The Beneficial Effects of Extract of Pinus densiflora Needles on Skin Health
Jieun Choi, Woong Kim, Jaeyoung Park, and Hyeonsook Cheong*
Department of Life Science & BK21-Plus Research Team for Bioactive Control Technology, Chosun University, Gwangju 61452, Republic of Korea
Received: March 24, 2016 / Revised: April 9, 2016 / Accepted: April 11, 2016
Introduction
Natural products isolated from plants are used as important resources for medical treatment or health improvement [37]. The natural habit at of Pinus densi- flora Sieb. et Zucc. (red pine) are Korea, Japan, China, and southeastern Russia, various parts of red pine are used as supplementary health food to improve health [39]. Also, the leaves of red pine are consumed as tradi- tional medicine for liver diseases, skin diseases, and
hypertension [19, 22]. Volatile chemicals of red pine pos- sess antioxidant activity and growth inhibition activities against human intestinal bacteria [13, 29].
The skin is the primary barrier between the body and the external environment and defenses against external factors such as microbial and chemical agents [18]. Oxi- dative stress disrupts the protective function of skin and causes roughness and wrinkling [6]. The use of antioxi- dants is an effective treatment to prevent symptoms associated with the aging of skin [24].
Common skin diseases associated with inflammation such as pityriasis versicolor are caused by a fungus of the genus Malassezia [12, 25]. Among the Malassezia species, Malassezia furfur is associated with pityriasis Pinus densiflora Sieb. et Zucc. (P. densiflora) contains several phenolic compounds that exhibit biological activities, such as antimicrobial, antioxidant, and antihypertensive effects. However, the anti-inflamma- tory effect of P. densiflora on skin has rarely been reported. Malassezia furfur (M. furfur) is a commensal microbe that induces skin inflammation and is associated with several chronic disorders, such as dandruff, seborrheic dermatitis, papillomatosis, and sepsis. The aim of our study was to identify the anti-inflamma- tory effects of P. densiflora needle extracts on skin health subjected to M. furfur-induced inflammation.
The methanolic extract of the pine needles was partitioned into n-hexane, EtOAc, n-BuOH, and water lay- ers. We measured the anti-inflammatory effects (in macrophages) as well as the antioxidant, antifungal, and tyrosinase inhibitory activity of each of these layers. The antioxidant activity of the individual layers was in the order EtOAc layer > n-BuOH layer > water layer. Only the n-BuOH, EtOAc, and n-hexane layers showed antifungal activity. Additionally, all the layers possessed tyrosinase inhibition activity similar to that of ascorbic acid, which is used as a commercial control. The EtOAc layer was not cytotoxic toward the RAW 264.7 cell line. Interleukin 1 beta and tumor necrosis factor (TNF)-α expression levels in M. furfur- stimulated RAW 264.7 cells treated with the EtOAc layer were decreased markedly compared to those in cells treated with the other layers. Taken together, we believe that the needle extracts of P. densiflora have potential application as alternative anti-inflammatory agents or cosmetic material for skin health improve- ment.
Keywords: Pinus densiflora, Malassezia furfur, skin health, anti-inflammation
*Corresponding author
Tel: +82-62-230-6667, Fax: +82-62-224-6678 E-mail: [email protected]
© 2016, The Korean Society for Microbiology and Biotechnology
versicolor [15], folliculitis [1], seborrheic dermatitis [2], atopic dermatitis [38], and psoriasis [31]. At high con- centrations, this yeast can diminish the normal protec- tive function of skin and affect the body’s ability to regulate inflammation [33]. As such, the goals of treat- ment are to regulate M. furfur’s growth and associated inflammation, and to prevent the outbreak of secondary infections [9, 11].
In the inflammatory response, macrophages recognize the infection and the secreted several pro-inflammatory cytokines such as tumor necrosis factor-α (TNF-α) and interleukin(IL)-1β, IL-4, IL-6, and IL-8 induces inflam- mation response [27]. Among them, TNF-α and IL-1β are important factor to chronic inflammatory disorders [3]. Kesavan [17] investigated the influence of the Malassezia on the production of IL-1, IL-6, and TNF-α by peripheral blood mononuclear cells (PBMC). Of these, TNF-α plays a particularly important role in skin dis- eases related to inflammation [21, 36]. Inflammatory responses to TNF-α are controlled through expression of IL-1, as well as via more pro-inflammatory cytokines [34]. The natural product research has developed a vari- ety of therapeutic agents that are proven to be therapeu- tically effective against a wide range of diseases [23].
Current interest has led to the search for novel natural products with anti-inflammatory activity [40].
In this study, layers with potential anti-inflammatory action were isolated from the methanolic extract of P.
densiflora needles. Subsequently, we confirmed their various functional activities relating to skin health: anti- fungal activity against M. furfur (causative factor of seb- orrheic dermatitis), antioxidant activity directed against aging [24], and tyrosinase inhibition activity associated with the whitening of skin color [35]. Cytotoxicity and anti-inflammatory activity were confirmed from the measurement of IL-1β and TNF-α expression levels in M. furfur-activated macrophages.
Materials and Methods
Preparation of needle extracts from Pinusdensiflora Fresh needles of red pine were picked in Gokseong province, Jeollanam-Do, South Korea. Harvested nee- dles of red pine were washed clean with tap water (cleaned with 5% charcoal) and dehydrated using a spin- drier. The dehydrated pine needles (100 kg) were dried
in the shade. The dried pine needles (49.97 kg) were treated with 500 L of 80% methanol (MeOH) at 69oC for 3 h. The resultantmethanolic extracts were concentrated to 20 L. This crude extract was partitioned successively to yield layers of n-hexane (3 × 10 L), EtOAc (3 × 10 L), n-BuOH (3 × 10 L) and water [28].
2,2-Diphenyl-1-picrylhydrazyl (DPPH) radical scavenging assay
The DPPH assay was performed according to a method described by Nenadis and Tsimidou [26] with some modifications. Briefly, a methanolic DPPH solu- tion (0.04 mM; 180 μl) was added to 20 µl of different concentrations of extracts and fractions. The mixture was shaken vigorously and a decrease in absorbance was measured at 517 nm after 30 min of incubation in the dark. The blank solution contained the same amount of DPPH reagent and 20 µl of methanol and each test was performed in triplicate. The percentage of DPPH inhibition was calculated as follows:
DPPH = (1 − As/Ac) × 100
Where Ac and As are the absorbance of the control and test samples, respectively. Ascorbic acid was used as the reference.
M. furfur and Raw 264.7 culture conditions
M. furfur strain KCTC7546 was used in all experi- ments. It was cultured either on modified YPD plates or liquid medium supplemented with Tween 80 (1% yeast extract, 1% peptone, 2% glucose, 1% Tween 80) for 1 day at 30ºC under aerobic conditions. The conditions were based on published information [7] regarding nutrients and trace components that promote the growth of M.
furfur.
Mouse lymphoid microvascular epithelium immortal- ized cell line, Raw 264.7, was obtained from the ATCC (TIB-71). Raw 264.7 was grown in Dulbecco’s Modified Eagle Medium (DMEM) containing 10% fetal bovine serum (FBS), 2 mM glutamine, and 100 μg/ml penicillin- streptomycin at 5% CO2 and 37oC humidified atmo- sphere.
Antifungal activity against M. furfur
Once grown, M. furfur in liquid medium was har- vested from a 1 ml culture and suspended in 100 µl PBS.
The suspended solutions were plated onto modified YPD agar medium using a sterile spreader. Plant extracts were dissolved in dimethyl sulfoxide (DMSO) to a con- centration of 100 mg/ml. Needle extract of P. densiflora (50 µl) and control respectively were impregnated into the sterile paper discs (8 mm diameter) and incubated at 30oC for 1 day. M. furfur was visualized and images of the plates were captured. The presence of a clear zone around the discs indicated the sensitivity of M. furfur to the P. densiflora samples. The total diameter of each clear zone was measured.
Tyrosinase inhibition analysis
The activity of tyrosinase was determined by following the L-DOPA assay protocol [15]. The analysis used 1 L of the reaction mixture, which was composed of 1 mM sodium phosphate buffer (pH 6.8, 1 M Na2HPO4, 1 M NaH2PO4), 10 mM of L-DOPA and 200 unit/ml tyrosi- nase. Ascorbic acid was used as the positive control.
Cytotoxicity analysis
3-[4,5-Dimethylthiazol-2-yl]-2,5 diphenyltetrazolium bromide (MTT) assay was performed using the method previously described by Boncler et al. [4]. Raw 264.7 cells were seeded at 1 × 105 cells per well in a 96-well plate and these cells were grown in a humidified atmosphere of 5% CO2 at 37oC. The cells were then exposed to vary- ing concentrations of the pine needle extract (0 to 200μg/ml) for 24 h. After that, 100 µl of fresh medium was added to the cells. MTT solution (5 mg/ml of PBS) was added and the plate was placed in an optimal atmo- sphere at 37oC to allow the metabolically active cells to reduce MTT to blue formazan crystals. After 4 h, MTT- formazan crystals were dissolved in 50% ethanol and 50% DMSO and the absorbance was measured at 570 nm on a multifunctional plate reader (Eon, Bio-tek, USA). Comparisons were made with untreated cells.
Determination of IL-1β and TNF-α mRNA expression Raw 264.7 cells (1 × 105 cells/well) were seeded in a 6- well culture plate containing DMEM. The cells were pre- treated with various concentrations of pine needle extracts (0 to 100 μg/ml) for 2 h and then incubated in the absence or presence of M. furfur for 6 h. After incu- bation, they were collected by centrifugation and total RNA was isolated from pine needle extract-treated cells
using Hybrid-R (GeneAll, Korea) according to the manu- facturer's protocol. To synthesize cDNA, 2 μg of total RNA was primed with oligo dT and made to react with Hyperscript mix (GeneAll, Korea). To measure the mRNA level of inflammatory cytokines including IL-1β and TNF-α, we designed primers for the target genes (Table 2). cDNA was amplified and the PCR products were visualized using fluorescent dye on a UV transillu- minator. mRNA expression of target genes was analyzed by real-time PCR using SYBR Green (Takara, Japan).
Results
Antioxidant activity
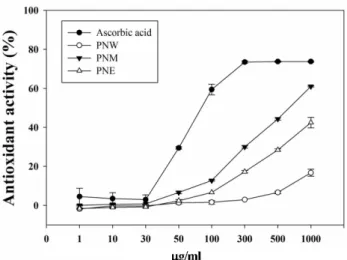
The methanolic extract of P. densiflora needles showed stronger antioxidant activity than the water or ethanol extracts. The results of dose-dependent DPPH scavenging activity are presented in Fig. 1. After parti- tioning the pine needle methanolic extract into n-hex- ane, EtOAC, n-BuOH, and H2O layers, the antioxidant activity of the individual layers was measured and was found to be in the order EtOAc layer > n-BuOH layer >
water layer (Fig. 2).
Comparison of M. furfur growth in various culture media For M. furfur culture, variable compositions of liquid and solid medium were tested to achieve optimum condi- tions. The growth of M. furfur in liquid culture was mon-
Fig. 1. DPPH (1,1-diphenyl-2-picrylhydrazyl) radical scav- enging activity of P. densiflora extracts. PNW; Pine needle water extraction, PNM; Pine needle methanol extraction, PNE;
Pine needle ethanol extraction. Ascorbic acid was used as an antioxidant and an experimental control.
itored by measuring OD600. After about 24 h, the organism was incubated on agar plates for another 24 h.
The media compositions have been described in Table 1.
Medium B and E did not have yeast extract or glucose.
M. furfur did not grow well in these media. In liquid Medium A and C containing olive oil, M. furfur grew well, but those cultures were extremely oily to spread over the agar plates. Among the five media formulations, Medium D, composed of yeast extract, peptone, and glu- cose but without olive oil, provided the best conditions for M. furfur growth in both agar and liquid culture.
Antifungal activity of pine needle extracts against M.
furfur
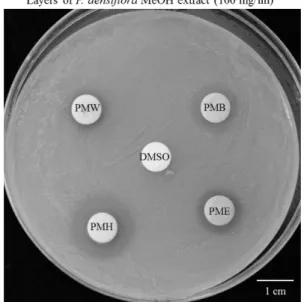
To determine fungicidal activities of pine needle
extracts, the extracts were applied at 100 mg/ml onto paper discs, which were then placed on M. furfur culture plates. Fig. 3 shows the inhibitory effect of pine needle extract on M. furfur growth. The presence of a clear zone around the disc (8 mm diameter) indicates growth inhi- bition. The total diameter of the zone of inhibition was measured for each disc and the results are given in Table 3. DMSO was used as an experimental control and to dissolve pine needle extract. The antifungal activity of its individual layers were observed to increase in the order n-hexane layer > EtOAc layer > n-BuOH layer >
Fig. 2. DPPH radical scavenging activity of various layers from P. densiflora needle methanolic extract. PME; Ethyl ace- tate layer of Pine needle methanolic extract, PMB; n-utyl alco- hol layer of Pine needle methanolic extract, PMH; n-hexane layer of pine needle methanolic extract, PNW; Water layer of pine needle methanolic extract.
Table 1. Compositions of Malassezia furfur media.
Composition OD600a
A 0.3% Yeast extract, 0.3% Malt extract, 0.5% Peptone, 1% D-glucose, 1% Olive oil, 1% Tween 80, 0.5% Glycerol, 0.1%
Monosterin
1.759
B 0.3% Sabouraud dextrose, 0.05% Glycerol monosterate, 0.4% Olive oil, 0.05% Tween 20, 0.1% Glycerol 0.451 C 0.01% Yeast extract, 1% Peptone, 0.5% Glucose, 0.05% Glycerol monoesterate, 0.4% Olive oil, 0.05% Tween 20,
0.1% Glycerol
1.121
Db 1% Yeast extract, 1% Peptone, 2% Glucose, 1% Tween 80 1.786
E 0.3% Malt extract, 0.5% Peptone, 0.1% Glycerol monoesterate, 5% Tween 20 0.501
aM. furfur was cultured in liquid medium for 24 h at 30°C and the growth of liquid cultured M. furfur were monitored by OD600.
bD [5] possessed the optimal conditions for liquid and solid medium to culture M. furfur.
Fig. 3. Antifungal activity of Pinus densiflora against M. furfur.
Antifungal activity of P. densiflora needle methanolic extract layers. PME; Ethyl acetate layer of pine needle methanolic extract, PMB, n-butyl alcohol layer of pine needle methanolic extract, PMH; n-hexane layer of pine needle methanolic extract, PNW; Water layer of pine needle methanolic extract. DMSO was used as solvent of P. densiflora.
water layer (Fig. 3, Table 3).
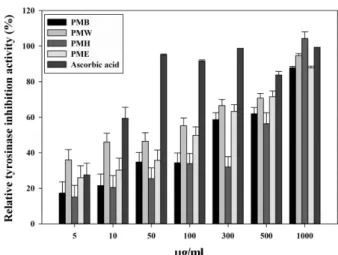
Tyrosinase inhibition activity of pine needle extract The tyrosinase inhibition activity of the pine needle extract is shown in Fig. 4. All the partitioned layers of the methanolic extract revealed significant tyrosinase inhibitory activity, comparable to that exhibited by ascorbic acid. The layers revealed dose-dependent activ- ity, and the water layer showed the strongest tyrosinase inhibition (Fig. 4).
Cytotoxicity of pine needle extract in Raw 264.7 cell line The cytotoxic effects of pine needle extract at molecu- lar and cellular levels were investigated in Raw 264.7 cultured cells via the MTT assay. The results indicated that at concentrations of 25 to 100 μg/ml, the n-hexane layer of the methanolic extract of pine needles displayed cytotoxicity while the EtOAc, n-BuOH, and water layers were not cytotoxic (Fig. 5).
Anti-inflammatory activity of pine needle extract
IL-1β and TNF-α are pro-inflammatory cytokines that are released from macrophages upon exposure to M.
furfur or other inflammatory conditions. Macrophages were infected by M. furfur (1:30) for 20 hours in the pres- ence or absence of methanolic extract of pine needles, including the EtOAc, n-BuOH and water layers which had no cytotoxicity (100 μg/ml pretreatment for 2 h).
Total RNA was then extracted. The effect of the three layers on IL-1β and TNF-α mRNA expression levels in
M. furfur-induced macrophages was measured by real- time PCR and RT-PCR using mouse IL-1β- and TNF-α- specific sense and antisense primers, as shown in Table 2. The relative intensity of each mRNA quantified was normalized against the mRNA expression of human β- Table 2. Sequence of pro-inflammatory gene primer
designs.
IL-1β F 5’-GTG TCT TTC CCG TGG ACC TT-3’
R 5’-TCG TTG CTT GGT TCT CCT TG-3’
TNF-α F 5’-GGC CTC TCT ACC TTG CC-3’
R 5’-TAG GCG ATT ACA GTC ACG GC-3’
GAPDH F 5’-TGC AGC ACC AAC TGC TTA G-3’
R 5’-GGA TGC AGG GAT GTT C-3’
Table 3. Diameter of inhibition zones caused by P. densiflora needle methanolic extract layers (100 mg/ml) in M. furfur culture plates.
PMH PMB PME PMW
mm 1.9 1.3 1.2 1.1 Fig. 4. Tyrosinase inhibition activity of P. densiflora needle methanolic extract layers. PME; Ethyl acetate layer of Pine needle methanolic extract, PMB; n-butyl alcohol layer of pine needle methanolic extract, PMH; n-hexane layer of pine needle methanolic extract, PNW; Water layer of pine needle methano- lic extract. All layers showed dose-dependent tyrosinase inhi- bition activity similar to that exhibited by ascorbic acid.
Fig. 5. Viability of RAW 264.7 cells upon exposure to P. den- siflora needle methanolic extract layers. All layers were applied to the cells for 24 h and cell viability was assessed by MTT assay as described in the text. n-hexane layer and 200 μg/
ml of EtOAc layer were cytotoxic and were excluded from the following experiments.
actin. The expression levels of IL-1β and TNF-α increased in M. furfur-treated macrophages, which con- curs with previously published data [16]. However, as shown Fig. 6A, when macrophages were treated with M.
furfur plus EtOAc layer, the EtOAc layer significantly downregulated the M. furfur-induced IL-1β mRNA expression in macrophages. Accordingly, the results of RT-PCR also showed that pre-treatment with pine nee- dle extract strongly inhibited M. furfur-induced produc- tion of IL-1β in macrophages (Fig. 6B). Moreover, we determined that the EtOAc layer significantly inhibited M. furfur-induced TNF-α mRNA expression level in macrophages (Fig. 7A). Correspondingly, as shown in Fig. 7B, RT-PCR findings indicate that the M. furfur- induced increase in expression levels of TNF-α is attenu- ated by EtOAc layer of pine needle extract among the other layers. Then, Macrophages were infected by M.
furfur (1:30) for 20 hours in the presence or absence of methanolic extract of pine needles, including the EtOAc layer (0−100 μg/ml pretreatment for 2 h). Total RNA was then extracted. The effect of the EtOAc layer on IL-1β
and TNF-α mRNA expression levels in M. furfur- induced macrophages was measured by real-time PCR and RT-PCR. As shown in Fig. 8A, when macrophages were treated with M. furfur plus EtOAc layer, the EtOAc layer significantly downregulated the M. furfur- induced IL-1β mRNA expression in macrophages.
Accordingly, the results of RT-PCR also showed that pre-treatment with pine needle extract inhibited M. fur- fur-induced production of IL-1β in macrophages (Fig.
8B). Moreover, we determined that the EtOAc layer slightly inhibited M. furfur-induced TNF-α mRNA expression level in macrophages (Fig. 9A). Correspond- ingly, as shown in Fig. 9B, RT-PCR findings indicate that the M. furfur-induced increase in expression levels of TNF-α is slightly attenuated by EtOAc layer of pine needle extract in a dose-dependent manner.
Discussion
The purpose of this study was to evaluate the effects of P. densiflora on skin health and its anti-inflammatory Fig. 6. Inhibition of M. furfur-induced IL-1β expression by layers from P. densiflora needle methanolic extract. (A) The mRNA expression level of inflammatory genes was determined by real-time PCR. Treatment of EtOAc layer of P. densiflora strongly reduced mRNA expression level of IL-1β, (B) Treatment of EtOAc layer of P. densiflora strongly reduced mRNA expression level of IL-1β.
Fig. 7. Inhibition of M. furfur-induced TNF-α expression by layers from P. densiflora needle methanolic extract. (A) Treatment of EtOAc layer of P. densiflora strongly decreased mRNA expression level of TNF-α, (B) Treatment of EtOAc layer of P. densiflora strongly decreased mRNA expression level of TNF-α.
Fig. 8. Inhibition of M. furfur-induced IL-1β expression by EtOAc layer from P. densiflora needle methanolic extract. (A) Treat- ment of EtOAc layer of P. densiflora dose-dependently reduced mRNA expression level of IL-1β, (B) Treatment of EtOAc layer of P.
densiflora dose-dependently reduced mRNA expression level of IL-1β.
activity. A wide range of diseases are caused by oxidative stress. Accelerated cell oxidation even leads to wrinkled skin [8]. In recent years, there has been increasing interest in examining natural antioxidants that can protect skin against oxidative stress. In this study, we confirm the antioxidant activity of P. densiflora needles, specifically themethanolic extract. We partitioned four layers from the extract and showed that the BuOH and EtOAc layerspossess the strongest antioxidant activity.
This was in contrast to the findings that the highest scavenging effects are exhibited by the water extract from P. densiflora needlesin a study by Park et al. [29].
The discrepancy may be attributed to the differences in the extraction method.
The skin is colonized by over 500 commensal microbial species estimated to form over 95% [32]. Malassezia, particularly M. furfur, a saprophyte occurring widely on human skin, are generally regarded as the causative agents of a number of common dermatological disorders relate to inflammation [20]. In this study, antifungal
activity against M. furfur was investigated via the clear zone test using paper discs and n-BuOH, EtOAc, and n- hexane layers of P. densifloraneedle methanolic extract were shown to possess fungicidal activity.
Tyrosinase is the enzyme responsible for transfer of the substrate tyrosine into melanin by melanocytes [10].
Inhibition of tyrosinase can lead to reduce or no melanin synthesis and whitening of skin. In our study, all layers of the methanolic extract were found to exhibit tyrosi- nase inhibition activity.
IL-1β and TNF-α are multi-functional cytokines with widely overlapping activities. These inflammatory cyto- kines have a central role in the pathology of chronic inflammatory diseases [30] through their regulation of the immune response to inflammatory stimuli. The mac- rophage is one of the major cell types in inflammatory response and influences many chronic inflammatory dis- eases. IL-1β and TNF-α expression levels in M. furfur- treated macrophages, which concurs with previously published data [16]. Thus, it is of therapeutic interest to Fig. 9. Inhibition of M. furfur-induced TNF-α expression by EtOAc layer from P. densiflora needle methanolic extract. (A) Treat- ment of EtOAc layer of P. densiflora slightly decreased mRNA expression level of TNF-α, (B) Treatment of EtOAc layer of P. densiflora slightly decreased mRNA expression level of TNF-α.
develop an efficient strategy using P. densiflora extract to down-regulate the expression of M. furfur-induced pro-inflammatory cytokines. The mRNA expression lev- els of IL-1β and TNF-α in M. furfur-stimulated RAW 264.7 cells decreased with increasing concentrations of EtOAc layerin the concentration range of 0–100 µg/ml.
We clearly demonstrated significant suppression of IL- 1β levels and a slight attenuation of TNF-α levels by the EtOAc layer in M. furfur-induced RAW264.7 murine macrophages.
In conclusion, the EtOAc and n-BuOH layer of P.
densiflora had antioxidant activity and the various lay- ers of the methanolic extract were found to have antifun- gal activity against the skin pathogen M. furfur.
Additionally, all layers of the extract had tyrosinase inhibition activity. We have also shown that the EtOAc layer partitioned from the methanolic extract of P. densi- flora pine needles has dose-dependent anti-inflamma- tory activity in M. furfur-stimulated Raw 264.7 cells through the down-regulation of IL-1β expression levels.
Thus, it can be inferred that P. densiflora needles and their components have the potential to be used as alter- native anti-inflammatory agents and cosmetic materials for skin health improvement.
Acknowledgments
This work (Grants No. C-035-4093) was supported by Business for Cooperative R&D between Industry, Academy, and Research Institute funded Korea Small and Medium Business Administration in 2015.
References
1. Bäck O, Faergemann J, Hörnqvist R. 1985. Pityrosporum folliculi- tis: a common disease of the young and middle-aged. J. Am.
Acad. Dermatol. 12: 56−61.
2. Bergbrant I, Faergemann J. 1990. Presented at the Seminars in dermatology.
3. Bingham CO. 2002. The pathogenesis of rheumatoid arthritis:
pivotal cytokines involved in bone degradation and inflamma- tion. J. Rheumatol. 65: 3−9.
4. Boncler M, Różalski M, Krajewska U, Podsędek A, Watala C. 2014.
Comparison of PrestoBlue and MTT assays of cellular viability in the assessment of anti-proliferative effects of plant extracts on human endothelial cells. J. Pharmacol. Toxicol. Methods. 69: 9−16.
5. Brunke S, Hube B. 2006. MfLIP1, a gene encoding an extracellular lipase of the lipid-dependent fungus Malassezia furfur. Microbiol- ogy 152: 547−554.
6. Chiba K, Kawakami K, Sone T, Onoue M. 2003. Characteristics of
skin wrinkling and dermal changes induced by repeated applica- tion of squalene monohydroperoxide to hairless mouse skin.
Skin Pharmacol. Physiol. 16: 242−251.
7. Chua KB, Chua I-L, Chua I, Chong K, Chua KH. 2005. A modified mycological medium for isolation and culture of Malassezia fur- fur. Malays. J. Pathol. 27: 99−105.
8. Finkel T, Holbrook NJ. 2000. Oxidants, oxidative stress and the biology of ageing. Nature 408: 239−247.
9. Gaitanis G, Bassukas ID, Velegraki A. 2009. The range of molecu- lar methods for typing Malassezia. Curr. Opin. Infect. Dis. 22: 119− 125.
10. García‐Borrón JC, Solano F. 2002. Molecular anatomy of tyrosi- nase and its related proteins: beyond the histidine‐bound metal catalytic center. Pigment Cell Res. 15: 162−173.
11. Gupta AK, Batra R, Bluhm R, Boekhout T, Dawson TL. 2004. Skin diseases associated with Malassezia species. J. Am. Aca. Dermatol.
51: 785−798.
12. Hort W, Mayser P. 2011. Malassezia virulence determinants. Curr.
Opin. Infect. Dis. 24: 100−105.
13. Jeon H-J, Lee K-S, Ahn Y-J. 2001. Growth-inhibiting effects of con- stituents of Pinus densiflora leaves on human intestinal bacteria.
Food Sci. Biotechnol. 10: 403−407.
14. Jeon MJ, Kim M, Jang H-J, Lee S-W, Kim J-H, Kim H-S, et al. 2012.
Whitening effect of Hizikia fusiformis ethanol extract and its frac- tions. J. Life Sci. 22: 889−896.
15. Keddie FM. 1967. Electron microscopy of Malassezia furfur in tinea versicolor. Med. Mycol. 5: 134−137.
16. Kesavan S, Holland K, Ingham E. 2000. The effects of lipid extraction on the immunomodulatory activity of Malassezia spe- cies in vitro. Med. Mycol. 38: 239−247.
17. Kesavan S, Walters C, Holland K, Ingham E. 1998. The effects of Malassezia on pro-inflammatory cytokine production by human peripheral blood mononuclear cells in vitro. Med. Mycol. 36: 97− 106.
18. Kim H, Han T-H, Lee S-G. 2009. Anti-inflammatory activity of a water extract of Acorus calamus L. leaves on keratinocyte HaCaT cells. J. Ethnopharmacol. 122: 149−156.
19. Kim K-Y, Chung H-J. 2000. Flavor compounds of pine sprout tea and pine needle tea. J. Agric. Food Chem. 48: 1269−1272.
20. Krämer HJ, Podobinska M, Bartsch A, Battmann A, Thoma W, Bernd A, et al. 2005. Malassezin, a novel agonist of the aryl hydro- carbon receptor from the yeast Malassezia furfur, induces apop- tosis in primary human melanocytes. Chembiochem. 6: 860−865.
21. LaDuca JR, Gaspari AA. 2001. Targeting tumour necrosis factor alpha: New drugs used to modulate inflammatory diseases. Der- matol. Clin. 19: 617−635.
22. Lee E. 2003. Effects of powdered pine needle (Pinus densiflora seib et Zucc.) on serum and liver lipid composition and antioxi- dative capacity in rats fed high oxidized fat. J. Korean Soc. Food Sci. Nutr. 32: 926−930.
23. Majdalawieh AF, Hmaidan R, Carr RI. 2010. Nigella sativa modu- lates splenocyte proliferation, Th1/Th2 cytokine profile, macro- phage function and NK anti-tumor activity. J. Ethnopharmacol.
131: 268−275.
24. Masaki H. 2010. Role of antioxidants in the skin: anti-aging effects. J . Dermatol. Sci. 58: 85−90.
25. Mayser P, Preuss J. 2012. Pityriasis versicolor. Der Hautarzt. 63:
859−867.
26. Nenadis N, Tsimidou M. 2002. Observations on the estimation of scavenging activity of phenolic compounds using rapid 1,1- diphenyl-2-picrylhydrazyl (DPPH•) tests. J. Am. Oil Chem. Soc. 79:
1191−1195.
27. Park H-H, Lee S, Oh J-M, Lee M-S, Yoon K-H, Park BH, et al. 2007.
Anti-inflammatory activity of fisetin in human mast cells (HMC- 1). Pharmacol. Res. 55: 31−37.
28. Park J, Choi H, Kim W, Kim H, Cheong H. 2015. Antithrombosis activity of protocatechuic and shikimic acids from functional plant Pinus densiflora Sieb. et Zucc needles. J. Nat. Med. DOI:
10.1007/s11418-015-0956-y
29. Park YS, Jeon MH, Hwang HJ, Park MR, Lee S-H, Kim SG, et al.
2011. Antioxidant activity and analysis of proanthocyanidins from pine (Pinus densiflora) needles. Nutr. Res. Prac. 5: 281−287.
30. Pennanen N, Lapinjoki S, Urtti A, Mönkkönen J. 1995. Effect of liposomal and free bisphosphonates on the IL-1β, IL-6 and TNFα secretion from RAW 264 cells in vitro. Pharm. Res. 12: 916−922.
31. Rosenberg EW, Noah PW. 1988. The Koebner phenomenon and the microbial basis of psoriasis. J. Am. Acad. Dermatol. 18: 151−
158.
32. Round JL, O'Connell RM, Mazmanian SK. 2010. Coordination of tolerogenic immune responses by the commensal microbiota. J.
Autoimmun. 34: J220−J225.
33. Ryu S, Choi S-Y, Acharya S, Chun Y-J, Gurley C, Park Y, et al. 2011.
Antimicrobial and anti-inflammatory effects of cecropin A (1-8)–
Magainin2 (1-12) hybrid peptide analog P5 against Malassezia furfur infection in human keratinocytes. J. Invest. Dermatol. 131:
1677−1683.
34. Schottelius AJ, Moldawer LL, Dinarello CA, Asadullah K, Sterry W, Edwards CK. 2004. Biology of tumor necrosis factor‐α–implica- tions for psoriasis. Exp. Dermatol. 13: 193−222.
35. Tengamnuay P, Pengrungruangwong K, Pheansri I, Likhitwitay- awuid K. 2006. Artocarpus lakoocha heartwood extract as a novel cosmetic ingredient: evaluation of the in vitro anti‐tyrosi- nase and in vivo skin whitening activities. Int. J. Cosmet. Sci. 28:
269−276.
36. Trent JT, Kerdel FA. 2005. Tumor necrosis factor alpha inhibitors for the treatment of dermatologic diseases. Dermatol. Nurs. 17:
97.
37. Villa FA, Gerwick L. 2010. Marine natural product drug discovery:
Leads for treatment of inflammation, cancer, infections, and neu- rological disorders. Immunopharmacol. Immunotoxicol. 32: 228− 237.
38. Waersted A, Hjorth N. 1984. Pityrosporum orbiculare--a patho- genic factor in atopic dermatitis of the face, scalp and neck? Acta dermato-venereologica. Supplementum. 114: 146−148.
39. Wang H, Gao XD, Zhou GC, Cai L, Yao WB. 2008. In vitro and in vivo antioxidant activity of aqueous extract from Choerospon- dias axillaris fruit. Food Chem. 106: 888−895.
40. Yuan G, Wahlqvist ML, He G, Yang M, Li D. 2006. Natural products and anti-inflammatory activity. Asia Pac. J. Clin. Nutr. 15: 143.
국문초록
솔잎추출물의 피부건강 개선효과 최지은, 김웅, 박재영, 정현숙*
조선대학교자연과학대학생명과학과
솔잎은동의보감과향약생약대사전에간, 위, 심, 피부등질환치료에효과가있다고하여예로부터솔잎을노화방지와피부질환
치료에활용하여왔다. Malassezia furfur (M. furfur)는친유성곰팡이로인간피부에존재하는흔한균이다. 이균은대게피부
의각질층에서주로발견되며, 어우러기, 지루성피부염, 비듬, 모낭염, 건성등의각종피부질환의주요요인이다. 본연구는사람 의대부분의염증실험에서사용되는 Raw 264.7 세포를이용하여솔잎의항염증활성을확인하였다. 솔잎을에탄올, 물, 메탄올 로추출하여항산화효과를확인한결과, 메탄올추출물에서항산화능이높게나타났으며, 이에따라헥산, 에틸아세테이트, 부탄 올, 물층으로나누었다. 이 4층을이용하여노화의원인이되는자유라디칼의소거능을확인한결과, 부탄올과에틸아세테이트
층에서강한항산화활성이나타났으며, M. furfur에대한항균활성을확인한결과부탄올, 에틸아세테이트층과헥산층에서항
균활성이나타났다. 또한티로시나아제저해에따른미백활성은 4층모두대조군인비타민C와유사하게나타났다. 세포실험을
위해먼저대식세포인 Raw 264.7에솔잎분획물을처리하여세포독성을확인한결과 Hexane층에서농도별로높은독성이나 타났으며나머지층에서는독성이나타나지않음을확인하였다. 이후 Malassezia furfur로염증반응이유도된 Raw 264.7에에 틸아세테이트, 부탄올, 물층을처리하여염증에관련된유전자들의발현량을 Real-Time PCR을통하여확인하였다. 이때사용된
유전자는 IL-1β, TNF-α로에틸아세테이트층이염증과깊은연관이있는 IL-1β와 TNF-α의발현량을낮추는것으로확인되었
다. 모든실험결과들을종합해보았을때, 솔잎은천연자원으로써피부건강과관련된건강식품, 건강보조제, 화장품등폭넓은
범위로사용될수있을것으로사료된다.