http://www.medicinalcrop.org http://dx.doi.org/10.7783/KJMCS.2013.21.5.361
맥문동 종실 안토시아닌 분획물의 멜라닌 생성 억제 및 미백 효과
정명근*·황영선*·김기쁨*·안경근*·심훈섭*·홍승범*·최재후**·유창연**·정일민***
김승현***·임정대*†
*강원대학교 생약자원개발학과, **강원대학교 식물자원응용공학과 ***건국대학교 응용생물과학과
Antimelanogenic Effect and Whitening of Anthocyanin Rich Fraction from Seeds of Liriope platyphylla
Myoung Gun Choung*, Young Sun Hwang*, Gi Ppeum Kim*, Kyung Geun Ahn*, Hoon Seob Shim*, Seung Beom Hong*, Jae Hoo Choi**, Chang Yeon Yu**, Ill Min Chung***,
Seung Hyun Kim*** and Jung Dae Lim*†
*Department of Herbal Medicine Resource, Kangwon National University, Samcheok 245-905, Korea.
**Department of Applied Plant Sciences, Kangwon National University, Chuncheon 200-701, Korea.
***Department of Applied Bioscience, Konkuk University, Seoul 143-701, Korea.
ABSTRACT : This study was performed to determine the antimelanogenic effect and tyrosinase inhibitory activities of anthocyanin rich fraction (AN-SLP) from Liriope platyphylla Wang et Tang seeds. Anthocyanins isolated from L. platyphylla seeds revealed the presence of four major anthocyanin components, which were tentatively identified as delphinidin-3- O- glucoside, delphinidin-3- O-rutinoside, petunidin-3-O-rutinoside, and malvidin-3-O-rutinoside using semipreparative HPLC,
1
H-NMR,
13C NMR, FAB-MS and LC/ES-MS. The inhibitory effect of AN-SLP on tyrosinase activity was studied using in vitro (against mushroom tyrosinase) and ex vivo (against B16 melanoma cell tyrosinase) models. Cellular tyrosinase activity was decreased by AN-SLP treatment in B 16 melanoma cells through dose dependent manner, but AN-SLP did not inhibit mushroom tyrosinase and L-DOPA oxidation directly. AN-SLP showed melanin inhibition by 53.2% at 50 ㎍/㎖ which was 0.7 times more efficient than the antimelanogenic effect of commercial arbutin and kojic acid (36.5%) also did not show cell toxicity. Additionally, AN-SLP inhibited the activity of α-glucosidase and the glycosylation of tyrosinase in melanoma cell.
The resulting unsaturated glycosylation of tyrosinase makes it unstable and disturb correct transportation. From theses results, we conclude that AN-SLP could be used as anti-melanogenic agent for skin whitening.
Key Words : Anthocyanin Rich Fraction, Antimelanogenic Effect, Seeds of Liriope platyphylla, Tyrosinase Inhibitory Activity
서 언
멜라닌 (melanin)은 적갈색 (pheomelanin) 또는 흑갈색 (eumelanin) 의 고분자화합물로서 주로 흑색이나 갈색을 나타내 는 유멜라닌 (eumelanin)과 노란색이나 적색 (red)을 나타내는 페오멜라닌 (pheomelanin)이 있다. 멜라닌세포 (melanocyte) 내 멜라닌소체 (melanosome)라는 소기관에서 합성 된다 (Hearing, 1999). 합성된 멜라닌색소는 멜라닌세포의 수지상돌 기 (dendrite)를 통하여 인접세포인 각질세포 (keratinocyte) 로
전달되며, 전달된 멜라닌색소는 각질세포를 통하여 피부 내 여 러 부위에 분포하게 된다 (Lowell, 1991).
멜라닌 합성 과정을 살펴보면, 멜라노사이트 내의 멜라노좀 이라는 소포체에서 먼저 티로시나제 (Tyrosinase)라는 효소에 의해 티로신이 도파 (DOPA)를 거쳐 도파퀴논 (DOPA quinone) 으로 전환되고, 도파퀴논으로부터 자동 산화 반응과 효소 반응으로 도파크롬 (DOPA chrome)을 거쳐 공중합체인 멜라닌이 생성된다 (Alaluf et al., 2001). 멜라닌은 햇빛에 포 함된 UV의 빛 에너지를 흡수하여 UV에 의한 손상으로부터
†
Corresponding author: (Phone) +82-33-540-3323 (E-mail) [email protected]
Received 2013 August 27 / 1st Revised 2013 September 11 / 2nd Revised 2013 September 23 /3th Revised September 26 / Accepted 2013 Revised September 26
This is an open access article distributed under the terms of the Creative Commons Attribution Non-Commercial License (http://creativecom-
mons.org/licenses/by-nc/3.0/) which permits unrestricted non-commercial use, distribution, and reproduction in any medium, provided the original
work is properly cited.
진피 이하의 피부 기관을 보호하는 역할을 하며, 피부 생체 내에 생겨난 유해산소 및 유리기 (free radical) 등을 잡아주는 등 외부 유해인자로부터 피부를 보호해주는 유용한 역할을 수 행한다. 그러나 멜라닌이 비정상적으로 소량 생산되면 백반증 과 같은 피부병변이 유발되고, 반대로 일광, 호르몬 변화, 염 증 또는 약제 등 여러 가지 환경적 요인에 의해 멜라닌이 과 도하게 합성되거나 색소가 침착되면 기미, 주근깨를 형성하며, 이와 같은 병변으로부터 나아가 피부암의 원인이 되기도 한다.
그러므로 이러한 색소 침착 현상을 방지하고 미백효과를 얻기 위해서는 멜라닌 생성 과정의 일부분을 저해하여 멜라닌의 생 성을 감소시켜야 한다.
현재 화장품 분야에서는 미백 성분으로서, 코지산 (Kojic acid), 알부틴 (Arbutin) 등과 같은 tyrosinase 효소 활성을 억 제하는 물질, 하이드로퀴논 (Hydroquinone), 비타민 C (L- Ascorbic acid) 등이 사용되어 왔는데, 이러한 물질들의 조성 은 안정성이 열악하여 분해 혹은 착색되거나, 이취의 발생, 불 안정성, 생체 레벨에서의 효능의 여부, 부작용 효과의 불분명 및 안전성 문제 등으로 그 사용이 제한되고 있는 실정이다 (Kim and Kim, 2008). 이러한 성분 외에도 천연물, 특히 식 물 중에서 멜라닌 생성 억제 및 색소 침착 현상을 방지할 수 있는 활성소재로 녹나무 (Ha et al., 2009), 백삼 (Jo et al., 2008), 죽황 (Lee et al., 2008), 홍화 (Roh et al., 2004), 뽕 잎 (Lee et al., 2002), 닥나무 (Jeon et al., 2001), 오디 (Jang et al., 1997) 등 다수의 식물 추출물 및 생약재 추출물 이 연구 되었는데, 이들 역시 안전성, 변색 가능성 등의 측면 에서 화장품이나 의약품에 유효 농도 이상으로 사용하는 데는 많은 문제점이 있으며, 형성되어진 tyrosinase에 대하여 직접적 인 소거 작용을 하는 메카니즘이 알려져 있을 뿐 tyrosinase 단백질의 번역단계 또는 발현 수준에서 저해활성을 가지는 소 재에 대한 검토는 미약한 수준이다. 이에 따라 tyrosniase에 대한 직접적인 소거 기능 뿐만 아니라 번역 및 발현 단계에서 효소의 형성 자체를 억제할 수 있는 효능이 뛰어난 천연소재 의 미백제가 절실히 요구되고 있다.
Tyrosinase 는 type I membrane glycoprotein으로서 사람은 7 개의 glycosylation site를 가지고 있으며 (Branza-Nichita et al., 2000) N-linked glycosylation 과정에 의해 만들어진다. 단 백질의 N-linked glycosylation은 endoplasmic reticulum (ER) 에서 단백질 합성과 함께 일어난다 (Petrescu et al., 1997).
Glycan unit 의 절반인 14개짜리의 당 (Glc
3Man
9GlcNAc
2) 은 ER 막의 바깥인 세포질쪽에서 만들어지며 이후 나머지 절반은 ER 내에서 만들어져 단백질로 전이 된다 (Kornfeld and Kornfeld, 1985). ER 에서 이런 합성 과정은 oligosaccharide의 말단 끝에 존재하는 mannose에서 3개의 glucose를 α- glucosidase I 과 II가 끊어줌으로서 시작되게 된다 (Winchester and Fleet, 1992). Tyrosinase 의 glucose trimming 과정은
castanospermine 이나 deoxynojirimycin (DNJ), N-butyl- deoxynojirimycin (NBDNJ) 과 같은 α-glucosidase의 저해제에 의해서 저해된다. 이 때 tyrosinase가 급속하게 folding되어 copper 가 없는 불활성화된 형태가 생성되어 melanosome으로 이동하게 된다 (Branza-Nichita et al., 1999). 이와 같이 α- glucosidase 저해제는 결과적으로 멜라닌의 합성을 저해한다고 알려져 있다 (Petrescu et al., 1997).
식물체는 일반적으로 UV에 노출로부터 조직을 보호하기 위 하여 많은 양의 phenolic acids, phenyl propanoids, 간단한 flavonoids 등의 polyphenolic compounds를 포함하고 있으며, 이러한 phenolic compounds 중 꽃, 과실, 줄기, 잎, 뿌리 등 식물체 각 부위에 폭넓게 함유되어 있는 안토시아닌 색소가 anti-tyrosinase activity 를 나타낸다는 연구 결과가 오디 (Aramwit et al., 2010), 검정쌀 껍질 (Miyazawa et al., 2003), 포도주 (Gómez-Cordovés et al., 2001) 등에서 보고된 바 있고, 이러한 안토시아닌의 구조 중 acylate가 첨가된 anthocyanin 의 경우 높은 α-glucosidase 저해활성을 나타낸다 고 보고하였다 (Matsui et al., 2001).
맥문동 (Liriope platyphylla Wang et Tang)은 백합과에 속하 는 다년생 초본식물로써 7월에 보라색의 꽃이 피어 10월에는 검은색 열매를 맺는데 씨방상위이며, 열매는 삭과로 둥글고 일 찍 과피 (果皮)가 벗겨지므로 종자가 노출되며 자흑색 (紫黑色) 이다. 맥문동의 약용부위는 뿌리의 끝이 땅콩처럼 굵어져 생 긴 괴근으로, sortase 활성 저해력에 의한 항미생물제 (Katon et al., 2009)의 기능과 천식 마우스 모델에서 airway inflammation 과 hyperresponsiveness의 저해효과, Th1/Th2 cytokine 의 불균형을 조절하는 immunomodulator로서의 기능 (Krentz et al., 2008), neurotrophic factor의 분비촉진을 통한 신경세포의 활성화, 신경세포의 분화와 성장에 중요한 NGF (Nerve Growth Factors) 의 분비 촉진 (Choi et al., 2010) 등 다양한 연구가 이루어졌으나, 맥문동 종실의 효능이나 약용 가 능성에 대한 연구는 매우 미흡한 실정이다. 최근 맥문동 열매 추출물에서 안토시아닌인 petunidin-3-O-rutinoside와 malvidin- 3- O-rutinoside가 높은 수준으로 함유되어 있고, delphinidin-3- O-glucoside, delphinidin-3-O-rutinoside, cyanidin-3-O-glucoside, petunidin-3- O-glucoside 및 malvidin-3-O-glucoside이 포함되어 있으며 이들이 높은 항산화 활성을 가진다는 정량 및 정성 분 석에 대한 결과가 보고되었다 (Lee and Choung, 2011).
따라서 본 연구는 맥문동의 종실 추출물로부터 안토시아닌
분획을 획득하고, 획득된 안토시아닌 분획의 조성을 확인하며,
이들에 대한 in vitro tyrosinase inhibition assay와 in vitro
DOPA oxidation inhibition assay 를 수행하였으며, melanin
합성에 관여하는 tyrosinase에 직접 작용하여 활성을 억제하는
지 혹은 세포수준에서 억제하는지를 알아보기 위해 B16
melanoma cell 를 이용하여 세포독성과 ex vivo tyrosinase
inhibition assay 및 α-glucosidase 효소 저해활성 연구를 수행 하였고, 멜라닌 생성과정의 최종 산물인 멜라닌생성의 억제정 도를 확인함으로써 맥문동 종실 추출물을 멜라닌 생성 억제 및 미백기능성 화장품 소재로 활용하기 위한 기초 자료를 제 공하고자 하였다.
재료 및 방법
1. 실험재료
본 실험에 사용된 맥문동 종실은 충남 청양군 농업기술센터 에서 재배된 ‘청심’ 품종을 대상으로 하였으며, 2009년 11월 에 과육이 완숙되고 완전히 건조되어 과피가 벗겨지고 종자가 노출되어 있는 형태의 단단한 자흑색의 종실을 사용하였다.
2. 맥문동 종실 추출물 제조 및 안토시아닌 분획의 획득 맥문동 종실 100 g을 대상으로 안토시아닌의 기본골격에 포 함되어진 flavylium cations의 안정화을 통해 안토시아닌을 효 율적으로 추출하기 위해 0.1% 염산이 함유된 20% 에탄올이 함유된 용액 3L를 첨가한 후 저온 (4℃) 암조건 하에서 48 시간 동안 방치하여 추출하였다 (Lozovskaya et al., 2012).
추출물은 advantec Toyo No.2 여과지로 여과하였으며, 잔류물 은 3회 반복 추출하였고, 추출물은 모두 합쳐 30℃ 감압농축 장치 (rotary evaporator)에서 농축하였다. 농축된 맥문동 종실 추출물은-70℃ 조건에서 동결한 후, 동결 건조하여 시료로 사 용하였다. 농축된 맥문동 종실 추출물을 Zorbax SB-C18 컬럼 (250 × 9.4 ㎜, 5 ㎛; Agilent Technologies, Wilmington, DE) 이 장착된 semipreparative HPLC를 이용하여 분획을 실시하 였고, 이때 검출기의 파장은 520 ㎚로, 컬럼의 온도는 30℃로 제한하여 분획을 실시하였다. Semipreparative HPLC의 분획조 건은 2가지 용매를 gradient elution system으로 조절하였으며, 용매 A는 5% formic acid을 함유한 증류수로, 용매 B는 5%
formic acid 를 함유한 acetonitrile로 하였다. 분획용매의 유속은 분당 3 ㎖로 조절하였고, 분취를 위한 용매조건은 0 ~ 10 분까 지 용매 B를 10 ~ 18%로, 10 ~ 18 분까지 18 ~ 28%, 18 ~ 19 분까지 28 ~ 40%, 19 ~ 21 분까지 40%, 21 ~ 23 분까지 40 ~ 10%, 23 ~ 25 분까지 10%로 변화시켜 사용하였다.
분취 시 시료 주입량은 2 ㎖로 조절하였고, 맥문동 종실의 추출물로부터 총 4종의 주요 안토시아닌을 분리하였다. 분리 된 4종의 주요 안토시아닌의 상대순도는 TSK gel ODS-120T 컬럼 (150 × 4.6 ㎜, 5 ㎛; Supelco Inc, Bellefonte, PA)을 이 용한 RP-HPLC를 사용하여 순도를 검정하였고, 모두 98%이 상의 순도를 확인하였다. RP-HPLC의 분석조건은 유속을 분 당 0.7 ㎖로 조절하였고 상기 기술된 semipreparative HPLC의 용매와 동일한 gradient elution 조건으로 분석을 실시하였다.
검출기는 photodiode array detection 방식으로 파장을 530 ㎚
으로 조절하였고, 시료 주입량은 20 ㎕로 조절하였다. 주입 시 분석용 시료는 0.45 ㎛ 여과막 필터 (membrane filter)로 여과 를 실시하였고, 분석용 컬럼을 보호하기 위해 Nova-Pak C
18guard insert column (Waters, Milford, MA) 을 사용하여 분석 을 실시하였다.
Semipreparative HPLC 에 의해 용출 분획된 맥문동 종실 안 토시아닌 분획 (Anthocyanin rich fraction form seeds of L.
platyphylla, AN-SLP)만을 별도로 회수하였다. 분획 회수된 맥 문동 종실 안토시아닌 분획을 감압 농축하고 질소충전 하 에 -4℃에 보관하였고 증류수로 희석하여 멜라닌 생성 억 제 및 미백 활성 검정에 사용하였다.
3. 맥문동 종실 안토시아닌 분획에서의 안토시아닌 종류 의 확인
맥문동 종실 안토시아닌 분획에서 안토시아닌의 정성적 평 가를 위하여
1H-NMR,
13C NMR, FAB-MS, LC/ES-MS 분 석을 실시하였는데,
1H-NMR (500 MHz) 과
13C NMR (125 MHz) 스펙트럼은 Varian Unity Plus 500 NMR을 사용하였 으며, 내부표준물질로서 TMS를 함유한 0.5% DCl-CD
3OD 를 용매로 측정하였다. FAB-MS의 측정은 기질로서 NBA (m- NO
2-Benzyl-OH) 를 사용하고, Jeol사의 JIMS 700 고해상 질 량분석기 (High Resolution Mass Spectrometer)를 이용하여 분자 량을 측정하였다. LC/ES-MS 분석은 Spectra system P-4000 HPLC 시스템에 Thermo Finnigan AQA single-quadrupole mass spectrometer 를 결합하여 on-line 상태로 분자량을 측정하였다.
4. 맥문동 종실 안토시아닌 분획의 mushroom tyrosinase 효소 저해 활성 검정
맥문동 종실 안토시아닌 분획의 Tyrosinase 효소 활성 저해 검정은 Vanni 등 (1990)에 의한 방법으로 수행하였다.
0.1 ㎎/㎖의 L-tyrosine을 포함하는 0.05 mM sodium phosphate buffer (pH 6.8) 0.5 ㎖에 농도별 (5 ㎍/㎖, 500 ㎍/㎖)로 조제된 맥문동 종실 안토시아닌 분획물 0.5 ㎖를 첨가하였다. 여기에 다 시 0.5 ㎖의 mushroom tyrosinase (70 units/0.5 ㎖)을 첨가하여 조제된 혼합물을 37℃에서 10분간 반응시킨 후 475 ㎚ (UV- 1200; Shimadzu, Kyoto, Japan) 에서 흡광도를 측정하였다. 맥문동 종실 안토시아닌 분획물을 첨가하지 않은 동일한 혼합물을 대조 구로 사용하였다. Tyrosinase 효소 활성 저해율은 다음 계산식에 의하여 산출하였으며 L-tyrosine과 경쟁적으로 작용하는 저해제 이며 (Maeda and Fukuda, 1996) 미백제로 가장 많이 사용되는 알부틴 (Sigma Chemical Co. St. Louis, USA)과 비교하였다.
% 저해율 = (A-B)/A × 100
A: 시료를 첨가하지 않았을 때의 흡광도
B: 시료를 첨가하였을 때의 흡광도
5. 맥문동 종실 안토시아닌 분획의 DOPA auto-oxidation 저해활성 검정
맥문동 종실 안토시아닌 분획의 DOPA auto-oxidation 저해 활성 검정은 Joshi 등 (1987)에 의한 방법을 변형하여 사용하 였다. 반응액의 조제는 500 mM의 L-DOPA를 포함하는 0.05 M sodium phosphate buffer (pH 6.8) 1 ㎖ 에 동일한 buffer 로 농도별 (100, 200, 400, 800, 1000 ㎍/㎖)로 희석된 맥문 동 종실 안토시아닌 분획 1 ㎖를 첨가하고 2일간 반응시켰다.
반응된 혼합물을 475 ㎚ (UV-1200; Shimadzu, Kyoto, Japan) 에서 흡광도를 측정하였으며 맥문동 종실 안토시아닌 분획물을 첨가하지 않은 동일한 혼합물을 대조구로 사용하였 고 DOPA auto-oxidation 에 대한 저해율은 다음 계산식에 의하여 산출하였으며 DOPA oxidation의 저해 메카니즘을 가진 kojic acid (Sigma Chemical Co. St. Louis, USA) 와 비교하였다.
효소활성저해도(%) = (ΔA0 - ΔA1)/ΔA0 × 100
ΔA0: 시료를 첨가하지 않은 대조군의 흡광도 변화 ΔA1: 시료를 첨가한 처리군에서의 흡광도 변화
6. 맥문동 종실 안토시아닌 분획의 B16 melanoma cell에 대한 세포독성 검정
멜라닌을 생산하는 세포주이며 일반적으로 미백물질 스크리 닝에 사용되는 B16 melanoma 세포주를 한국 세포주 은행에 서 분양받아 사용하였다. B16 melanoma 세포 배양에 사용된 배지는 10% FBS (fetal bovine serum), penicillin G (100 IU/ ㎖), streptomycin (100 ㎍/㎖)을 포함한 DMEM (Dulbecco's modified Eagle's medium, Bio-Whittaker Co. Walkersville, USA), 배지를 사용하였으며, 온도 37℃와 5%의 CO
2를 유지하 면서 incubator에서 배양 하였다.
B16 melanoma cell 에 대한 compounds의 처리농도를 결정 하기 위한 MTT assay는 Mosmann (1983) 에의 방법을 변형 하여 사용하였다. 이 분석법은 노란색의 수용성 기질인 MTT (3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyl-tetrazolium bromide) 를 진청색의 비수용성인 formazan 물질로 변화시키는 살아있 는 세포의 mitochondria dehydrogenase의 능력을 이용한 방법 으로서 생성된 formazan의 양을 측정하여 살아있는 세포의 생 존률을 검정하는 방법이다. B16 melanoma 세포주를 각 well 에 5 × 10
4개의 농도로 넣어준 다음 1일 후 맥문동 종실 안 토시아닌 분획물을 농도별 (5, 50, 100, 500 ㎍/㎖) 로 넣어준 배지로 교환하였다. 맥문동 종실 안토시아닌 분획물을 처리한 지 3일 후에 PBS (phosphate buffer saline)로 세척하여 주고, 2 ㎎/㎖ 농도의 MTT 용액을 40 ㎕ 첨가하고 4 시간 동안 반
응시켰다. (37℃, 5 % CO
2incubator). 배양 중 불용성의 blue formazane 이 생성되는데 이는 각 세포주에 따라 일정하 게 살아 있는 세포 수에 비례하여 형성된다. 반응 후 각 well 을 1,000 rpm으로 원심분리하고 각 well에 들어 있는 여 분의 배지를 제거하였으며, DMSO를 well 당 150 ㎕를 넣고 30 분 동안 교반 용해시켰다. 반응 후 ELISA microplate reader 를 이용 540 ㎚에서 흡광도를 측정하였다. 항미백 소재로 가장 많이 사용되고 있으나 멜라닌 세포에 대한 독성 및 색소 침착 등의 부작용을 가지고 있어 제한적 농도에서의 사용이 허 가되어 있는 hydroquinone (Aldrich Chemical Co. Milwaukee, USA) 을 5 ㎍/㎖ 로 처리하여 대조구로 사용하였고 arbutin과 kojic acid 를 50 ㎍/㎖의 수준으로 처리하여 비교하였다.
7. B16 melanoma cell 내에서의 맥문동 종실 안토시아닌 분획물의 tyrosinase 저해활성 및 멜라닌 생성 저해량 측정
세포를 배양하여 24 well plate에 각 well당 세포를 1 × 10
5개씩 넣고 mushroom tyrosinase 효소에 대해 저해 활성을 나 타내었던 맥문동 종실 안토시아닌 분획 시료를 농도별 (5, 50, 100, 500 ㎍/㎖)로 처리한 후 48 시간 배양하였다. 48 시간 배양한 후 각 well의 세포를 10 mM PBS로 세척 하였으며, Triton X-100 1% 를 함유한 10 mM PBS 100 ㎕에 현탁시켰 다. 현탁된 이 액을 vortexing한 후 원심분리하여 상징액을 효 소액으로 사용하였다. 96 well plate에 이 효소액을 40 ㎕ 넣 고 기질인 L-dopa (2 ㎎/㎖) 100 ㎕ 를 첨가하였다. 37℃에서 1 시간 동안 반응을 진행시킨 뒤, ELISA reader를 이용하여 405 ㎚에서 흡광도를 측정하였다. tyrosinase의 활성도는 대조 군의 흡광도에 대한 백분율로 계산하였으며 양성대조군으로 arbutin 을 동일한 농도로 적용하였다.
멜라닌생성량 측정은 DMEM으로 계대배양 된 B16 melanoma 세포주를 100 ㎜ culture dish에 2 × 10
5개의 농도로 넣어준 후 1일 뒤 에 맥문동 종실 안토시아닌 분획을 농도 별 (5, 50, 100, 500 ㎍/㎖) 로 처리하였다. 3일째에 세포를 인산완충액 (pH 7.4)으로 세척하고 0.25 M trypsin-EDTA 용액으로 세포를 바닥에서 떼어준 후, DMEM으로 trypsin- EDTA 의 작용을 중화시켰다. 10 min 동안 2,500 rpm, 4℃
조건에서 원심분리하여 세포를 수확하였다. Hematocytometer 로 B16 세포를 세어서 1 × 10
6세포 수 당 1 ㎖ 의 5%
TCA 로 처리 교반하였다. 2,500 rpm으로 2회 원심분리한 후
분리된 멜라닌을 인산완충액으로 세척하였다. 에테르와 에탄
올을 1:3 비율로 하여 2회 원심분리한 후 에테르 1 ㎖로 세
척 건조시켰다. 건조된 멜라닌에 1 N NaOH를 가하여 1시간
동안 56℃ 항온조에서 반응시킨 후 분광광도계로 475 ㎚ 에
서 흡광도를 측정하고 멜라닌 양은 합성 멜라닌을 사용하여
작성된 표준 직선에서 구하였고 맥문동 종실 추출물 처리군의
멜라닌 양은 대조군의 멜라닌 양에 대한 백분율로 계산하였다.
8. 맥문동 종실 안토시아닌 분획물의 α-glucosidase 저해활 성 검정
동물의 소장 점막에서 분비되는 탄수화물소화효소의 일종인 α-glucosidase는 Kim 등(2000)이 사용한 방법에 따라 기질로서 p-nitrophenyl- α-D-glucopyranoside를 사용하고 효소원으로는 rat intestinal powder (Sigma I 1630) 로부터 추출하여 제조한 α-glucosidase를 이용하여 용매만을 넣은 대조군 (ΔA0)과 맥문 동 종실 안토시아닌 분획을 넣어 반응시킨 처리구의 흡광도변 화 (ΔA1)를 관찰하여 시료의 효소 저해 활성정도를 평가하였 다 즉 rat intestinal α-glucosidase 효소액을 0.2% bovine serum albumin 과 0.02% NaN
3를 함유하는 0.1M sodium phosphate buffer (pH 7.0) 에 0.7 U/㎖가 되도록 용해하였다.
Rat intestinal α-glucosidase 효소액은 100 ㎎의 rat intestinal powder 를 0.9% NaCl 용액 3 ㎖에 가하여 ice bath에서 30초 간 12회 sonication하여 효소를 추출하고 10,000 g에서 30분간 원심분리하여 제조하였으며 상층액을 효소저해활성측정에 사 용하였다 (Oki et al., 1999). 기질은 ρ-nitrophenyl-α-D- glucopyranoside (Sigma N 1377) 를 0.1M 인산완충액 (pH 7.0) 에 5 mM 농도로 용해하여 사용하였다. 효소액 50 ㎕와 농 도별 (5, 50, 100, 500 ㎍/㎖) 맥문동 종실 안토시아닌 분획물 ( 대조군의 경우 추출에 사용한 용매) 10 ㎕를 96-well plate에 넣고 405 ㎚에서 흡광도를 측정하고 (A at time = 0), 5분간 실온에 방치한 후 기질을 50 ㎕ 가하고 실온에서 5분간 반응 시킨 후 다시 405 ㎚에서 흡광도 (A at time = 5 min)를 측정 하였다. 양성 대조군으로 acarbose (AC)를 사용하였으며 효소 활성의 저해정도는 다음 식에 의하여 산출하였다.
효소활성저해도(%) = (ΔA0-ΔA1)/ΔA0 × 100 ΔA0: 시료를 첨가하지 않은 대조군의 흡광도 변화 ΔA1: 시료를 첨가 한 처리군에서의 흡광도 변화
9. 통계처리
맥문동 종실 안토시아닌 분획물의 농도별 처리에 따른 in vitro tyrosinase 저해활성과 in vitro DOPA oxidation 저해 효과, B16 melanoma cell을 이용하여 세포독성과 ex vivo tyrosinase 저해효과 및 α-glucosidase 효소 저해활성, 멜라닌 생성의 억제정도를 확인하기 위한 실험에서 얻어진 각 자료는 SAS 프로그램 (Statistical software, SAS institute)을 이용하 여 ANOVA에 의한 Duncan (DMRT, Duncan Multiple Range Test) 검정을 수행 하였다.
결과 및 고찰
1. 맥문동 종실 안토시아닌 분획의 정성적 확인
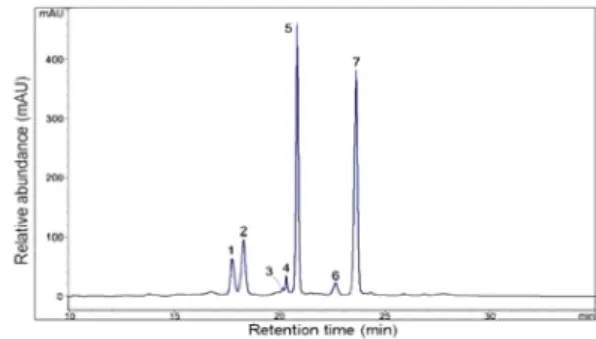
Semipreparative HPLC 를 이용하여 맥문동 종실에서 안토시
아닌 색소를 분리한 결과 총 7가지의 안토시아닌이 분리되었 다 (Fig. 1). 상기 HPLC를 통하여 맥문동 종실 안토시아닌 분획에서 총 7종의 피크를 확인하였고, 분리된 7종 색소성분 의 UV-VIS 및 MS data와 화학구조식은 Table 1에 나타내었 으며, 그 중 함량이 높은 화합물 1, 2, 5, 7에 대해 구조 동 정을 실시하였다.
화합물 (compound) 1은 NMR, LC/ES-MS, FAB-MS, 및 UV/Vis spectral data 분석 결과 delphinidin 3-glucoside임을 확인하였다. δ 8.95 (H-4)에서 1H singlet, δ 7.7 6(H-2', 6') 에서 2H singlet, δ 6.65 (H-6)와 6.87 (H-8)에서 2H AX system 이 보이는 화합물 1의 1차원
1H-NMR 스펙트럼에서 aromatic region 은 delphinidin 핵과 일치하였다.
13C spin echo Fourier transform NMR spectrum 에서 δ 60 ~ 80 사이 의 스펙트럼 영역은 한 개의 hexose와 일치하는 anomeric carbon 과 함께 다섯 개의 공명구조 (resonance)를 가졌다. One bond heteronuclear shift correlation NMR 에서 나타난
1H 와
13
C shift value 와 iterative spin simulation (PANIC)에 의해 밝혀진 당 고리 양성자 (sugar ring proton) (7 ~ 10Hz)의
1H-
1
H 짝지움 상수 (coupling constant)는 β-D-glucopyranoside와 일치하였다. 당의 결합 사이트는 HMBC에서 anomeric proton 과 aglycon C-3사이의 long range correlation peak를 통해 aglycon 의 3번 위치임을 확인하였다. 또한 On-line HPLC에서 얻어진 화합물 1의 UV-VIS 스펙트럼은 526 ㎚에서 흡수극대 를 보이고 있으며, 이는 delphinidin 핵을 가진 3-glucoside와 일치하였다. 화합물 1의 LC/ES/MS와 FAB-MS spectrum에 서는 m/z 465에서 delphinidin hexose와 일치하는 [M
+] 를 얻었 고, m/z 303에서 delphinidin aglycon의 존재를 의미하는 베이스 피크를 얻었으며 화합물 1의 NMR 데이터는 Delphinidin-3-O- glucoside 의 문헌상의 값과 일치하였다 (Cerezo et al., 2010).
화합물 (Compound) 2는 0.5% HCl-MeOH 용액에서 UV- VIS 측정 결과 λ max가 278과 530 ㎚에서 나타났고, FAB- MS 에서는 m/z 611에서 [M
+] 를 얻었다. 이는 delphinidin 3- glucoside (m/z 465), delphinidin (m/z 303) 과 일치하는 플레
Fig. 1. HPLC profile of anthocyanins fraction in seed of L.
platyphylla detected at 530 ㎚.
그먼트 (fragment)를 가진 C
27H
31O
16+의 분자식을 가진다. 화합 물 2의 aglycone인 delphinidin의 양성자와 carbon signal은 짝지 움 상수와 chemical shift에 관한 정보를 이용하여 나타내었다.
Hexose 의 동정은 1차원
1H NMR 스펙트럼과
1H-
1H COSY 로 행하였다. 당에 인접한 모든 짝지움 상수는 δ 5.32 (d, J = 7.8Hz) 에 아노머 양성자를 포함한 7.8-9.5Hz에서 나타났다.
따라서 당은 베타-디-글루코피라노사이드 (β-D-glucopyranoside) 이다. 글루코실(δ 5.32) H-1''의 조사 (irradiation)에 의해 핵의 H-4 ( δ 8.87)에서 강한 negative NOE가 관찰되었는데, 이것은 해당 당 잔기 (sugar moiety)가 델피니딘의 3-OH에 연결되어 있다는 것을 나타낸다. 또한 글루코실 (δ 4.06; 3.59)의 H-6와 람노실(rhamnosyl) (δ 4.65)의 H-1'' 사이에 약한 negative NOE 가 나타나는 것은 1→6 결합(linkage)임을 나타낸다. 화합 물 2의 스펙트럼 데이터는 delphinidin-3-O-rutinoside의 문헌 상에 보이는 데이터와 일치하였다 (Due
òas et al., 2008).
On-line HPLC 에서 얻어진 화합물 5의 UV-Vis 스펙트럼은 529 ㎚에서 흡수극대파장을 보였다. 화합물 5의 NMR 스펙트 럼은 화합물 2의 스펙트럼과 많은 유사함을 보였다. 양성자 공 명을 해석한 결과 화합물 5는 한 개의 메톡시 그룹 (δ 3.92)과 두 개의 하이드록실 그룹을 가진 비대칭 안토시아니딘 B-고리 (asymmetric anthocyanidin B-ring) 로써 페츄니딘 (petunidin)과 일치하였다. HMBC 스펙트럼의 5.34/145.1 (H-1''/C-3)에서의 크로스 피크 (cross peak)로 글루코서크 아글리콘 (glucoserk aglycone) 의 3-OH에 결합해 있다는 것을 확인하였고 화합물 5 의
13C 스펙트럼에서 C-6'' (67.7 ppm)의 downfield shift는 3- glucose 와 rhamnose가 6''-OH에서 결합된다는 것을 나타낸다.
HMBC 스펙트럼의 4.70/67.7 (H-1''/C-6''), 4.10/102.2 (H-6"a/
C-1'"), 그리고 3.59/102.2 (H-6"b/C-1'")에서의 크로스 피크를 보면 rhamnose와 inner glucose 사이에 6"-OH에서 결합이 이루어짐을 알 수 있었고 따라서 화합물 5는 petunidin-3-O- rutinoside 로 확인되었다.
화합물 7의 [M
+] 는 화합물 5보다 14 mass unit가 더 컸다.
1
H-NMR 스펙트럼을 보면 B-고리에 두 개의 동등한 aromatic proton ( δ 7.88)과 두 개의 methoxy group (δ 3.98)이 있음 을 알 수 있고, 따라서 화합물 7의 algycone은 malvidin이며 당 잔기의
1H 와
13C-NMR 스펙트럼은 화합물 2, 5와 거의 동등했고,
1H-
1H COSY, HMBC, NOE 스펙트럼으로 확인하 였다. UV-VIS 스펙트럼은 0.5% HCl-MeOH 용액에서 282 ㎚와 533 ㎚일 때 λ max를 나타내었고, FAB-MS에서는 m/z 639 에서 [M
+] 를 보여주었다. 말비딘-3-글루코사이드 (m/z 493), 말비딘 (m/z 331)과 일치하는 프레그먼트를 가지고 C
29H
35O
16+의 분자구조식을 얻을 수 있었다. 따라서 화합물 7 은 malvidin-3-O-rutinoside임을 확인하였다.
상기한 결과를 종합하면, 맥문동 종실 추출물에서 분리한 화 합물 1은 델피니딘-3-글루코사이드 (delphinidin-3-glucoside), 화
합물 2는 델피니딘-3-루티노사이드 (delphinidin-3-rutinoside), 화 합물 5는 페츄니딘-3-루티노사이드 (petunidin-3-rutinoside), 화 학물 7은 말비딘-3-루티노사이드 (malvidin-3-rutinoside)로 동정 되었으며, 맥문동 종실을 구성하고 있는 안토시아닌은 맥문동 종실 추출물에서 delphinidin-3-O-rutinoside, cyanidin-3-O- glucoside, petunidin-3- O-glucoside, petunidin-3-O-rutinoside, malvidin-3- O-glucoside, malvidin-3-O-rutinoside 의 총 7개 안 토시아닌의 분리 동정하여 맥문동 열매를 구성하고 있는 안토 시아닌의 구성과 종류를 보고한 결과 (Lee and Choung, 2011) 와 유사하였으며, 종실에 포함되어져 있는 안토시아닌의 종류 는 모두 열매에 포함되어져 있는 결과를 나타내었다.
2. 맥문동 종실 안토시아닌 분획의 mushroom tyrosinase (in vitro) 및 DOPA auto-oxidation 저해활성
미백효능의 in vitro 계 효력시험으로 in vitro tyrosinase inhibition assay 와 in vitro DOPA oxidation inhibition assay 가 가장 많이 사용되고 있다. Tyrosinase은 구리 (copper)를 포함하는 polyphenol oxidase로서 미생물과 동물, 식물조직에 존재하여 멜라닌 합성과 색소 생성에 기여하는 monophenol의 hydroxylation 을 촉매하는 효소이다 (Chen and Kubo, 2002).
Tyrosinase 는 인체 내의 멜라닌 생합성 경로에서 가장 중요한 초기 속도결정단계에 관여하는 효소로서 많은 미백 성분이 이 효소를 억제 하는 작용기전을 가지고 있다. 이 시험은 시험관 내에서 타이로시나제 효소의 활성을 저해하는 정도를 평가하 는 방법이다 (Ishihara et al., 1991). 또한 DOPA oxidation assay 는 멜라닌 합성과정의 속도결정단계를 촉매하는 tyrosinase 의 DOPA 산화반응에 대한 활성저해를 측정하여 미백 성분의 효과를 평가하는 것이다 (Choi et al., 2001).
맥문동 종실 안토시아닌 분획을 대상으로 하여 mushroom
tyrosinase 저해 활성 (in vitro)과 B16 melanoma cell을 이용
한 tyrosinase 저해 활성 (ex vivo)을 검정하여 L-tyrosine과
경쟁적으로 작용하는 저해제이며 (Maeda and Fukuda, 1996)
미백제로 가장 많이 사용되는 알부틴과 비교하여 본 결과
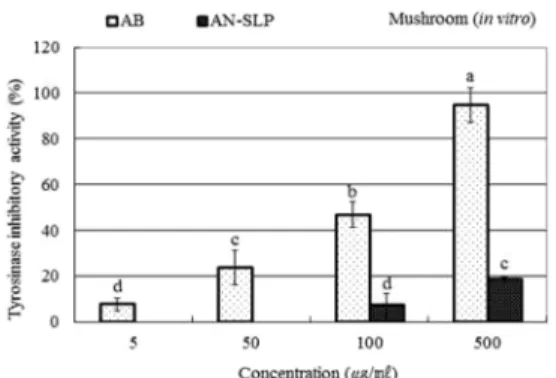
arbutin 을 5 ㎍/㎖ 처리 시 mushroom tyrosinase 저해 활성이
7.6% 로 나타나고 농도를 50, 100, 500 ㎍/㎖로 증가시켰을 때
각각 23.5%, 46.8%, 94.6%로 증가한 반면 맥문동 종실 안토
시아닌 분획물의 mushroom tyrosinase 저해 활성은 5에서
100 ㎍/㎖ 의 농도로 처리한 경우 활성이 검정되지 않았고
500 ㎍/㎖의 고농도에서 약 18.6%의 저해활성을 나타내어 비
교적 낮은 저해효과를 나타내었다 (Fig. 2). 또한 맥문동 종실
안토시아닌 분획물의 DOPA auto-oxidation 저해활성에서는 양
성대조군인 kojic acid가 200 ㎍/㎖의 농도에서 61.7%의 저해
활성을 나타낸 반면 맥문동 종실 안토시아닌 분획물은 동일한
농도에서 6.4%의 낮은 저해활성을 나타내었고 양성대조군인
kojic acid 가 400 ㎍/㎖의 농도에서 86.8%, 800 ㎍/㎖의 농도
에서 95.4%, 1000 ㎍/㎖ 에서 98.7%로 나타난 반면 맥문동 종실 추출물의 경우 1000 ㎍/㎖ 이상이 되어야 66.5%의 저해 활성을 나타내었다 (Fig. 3). 이러한 결과를 통하여 kojic acid 가 tyrosinase 활성 부위의 copper를 킬레이트화하여 tyrosine 에서 DOPA로 그리고 DOPA에서 DOPA quinone으로 진행되 는 과정을 저해하는 메카니즘을 가지나 (Battaini et al., 2000) 맥문동 종실의 안토시아닌의 경우 이러한 tyrosinase의 효소 활성 부위에 직접적으로 작용하여 효소활성을 억제하는 역할은 매우 적은 것으로 판단된다.
3. 맥문동 종실 안토시아닌 분획의 B16 melanoma cell에 대한 세포독성 검정
미백활성 성분의 멜라닌생성 억제력 시험은 기본적으로 대
상성분이 세포에 독성을 나타내거나 성장을 억제하지 않아야 한다는 것을 전제로 한다. 이유는 세포독성에 의한 멜라닌 생 성량 감소는 멜라닌생성 과정에 대한 저해에 해당되지 않기 때문이다. 따라서 본 실험에서는 맥문동 종실 안토시아닌 분 획물의 적용농도 5, 50, 100, 500 ㎍/㎖에서의 세포독성 여부 을 확인하고자 하였다. Figure 4에서 보는 바와 같이 맥문동 종실 추출물 500 ㎍/㎖ 이하의 농도에서는 세포의 성장에 영 향을 미치지 않아 세포독성이 없는 것을 확인하였다. 맥문동 종 실 안토시아닌 분획물 500 ㎍/㎖ 처리 시에는 약 6.8%의 세포 생장을 억제하였으나 통계적으로 유의한 수준은 아니었으며 기 존에 알려진 arbutin이나 kojic acid와 마찬가지로 세포의 생장 에 영향을 미치지 않아 세포 독성이 없는 것으로 확인되었다.
반면 hydroquinone의 경우 5 ㎍/㎖의 낮은 농도에서도 세포 성 장에 영향을 주는 세포독성이 있다는 것을 확인하였다 (Fig. 4).
4. B16 melanoma cell에서 맥문동 종실 안토시아닌 분획의 ex vivo tyrosinase 저해 활성 및 멜라닌 생성 억제 효과
맥문동 종실 안토시아닌 분획을 대상으로 B16 melanoma cell 을 이용한 ex vivo tyrosinase 저해 활성을 검정하여 본 결 과 맥문동 종실 안토시아닌 분획물을 5 ㎍/㎖의 수준으로 처 리하는 경우 ex vivo tyrosinase 저해 활성이 약 12.5%를 나 타내고 50, 100, 500 ㎍/㎖ 로 농도를 증가시켰을 때 각각 25.6%, 48.7%, 97.6% 의 저해활성을 나타내어 arbutin과 유사 하거나 약간 높은 활성을 나타내었다 (Fig. 5).
또한 맥문동 종실 안토시아닌 분획물의 세포수준에서의 멜 라닌 생성 억제효과를 검정하여 본 결과 일반적인 미백 물질 로 알려진 arbutin과 kojic acid의 경우, 50 ㎍/㎖의 농도에서 멜라닌 생성 억제율이 36.5%인 것에 반해, 맥문동 종실 안토 시아닌 분획물은 50 ㎍/㎖ 농도에서 53.2%로 나타나 arbutin
Fig. 2. The inhibitory effect on mushroom tyrosinase activity(in vitro) by anthocyanin rich fraction (AN-SLP) from L. platyphylla seed. Abbreviations are AB: arbutin and AN-SLP: anthocyanin rich fraction from L. platyphylla seed. The data shown represent mean± S. D. (n = 3) and the inhibitions were expressed as percentage. Mean with difference letter (a-d) within a column are significantly different at p < 0.05.
Fig. 3. Concentration-dependent inhibition of DOPA auto- oxidation activity by anthocyanin rich fraction (AN- SLP) from L. platyphylla seed. Abbreviations are KA;
kojic acid, AN-SLP; anthocyanin rich fraction from L.
platyphylla seed. The data shown represent mean ± S. D.
(n = 3) and the inhibitions were expressed as percentage.
Mean with difference letter (a-d) within a column are significantly different at p < 0.05.
Fig. 4. Effects of anthocyanin rich fraction (AN-SLP) from L.
platyphylla seed on cell viability of B16 melanoma cells. Cells were incubated with plant extracts for three days. Concentration of positive control agents such as kojic acid (KA) and arbutin (AB) were 50㎍/㎖.
Abbreviations are AN-SLP OOO; concentrations of anthocyanin rich fraction (AN-SLP) of L. platyphylla seed (㎍/㎖), HQ ; hydroquinone (5 ㎍/㎖), The data shown represent mean± S. D. (n = 3) and the absorbance was measured at 540 nm. Mean with difference letter (a and b) within a column are significantly different at p < 0.05.
과 kojic acid보다 약 0.7배 정도 더 높은 멜라닌 생성 저해 효과를 나타냄을 확인하였다 (Fig. 6). 또한 맥문동 종실 안토 시아닌 분획물의 농도를 달리하여 처리하는 경우, 멜라닌 생 성의 감소에 의해 세포의 색은 농도 의존적으로 흐려지는 것 을 관찰할 수 있었으며 IC
50값은 57.5 ㎍/㎖이었다 (Fig. 7).
상기 결과를 통하여 맥문동 종실 안토시아닌 분획물은 in vitro tyrosinase inhibition assay와 in vitro DOPA oxidation inhibition assay 에서 낮은 저해활성을 나타내나 B16 melanoma cell 을 이용한 ex vivo tyrosinase inhibition assay 와 멜라닌 생성 저해 평가에서 양성 대조구인 kojic acid나 arbutin 과 유사한 수준으로 높은 활성을 나타낸다는 것을 알
수 있었다. 이러한 것은 맥문동 종실 안토시아닌 분획물이 kojic acid 나 arbutin과 같이 tyrosinase의 효소활성에 직접적으 로 저해작용을 하여 멜라닌이 합성되는 되는 것을 저해하는 것이 아닌 melanoma 세포에 작용하여 tyrosinase의 활성을 억 제시키기 때문인 것으로 생각된다.
Chang 등 (2005)은 콩으로부터 분리한 genistein (IC
50= 0.822 mM) 의 mushroom tyrosinase 저해 활성이 kojic acid 에 비해 (IC
50= 0.054 mM) 수십 배가 낮아 활성이 있다고 보기 어렵다고 보고하였으나 이후 Yang 등 (2008)에 의하면 세포수준에서 genistein의 멜라닌 생성 저해 효과가 50 ㎛에서 55% 로 비교적 높게 나타남으로써 genistein이 tyrosinase에 직 접 작용하여 저해하지는 않지만 다른 경로로 멜라닌 생성을 저해할 것으로 추정하였다. 본 연구 결과에서는 이와 유사하 게 맥문동 종실 안토시아닌 분획물은 tyrosinase에 대하여 직 접적인 저해 효과는 적으나 세포수준에서 tyrosinase 활성을 농도 의존적으로 억제하였고 (Fig. 5.) 그 정도는 멜라닌 생성 억제율과 상관성이 매우 높았다는 결과를 얻었다 (Fig. 7). 이 런 결과로 보아 맥문동 종실 추출물은 tyrosinase의 발현의 전 사 단계에 관련하거나 혹은 tyrosinase 발현 및 효소활성화 이후 또는 단백질의 번역 후 변형 (post-translation modification) 단 계에 관여하여 미백 효과를 나타내는 것으로 추정해 볼 수 있 을 것으로 생각된다.
Fig. 5. The inhibitory effect on B16 melanoma cell tyrosinase activity (ex vivo) by anthocyanin rich fraction (AN- SLP) from L. platyphylla seed. Abbreviations are AB;
arbutin, AN-SLP; anthocyanin rich fraction from L.
platyphylla seed. The data shown represent mean ± S.
D. (n = 3) and the inhibitions were expressed as percentage. Mean with difference letter (a-e) within a column are significantly different at p < 0.05.
Fig. 6. Effects of anthocyanin rich fraction (AN-SLP) form L.
platyphylla seed on melanin synthesis of B16 melanoma cells. Cells (2× 105 cells/ well) were incubated with anthocyanin rich fraction (AN-SLP) for three days. Concentrations of AN-SLP and control agents such as kojic acid (KA) and arbutin (AB) were 50㎍/㎖.
Mean values± S. D. from triplicate separated experiments are shown. Mean with difference letter (a and b) within a column are significantly different at p < 0.05.
Abbreviations are AB; arbutin, KA; kojic acid, AN-SLP;
anthocyanin rich fraction from L. platyphylla seed.
Fig. 7. Images (A) and rates (B) of concentrations dependent inhibition of melanin synthesis in B16 melanoma cells by anthocyanin rich fraction (AN-SLP) from L.
platyphylla seed. Relative melanin synthesis value was expressed compared with control as 100%. Mean values± S. D. from triplicate separated experiments are shown. Mean with difference letter (a~d) within a column are significantly different at p < 0.05.
5. 맥문동 종실 안토시아닌 분획의 α-glucosidase 억제 효과 상기 결과로부터 맥문동 종실 안토시아닌 분획물은 tyrosinase 활성화 이후 단계에서 작용하여 melanoma 세포에 서 멜라닌 생성 억제효과를 나타내며 세포의 tyrosinase 활성 을 저해하는 것으로 나타났다. 다양한 미백소재 중에서도 glucosamine 과 tunicamycine는 tyrosinase를 직접 저해하지도 않고, 발현이나 분해와도 관련되지 않으면서 세포수준에서는 tyrosinase 활성을 저해하여 미백효과를 나타내는데 이러한 것 은 glucosamine과 tunicamycine는 tyrosinase의 post-translational modification 단계에서 작용하기 때문이라고 보고하고 있다 (Mishima and Imokawa, 1983). 맥문동 종실 안토시아닌 분 획물은 tyrosinase 발현 이후의 과정을 저해할 가능성이 높음 으로, α-glucosidase 저해제가 번역 후 활성화를 위한 tyrosinase 의 glucose trimming 과정을 저해함으로써 tyrosinase 의 glycosylation 반응을 억제하여 멜라닌 합성이 억제할 수 있음 (Branza-Nichita et al., 1999)을 감안하여 맥문동 종실 안토시아닌 분획물이 α-glucosidase의 활성 억제 효과가
있는지 확인 하였다.
맥문동 종실 안토시아닌 분획물의 농도별 α-glucosidase 저 해활성을 검정하여 대조구인 acarbose (Vedula et al., 1991) 와 비교하여 본 결과 시료를 50 ㎍/㎖의 수준으로 처리한 경 우 양성대조군인 acarbose가 7.8%의 저해율을 나타낸 반면 맥 문동 추출물은 2.3%의 저해율을 나타내었다가 100 ㎍/㎖로 처리되어진 경우에서는 acarbose 는 14.3%, 맥문동 종실 안토 시아닌 분획물은 이보다 높은 17.1%의 저해율을 나타내었다.
500 ㎍/㎖ 의 고농도로 처리되어진 경우 양성대조군 인 acarbose 는 71.4%의 저해율을 나타내었고 맥문동 종실 안토시 아닌 분획물은 이 보다 약한 33.9%의 α-glucosidase 저해활성 을 나타내어 농도의존적으로 저해활성이 증가하는 경향 을 나타내었다. 이러한 결과는 α-glucosidase 저해제인 DNJ (deoxynojirimycin) 와 α-mannosidase 의 저해제 인 deoxymannojirimycin (DMJ)이 tyrosinase의 glycosylation을 차단하여 멜라노좀으로의 이동을 억제하며 멜라닌 생성을 억제 할 수 있다고 보고한 결과 (Choi et al., 2007)와 유사하였으 며 제네스테인 (genestein)이 tyrosinase에 대한 저해와 멜라닌 생성에 대한 억제 효과가 제네스테인이 가지는 α-glucosidsase 의 저해활성에 기인한다는 보고와도 유사하였다 (Yang et al., 2008). Tyrosinase glycosylation 저해제에 대한 보고들에 의하 면 Imokawa 등 (1984)은 B16 melanoma 세포 배양에서 glucosamine 과 tunicamycin 같은 물질들은 tyrosinase의 glycosylation 반응을 저해하여 멜라닌 합성이 완전히 억제될 수 있음을 보고하였으며 Petrescu 등 (1997)에 의하면, ER processing enzyme 인 α-glucosidase I, II의 저해제인 NB- DNJ 가 B16 melanoma 세포의 활성을 억제하며 비처리군에 비해 2%의 멜라닌만이 합성되었다고 보고하여 tyrosinase glycosylation 저해제가 멜라닌 생성을 억제할 수 있음을 알 수 있다 (Imokawa et al., 1984; Petrescu et al., 1997). 그러므 로 delphinidin-3-glucoside, delphinidin-3-rutinoside, petunidin- 3-rutinoside, malvidin-3-rutinoside 의 안토시아닌을 포함하는 맥문동 종실 안토시아닌 분획물은 세포 수준에서 tyrosinase의
Fig. 8. Inhibitory activities of anthocyanin rich fraction (AN-SLP) from L. platyphylla seed against rat intestinal α- glucosidase. Abbreviations are AC; acarbose, AN-SLP ; anthocyanin rich fraction from L. platyphylla seed. Mean values± S. D. from triplicate separated experiments are shown. Mean with difference letter (a~d) within a column are significantly different at p < 0.05.
Table 1. Peak assignments and mass spectral data of anthocyanins in the seeds of L. platyphylla, together with cited references.
Peak number
Retention time
(min) λmax (㎚) MS
〔 M+〕 Chemical
formular Identity Reference
1 17.8 278, 526 465
303 C21H21O12+ Delphinidin-3-O-glucoside Cerezo et al., 2010
2 18.3 278, 530
611 303 465
C27H31O16+ Delphinidin-3-O-rutinoside Dueòas et al., 2008
5 20.8 278, 529
625 317 479
C28H33O16+ Petunidin-3-O-rutinoside Pazmiòo-Duran et al., 2001
7 23.6 282, 533
639 331 493
C29H35O16+ Malvidin-3-O-rutinoside Pazmiòo-Duran et al., 2001
활성을 억제하고 멜라닌 생성을 억제하는데 이것은 tyrosinase 효소에 대해 직접적인 저해 작용이나 발현을 억제하는 것이 아 닌 rat intestinal 기원의 α-glucosidase 활성을 농도 의존적으로 저해한 것으로 나타났으므로 맥문동 종실 안토시아닌 분획물은 tyrosinase 단백질의 번역 후 변형 (post-translation modification) 단계에서 glycosylation을 억제함에 따라 tyrosinase을 억제하여 멜라닌 생성을 억제하는 것으로 판단된다.
안토시아닌과 유사한 polyphenol류 화합물인 kampferol, quercetin 과 morin 등의 flavonoid가 tyrosinase 효소의 활성 부위 (active center)에 존재하는 copper에 대한 chelating 효 과에 의해 tyrosinase 저해 활성을 가진다는 보고 (Sánchez- Ferrer et al., 1995)와 연관하여 맥문동 종실로부터 획득한 안 토시아닌 분획은 500 ㎍/㎖ 이상의 고농도로 처리되는 경우 미약한 수준이나마 tyrosinase에 대하여 직접적인 저해활성과 L-DOPA oxidation 의 저해 활성을 나타낼 뿐만 아니라 실제 세포 수준에서는 피부세포에서의 tyrosinase 효소활성을 저해 하고 멜라닌의 생성을 arbutin 보다 더 높은 수준으로 저해하 는 결과를 나타내었고 맥문동 종실 안토시아닌 분획이 α- glucosidase 활성을 농도 의존적으로 저해함을 검정함으로써 세포에서 tyrosinase 효소의 활성을 위한 glycosylation을 억제 시킬 수 있 수 있고 tyrosinase 효소의 번역 후 변형을 통해 melanogenesis 를 방지할 수 있는 미백화장품으로서의 개발 가 능성이 매우 높다고 할 수 있겠다.
LITERATURE CITED
Alaluf S, Heath A, Carter N, Atkins D, Mahalingam H, Barrett K, Kolb R and Smit N. (2001). Variation in melanin content and composition in type V and VI photoexposed and photoprotected human skin: The dominant role of DHI.
Pigment Cell Research. 14:337-347.
Aramwit P, Nipaporn B and Srichana T. (2010). The properties and stability of anthocyanins in mulberry fruits. Food Research International. 43:1093-1097.
Battaini G, Monzani E, Casella L, Santagostini L and Pagliarin R. (2000). Inhibition of the catecholase activity of biomimetic dinuclear copper complexes by kojic acid. Journal of Biological Inorganic Chemistry. 5:262-268.
Branza-Nichita N, Negroiu G, Petrescu AJ, Garman EF, Platt FM, Wormald MR, Dwek RA and Petrescu SM. (2000).
Mutations at critical N-glycosylation sites reduce tyrosinase activity by altering folding and quality control. Journal of Biological Chemistry. 275:8169-8175.
Branza-Nichita N, Petrescu AJ, Dwek RA, Wormald MR, Platt FM and Petrescu SM. (1999). Tyrosinase folding and copper loading in vivo: A crucial role for calnexin and alpha-glucosidase II. Biochemical and Biophysical Research Communications.
261:720-725.
Cerezo AB, Cuevas E, Winterhalter P, Garcia-Parrilla MC and Troncoso AM. (2010). Isolation, identification, and antioxidant
activity of anthocyanin compounds in Camarosa strawberry.
Food Chemistry. 123:574-582.
Chang TS, Ding HY and Lin HC. (2005). Identifying 6,7,4’- trihydroxyisoflavone as a potent tyrosinase inhibitor. Bioscience, Biotechnology and Biochemistry. 69:1999-2001.
Chen QI and Kubo I. (2002). Kinetics of mushroom tyrosinase inhibition by quercetin. Journal of Agricultural and Food Chemistry. 50:4108-4112.
Choi H, Ahn S, Chang H, Cho NS, Joo K, Lee BG, Chang I and Hwang JS. (2007). Influence of N-glycan processing disruption on tyrosinase and melanin synthesis in HM3KO melanoma cells. Experimental Dermatology. 16: 110-117.
Choi SI, Park JH, Her YK, Lee YK, Kim JE, Nam SH, Goo JS, Jang MJ, Lee HS, Son HJ, Lee CY and Hwang DY.
(2010). Effects of water extract of Liriope platyphylla on the mRNA expression and protein secretion of nerve growth factors. Korean Journal of Medicinal Crop Science. 18:291-297.
Choi SS, Noh HS, Cho SH and Kong KH. (2001). Screening of inhibitors against tyrosinase activity from natural products.
Yakhak Hoeji. 45:522-528.
Dueòas M, Perz-Alonso JJ, Santos-Buelga C and Escribano- Bailn T. (2008). Anthocyanin composition in fig(Ficus caruca L.). Journal of Food Composition and Analysis. 21:107-115.
Gómez-Cordovés C, Bartolomé B, Vieira W and Virador VM.
(2001). Effects of wine phenolics and sorghum tannins on tyrosinase activity and growth of melanoma cells. Journal of Agricultural and Food Chemistry. 49:1620-1624.
Ha SK, Moon EJ, Lee MJ, Park HM, Yoo ES, Oh MS and Kim SY. (2009). Effect of the BuOH soluble fraction of Cinnamomum camphora on melanin biosynthesis. Korean Journal of Medicinal Crop Science. 17:293-300.
Hearing VJ. (1999). Biochemical control of melanogenesis and melanosomal organization. Journal of Investigative Dermatology Symposium Proceedings. 4:24-28.
Imokawa G and Mishima Y. (1984). Functional analysis of tyrosinase isozymes of cultured malignant melanoma cells during the recovery period following interrupted melanogenesis induced by glycosylation inhibitors. Journal of Investigative Dermatology. 83:196-201.
Ishihara Y, Oka M, Tsunakawa M, Tomita K, Hatori M, Yamamoto H, Kamei H, Miyaki T, Konishi M and Oki T.
(1991) Melanostatin, a new melanin synthesis inhibitor.
Production, isolation, chemical properties, structure and biological activity. Journal of Antibiotics. 44:25-32.
Jang DI, Lee BG, Jeon CO, Jo NS, Park SY, Lee H and Koh JS. (1997). Melanogenesis inhibitor from paper mulberry.
Cosmetics and Toiletries. 112:59-62.
Jeon KH, Lee BG, Park DG and Lee BS. (2001). Cosmetic composit for whitening containing Broussonetia kazinoki extract. Korea Patent. 10-0070824.
Jo YC, Kong YH, Lee YC, Kim SS and Choi SY. (2008).
Inhibitory effect of white ginseng fraction on skin pigmentation.
Korean Journal of Medicinal Crop Science. 16:192-194.
Joshi PC, Carraro C and Pathak MA. (1987). Involvement of reactive oxygen species in the oxidation of tyrosine and dopa to melanin and in skin tanning. Biochemical and Biophysical Research Communications. 142:265-274.
Katon W, Russo J, Lin EHB, Heckbert SR, Karter AJ, Williams LH, Ciechanowski P, Ludman E and Von Korff M. (2009).
Diabetes and poor disease control: Is comorbid depression associated with poor medication adherence or lack of treatment intensification? Psychosomatic Medicine. 71:965-972.
Kim EG and Kim DM. (2008). Extract of Nigella glandulifera Freyn et Sint having whitening activity and the cosmetic composition comprising thereof. Korea Patent. 10-0850686.
Kim JS, Kwon CS and Son KH. (2000). Inhibition of alpha- glucosidase and amylase by luteolin, a flavonoid. Bioscience, Biotechnology and Biochemistry. 64:2458-2461.
Kornfeld R and Kornfeld S. (1985). Assembly of asparagine linked oligosaccharides. Annual Review of Biochemistry.
54:631-664.
Krentz AJ, Patel MB and Bailey CJ. (2008). New drugs for type 2 diabetes mellitus: What is their place in therapy? Drugs.
68:2131-2162.
Lee JH and Choung MG. (2011). Identification and characterization of anthocyanins in the antioxidant activity- containing fraction of Liriope platyphylla fruits. Food Chemistry. 127:1686-1693.
Lee KM, Lee EC, Cho SC and Moon SS. (2008). The antimelanogenic effects of compounds extracted from bamboo inner film. Journal of the Society Cosmetic Scientists of Korea.
34:287-301.
Lee SH, Choi SY, Kim H, Hwang JS, Lee BG, Gao JJ and Kim SY. (2002). Mulberroside F isolated from the leaves of Morus alba inhibits melanin biosynthesis. Biological and Pharmaceutical Bulletin. 25:1045-1048.
Lowell AG. (1991). Physiology, biochemistry and molecular biology of the skin(2nd ed.) Oxford University Press Inc. New York, USA. p.891.
Lozovskaya T, Brenner-Weiss G, Franzreb M and Nusser M.
(2012). Recovery of anthocyanins from grape pomace extract(Pinot Noir) using magnetic particles based on poly(vinyl alcohol). Celluslose Chemistry and Technology. 46:427-433.
Maeda K and Fukuda M. (1996). Arbutin: Mechanism of its depigmenting action in human melanocyte culture. Journal of Pharmacology and Experimental Therapeutics. 276:765-769.
Matsui T, Ueda T, Oki T, Oshiro Sugita K, Terahara N and Matsumoto K. (2001).
α
-Glucosidase inhibitory action of natural acylated anthocyanins. 2.α
-Glucosidase inhibition by isolated acylated anthocyanins. Journal of Agricultural and Food Chemistry. 49:1952-1956.Mishima Y and Imokawa G. (1983). Selective aberration and pigment loss in melanosomes of malignant melanoma cells in vitro by glycosilation inhibitors: Premelanosomes as glycoprotein. Journal of Investigative Dermatology. 81:106-114.
Miyazawa M, Oshima T, Koshio K, Itsuzaki Y and Anzai J.
(2003). Tyrosinase inhibitor from black rice bran. Journal of Agricultural and Food Chemistry. 51:6953-6956.
Mosmann T. (1983). Rapid colorimetric assay for cellular growth and survival: Application to proliferation and cytotoxicity assays. Journal of Immunological Methods. 65:55-63.
Oki T, Matsui T and Osajima Y. (1999). Inhibitory effect of
α
- glucosidase inhibitors varies according to its origin. Journal of Agricultural and Food Chemistry. 47:550-553Pazmino-Durn EA, Giusti MM, Wrolstad RE and Glria MBA.
(2001). Anthocyanins from banana bracts(Musa X Paradisiaca) as potential food colorants. Food Chemistry. 73:327-332.
Petrescu SM, Petrescu AJ, Titu HN, Dwek RA and Platt FM (1997). Inhibition of N-glycan processing in B16 melanoma cells results in inactivation of tyrosinase but does not prevent its transport to the melanosome. Journal of Biological Chemistry. 272:15796-15803.
Roh JS, Han JY, Kim JH and Hwang JK. (2004). Inhibitory effects of active compounds isolated from safflower(Carthamus tinctorius L.) seeds for melanognensis. Biological and Pharmaceutical Bulletin. 27:1976-1978.
Sánchez-Ferrer A, Rodríguez-Lopez JN, García-Cánovas F and Garcia-Carmona F. (1995). Tyrosinase: A comprehensive review of its mechanism. Biochimica et Biophysica Acta.
1247:1-11.
Vanni A, Gastaldi D and Giunata G. (1990). Kinetic investigations on the double enzymatic activity of the tyrosinase mushroom. Annali Di Chimica-Rome 80:35-60.
Vedula U, Schnitzer-Polokoff R and Tulp OL. (1991). The effect of acarbose on the food intake, weight gain, and adiposity of LA/N-cp rats. Comparative Biochemistry and Physiology.
100:477-482.
Winchester B and Fleet GW. (1992). Amino-sugar glycosidase inhibitors: Versatile tools for glycobiologists. Glycobiology.
2:199-210.
Yang ES, Hwang JS, Choi HC, Hong RH and Kang SM.
(2008). The effect of genistein on melanin synthesis and in vivo whitening. Korean Journal of Microbiology and Biotechnology.
36:72-81.