methylobacterium sp.
HJM27. Cells were cultured in PSY medium at 25oC and 150 rpm. Relative activity are shown as percentages of the enzyme activity (100%, 656 U) observed at 25oC for 30 minutes.
Fig. 6. The effect of reaction temperature on the activity of extracellular alginate-degrading enzyme of Methylobacterium sp. HJM27. Cells were cultured in PSY medium (peptone 0.5%
(w/v), yeast extract 0.3%(w/v), sodium alginate 1.0%(w/v), NaCl 1.5%(w/v), pH 7) at 25oC and 150 rpm. Relative activity are shown as percentages of the enzyme activity (100%, 2090 U) observed at 25oC for 30 minutes.
Fig. 7. The effect of pH on the activity of extracellular alginate- degrading enzyme of Methylobacterium sp. HJM27. Used buffer was 200 mM sodium acetate (pH 3-6), 40 mM sodium phosphate (pH 6-8), and 100 mM glycine-NaOH (pH 8-11). Relative activity are shown as percentages of the enzyme activity (100%, 2090 U) observed at 25oC for 30 minutes.
영향을 미치지 않았겠지만 세포성장 측정에는 영향을 미쳤 을 것이다. 따라서 NaCl이 1.5%일 때 단위 배양액당 가장 높은 알긴산 분해효소 활성을 보이는 것을 알 수 있었다. 이 는 B. licheniformis의 성장과 환원당 생성의 최적 NaCl 농 도가 2.0%라는 보고[21]와 Vibrio sp. AL-145의 경우 0.5M NaCl에서 효소활성이 가장 높은 보고[9]와 비교하여 낮은 NaCl 농도였다.
Joo 등[8]은 환원당 생성능이 0.355 g/L인 균주를 고활성 균 주로 간주하였고 Uo 등[21]은 분리한 Bacillus licheniformis 의 환원당 생성능이 0.899 g/L로 상업성이 있다고 보고하였 다. 본 연구에서 분리한 HJM27 균주는 환원당 생성능이 1.217 g/L로 Joo 등[8]의 보고에 비해 343%, Uo 등[21]의 보고에 비해 35% 이상 높은 활성을 보여 상업적 활용 가능 성이 높은 것으로 판단되었다. 전분, 카라기난, 펙틴, 한천 등의 다당류에 대한 분해활성을 측정하였으나 알긴산에 대 해서만 분해활성을 보여 (data not shown) 분리한 균주는 알 긴산 분해효소를 생산하는 것으로 판단되었다[21].
향후 Methylobacterium sp. HJM27에서 알긴산 분해 효소 의 분리와 함께 yeast extract 농도 등을 이용한 효소생산 최 적화가 이루어진다면 해조류를 이용한 바이오에너지 개발 분야[18] 및 식의약 산업에서 유용한 알긴산 분해효소의 생 산과 응용에 아주 유용할 것으로 판단되었다.
요 약
본 연구는 알긴산 분해효소 활성이 뛰어난 세균을 확보하 고 선정된 균주의 효소생산과 활성에 영향을 미치는 알긴산 과 NaCl의 농도, 질소원 종류, 온도, pH 등을 파악하고자 하 였다. 해조류 섭식 동물인 전복, 소라, 해삼, 멍게, 개불 등
에서 유래한 총 5만여 콜로니 중 알긴산 분해효소 활성이 우수한 27개 균주를 분리하였고 최종적으로 전복 유래의 균 주를 선정하였다. 16S rDNA 염기서열 분석으로 선정된 균 주를 Methylobacterium sp. HJM27으로 명명하였고 알긴산 분해효소의 활성은 1.0% sodium alginate, 0.5% peptone, 0.3% yeast extract, 1.5% NaCl, 25oC, 48시간 배양에서 가 장 높았다. 알긴산 분해효소의 활성은 25oC, pH 9에서 최대 로 0.8%(w/v) sodium alginate 용액에서 30분만에 1.217 g/L 의 환원당을 생성하였다. 분리된 균주와 이 균주의 알긴산 분해효소를 이용하여 해조류를 바이오에너지와 식·의약 소 재로 활용 할 수 있을 것으로 판단된다.
감사의 글
본 연구는 2009년도 지식경제부의 재원으로 한국에너지 기술평가원(KETEP)의 지원을 받아 수행한 연구 과제입니다 (No. 20093020090020).
REFERENCES
1. Drancourt, M., P. Berger, and D. Raoult. 2004. Systematic 16S rRNA gene sequencing of atypical clinical isolates identified 27 new bacterial species associated with humans.
J. Clin. Microbiol. 42: 2197-2202.
2. Eller, J. and W. J. Payne. 1960. Studies on bacterial utiliza- tion of uronic acid. IV. Alginolytic and mannuronic acid oxidizing isolates. J. Bacteriol. 80: 193-199.
3. Fisher, F. G. and H. Dorfel. 1955. The polyuronic acids of brown algae. Part I. Z. Physiol. Chem. 302: 186-203.
4. Gacesa, P. and F. S. Wusteman. 1990. Plate assay for simultaneous detection of alginate lyase and determination of substrate specificity. Appl. Environ. Microbiol. 56: 2265- 2267.
5. Guven, K. C., Y. Ozsoy, and O. N. Ulutin. 1991. Anticoagu- lant, fibrinolytic and antiaggregant activity of carrageenans and alginic acid. Botan. Marin. 34: 429-435.
6. Haug, A., B. Larsen, and O. Smidsrød. 1966. A study of constitution of alginic acid by partial acid hydrolysis. Acta.
Chemica. Scand. 20: 183-190.
7. Jung, J. Y., S. S. Hur, and Y. H. Choi. 1999. Studies on the efficient extraction process of alginic acid in sea tangle.
Food Eng. Prog. 3: 90-97.
8. Joo, D. S., S. Y. Cho, and E. H. Lee. 1993. Isolation of alginate-degrading bacteria and production of alginate- degrading activities by bacteria. Kor. J. Appl. Microbiol.
Biotechnol. 21: 207-213.
9. Joo, D. S., J. S. Lee., J. J. Park., S. Y. Cho., C. B. Ahn, and E. H. Lee. 1995. Purification and characterization of the intracellular alginase from Vibrio sp. AL-145. Kor. J. Appl.
Microbiol. Biotechnol. 23: 432-438.
10. Joo, D. S., J. S. Lee., J. J. Park., S. Y. Cho., H. K. Kim, and Fig. 8. The effect of NaCl concentration on growth and the
activity of extracellular alginate-degrading enzyme of Methylo- bacterium sp. HJM27. Composition of basal medium and growth condition: Peptone 0.5%, sodium alginate 1%, yeast extract 0.3%, pH 7, incubation temperature 25oC, 150 rpm for 48 hrs. Relative activity are shown as percentages of the enzyme activity (100%, 2090 U) observed at 25oC for 30 minutes.
E. H. Lee. 1996. Preparation of oligosaccharides from alginic acid enzymatic hydrolysis. Korean J. Food Sci.
Technol. 28: 146-151.
11. Kim, B. J., S. D. Ha., D. J. Lim., C. Song, and J. Y. Kong.
1998. Production of agarase from marine bacterium Bacillus cereus ASK202. Korean J. Biotechnol. Bioeng. 13: 524-529.
12. Kim, H. K., J. C. Lee, N. H. Kang, S. H. Kim, J. G. Kim, and K. C. Chung. 2007. Purification and characterization of the extracellular alginate lyase from Streptomyces sp. MET 0515. J. Life Sci. 17: 625-633.
13. Kim, Y. S. 2008. Development and utilization trend of biofuel from lignocellulosic biomass. KFBS 27: 1-10.
14. Lee, B. H., S. B. Lee, and W. K. Kim. 2009. Alginate fiber.
Fiber Technol. Ind. 13: 21-24.
15. Lee, J. H. and E. Y. Lee. 2003. Isolation of alginate degrading marine bacteria and characterization of alginase.
J. Life Sci. 23: 718-722.
16. Lee, K. E., J. Y. Lee, and K. Kim. 2008. Effect of content of crop component on the bioethanol production. Korean J.
Crop Sci. 53: 339-346
17. Miller, G. L. 1959. Use of dinitrosalicylic acid reagent for determination of reducing sugar. Anal. Chem. 31: 426-428.
18. Park, J. I., H. C. Woo, and J. H. Lee. 2008. Production of bio-energy from marine algae : status and perspectives.
Korean Chem. Eng. Res. 46: 833-844.
19. Park, Y. H., D. S. Chang, and S. B. Kim. 1994. Symbiotic formation of alginate lyase in mixed culture of bacteria
isolated from soil. J. Ferment. Bioeng. 69: 192-194.
20. Sutherland, I. W. 1995. Polysaccharide lyases. FEMS Microbiol. Rev. 16: 323-347.
21. Uo, M. H., D. S. Joo, and S. Y. Cho. 2006a. Screening and cultivation characteristics of alginate degrading bacteria. J.
Korean Soc. Food Sci. Nutr. 35: 109-114.
22. Uo, M. H., D. S. Joo, S. Y. Cho, and T. S. Min. 2006b.
Purification and characterization of the extracellular alginase produced by Bacillus lichenformis AL-577. J. Korean Soc.
Food Sci. Nutr. 35: 231-237.
23. Wong, T. Y., L. A. Proston, and N. L. Schiller. 2000. Alginate lyase: review of major sources and enzyme characteristics, structure-fuction alalysis, biological roles, and application.
Annu. Rev. Microbiol. 54: 289-340.
24. Waksman, S. A. and M. C. Allen. 1934. Decomposition of polyuronic by fungi and bacteria. J. Bacteriol. 28: 213-220.
25. Yang, J. S. and S. R. Lee. 1997. Effect of ionizing radiation on the extraction yield and viscosity of alginate. Korean J.
Food Sci. Technol. 9: 194-198.
26. Yonemoto, Y., K. Mututa., A. Kimura., H. Yamaguchi, and K. Okayama. 1991. Bacterial alginate lyase; characterization of alginate lyase-producing bacteria and purification of the enzyme. J. Ferm. Bioeng. 72: 152-157.
27. http://www.chem.qmul.ac.uk/iubmb/enzyme/EC4/2/2/3.html
(Received April 22, 2010/Accepted June 7, 2010)