A549 폐암 세포주에서 AMPK/Akt/mTOR 신호 경로를 통한 소나무 담쟁이덩굴 에탄올 추출물의 Apoptosis 유도효과
조경조1, 남건희1, 박예슬1, 김상용2, 구봉성3, 김영민1*
Apoptotic Effects of Parthenocissus tricuspidata (Siebold & Zucc.) Planch Ethanol Extract by regulating AMPK/Akt/mTOR Signaling Pathways in A549 Cancer Cells
Kyung Jo Jo1, Gun He Nam1, Ye Seul Park1, Sang Yong Kim2, Bong Seong Koo3, and Young Min Kim1*
Received: 29 July 2019 / Revised: 13 August 2019 / Accepted: 26 August 2019
© 2019 The Korean Society for Biotechnology and Bioengineering
Abstracts: Lung cancer can be caused by several environ- mental factors, such as smoking and urban and industrial pollution. Lung cancer also has a higher recurrence rate and a lower cure rate than other types of cancer. Parthenocissus tricuspidata (Siebold & Zucc.) Planch Extract (PTE) is known to have antioxidant, antidiabetic, and anti-inflammatory effects;
however, its efficacy as an anticancer agent has not been reported. The purpose of this study was to investigate the effect of PTE on apoptosis-related proteins in A549 lung cancer cells and to determine how apoptosis is induced through the AMP- activated protein kinase (AMPK)/Akt/mTOR signaling pathway.
AMPK is activated by metabolic stress, such as glucose deficiency, and induces apoptosis through the intrinsic pathway that activates tumor suppressor p53, thereby regulating the bcl- 2 family protein, releasing cytochrome c from the mitochondria,
and activating caspase. In addition, Akt plays an important role in the metabolism, growth, and survival of cancer cells. Inhibition of Akt induces apoptosis by regulating key signaling molecules, such as the mammalian target of rapamycin and tumor suppressor p53. In this study, we examined the effect of PTE on the apoptosis of A549 cells and determined how the regulation of apoptosis proteins through the intrinsic pathway is achieved.
Our results demonstrated that PTE regulates bcl-2 proteins and induces apoptosis through the intrinsic pathway. In addition, AMPK and Akt inhibitors were treated to determine whether these apoptotic effects were dependent on AMPK and Akt.
Keywords: A549, apoptosis, Parthenocissus tricuspidata (Sie- bold & Zucc.) Planch, anti-cancer, AMPK/Akt/mTOR path- way, Intrinsic pathway, Bcl-2 family
1. INTRODUCTION
암은 우리나라를 포함한 전 세계적으로 생명을 위협하는 가 장 중요한 요소로 지목되고 있으며, 정상 세포와는 다르게 무분별한 증식을 하여 형성된 악성종양을 말한다 [1]. 기존 의 암 치료 방법들은 치료 효과를 기대하기 전에 다양한 부 작용들을 일으킨다. 일반적으로 오심이나 구토, 식욕부진, 구내염, 탈모, 피로, 설사, 빈혈 등의 신체적 증상과 이에 따 라 불안이나 우울, 두려움, 절망감과 같은 심리적인 문제들
1한남대학교 생명나노대학 생명시스템과학과
1Department of Biological Sciences and Biotechnology, Hannam University, Daejeon 34054, Korea
Tel: +82-42-629-8753, Fax: +82-42-629-8873 e-mail: [email protected]
2신안산대학교 식품생명과학과
2Department of Food Science & Technology, Shinansan University.
Ansan 15435, Korea
3포바이오코리아 기술개발연구소
3Research Center, ForBioKorea Co., Ltd., Seoul 08592, Korea
Research Paper
을 야기하는데 이러한 신체적 심리적 문제들로 인해 암 환자 들의 삶의 질이 낮아진다 [2].
따라서 암세포에만 작용하여 정상 세포에는 영향을 주지 않는 천연물질을 이용한 항암치료제를 개발하려는 연구가 활발히 진행되고 있으며, 이러한 천연물질을 이용하여 암세 포 주에서 세포 신호 분자 조절을 통한 apoptosis 유도 효과 에 관한 연구가 진행되고 있다 [3,4].
apoptosis는 프로그램된 세포 죽음으로서 DNA의 손상이나 산화적 스트레스, 대사 스트레스 등에 의하여 세포가 더는 생존할 수 없을 때 발생하며, 세포 수축, 핵의 응축 및 세포막 기포 형성의 특징을 갖는다 [5,6]. 또한 급성 신경 손상, 신경 퇴행성 질환 및 암을 포함하는 다양한 병리학적 상태에 관련 된다. 따라서 이를 이용한 새로운 치료 전략을 설계하는데 중요할 수 있다 [7]. apoptosis는 death receptor 의존적 과정인 extrinsic pathway와 미토콘드리아 의존적 과정인 intrinsic pathway로 나누어진다 [8]. 내인성 경로는 미토콘드리아의 기 능 이상으로 미토콘드리아 막 투과성(mitochondrial membrane permeability, MMP)의 변화가 유발되어 미토콘드리아 내의 cytochrome C와 같은 apoptosis 유도 분자들이 세포질로 방 출됨으로써 apoptosis가 일어나는 것이다 [9]. 특히 내인성 경 로는 Bcl-2 family에 의해서 주로 조절되는데, 이들 단백질은 미토콘드리아 막 투과성의 변화를 유발하는 데 중요한 역할 을 하는 것으로 알려져 있다. 이러한 Bcl-2 family 중에서 apoptosis 억제 유전자인 Bcl-2와 이를 유도하는 유전자인 Bax 가 존재하여 길항적으로 조절된다고 알려져 있다 [10,11].
Protein kinase B (PKB/Akt)는 암세포의 대사와 성장, 그리고 생존에 중요한 역할을 하며, mammalian target of rapamycin (mTOR)나 p53과 같은 중요 신호 분자들을 조절한다 [12]. 또 한 AMPK의 활성은 p53을 활성화하여 내인성 경로를 통한 apoptosis를 유도한다 [13]. 소나무 담쟁이덩굴(Parthenocissus tricuspidata (Siebold & Zucc.) Planch)은 포도과 담쟁이덩굴 속의 낙엽 활엽만 목으로 한국, 중국, 일본, 대만 등지에 분포 하며 한국의 경우 전국의 표고 100-1,600m에서 자생한다. 소 나무 담쟁이덩굴은 항산화, 항당뇨 및 항염증 효과가 있는 것으로 알려져 있지만 항암효과로서의 효능은 아직 보고되 지 않았다. 이에 따라 본 연구에서는 소나무 담쟁이덩굴의 추출물 (PTE)이 A549 폐암 세포에서 apoptosis와 관련된 단 백질 발현에 미치는 영향과 apoptosis 유도 효과를 확인하고 자 하였으며, AMPK/Akt/mTOR 신호전달 경로를 확인하여 PTE 에 의한 apoptosis 효과가 어떠한 신호 경로를 통하여 이 루어지는지 제시하고자 한다.
2. MATERIALS AND METHOD
2.1 실험재료
본 실험에서 사용된 소나무 담쟁이덩굴의 원산지는 경상남 도 산청군이다. 소나무 담쟁이덩굴은 ㈜지리산 한방식품에 서 구매하여 분쇄 후 100 g에 95% Ethanol 800 mL을 가하여
72시간 동안 상온에서 환류 추출하였다. 추출된 소나무 담쟁 이덩굴은 감압 농축기 (N-1200A, Tokyo Rikakikai Co., Tokyo, Japan)를 이용하여 감압 농축시키고 농축된 추출물은 실험에 사용하기 전까지 –86°C에서 보관하였다. 이렇게 농 축된 추출물은 dimethyl sulfoxide (DMSO)를 이용하여 200 mg/mL stock으로 만들어 –20°C에 보관하여 사용하였다. 3- (4,5-Dimethylthiazol-2-yl)-2, 5-diphenyltetrazolium bromide (MTT) 는 Sigma-Aldrich (Sigma-Aldrich, USA)에서 구입하여 5 mg/
mL 농도로 만들어서 사용하였다. 또한, LY294002 (Sigma- Aldrich, USA)와 Compound C (Merck Millipore, USA)는 DMSO에 용해시켜 각각 20 mM과 10 mM로 만들어서 사용 하였다.
2.2 세포배양
본 연구에서 사용한 A549 폐암 세포는 American Type Culture Collection (ATCC, Rockville, MD, USA)에서 분양받 았으며, 10% FBS (HyClone Laboratories Inc., Logan, UT, USA)와 1% antibiotics (HyClone Laboratories Inc.)가 포함된 RPMI 1640 media (HyClone Laboratories Inc.)를 사용하였다.
Fibroblast 세포는 American Type Culture Collection (ATCC, Rockville, MD, USA)에서 분양 받았으며, 10% FBS (HyClone Laboratories Inc, Logan, UT, USA) 와 1% antibiotics (HyClone Laboratories Inc.)가 포함된 DMEM media (HyClone Laboratories Inc.)를 사용하여 5% CO
2, 37°C 조건으로 배양하였다. 48시 간마다 Trypsin-EDTA(HyClone Laboratories Inc.)를 이용하 여 세포를 부유 상태로 만든 다음 1×10
4cells/mL로 분주하고 계대배양하였다.
2.3 MTT assay
A549 세포와 fibroblast 세포를 12 well plate에 1×10
4cells/mL 로 seeding 한 후 5% CO
2, 37
oC 조건하에 incubator에서 24시 간 배양하였다. 그 후 PTE를 농도별 (25, 50, 75, 100, 125, 150 µg/mL)로 처리하고 5% CO
2, 37 조건하에 incubator에 24시간, 48시간 배양한 후, MTT solution을 well에 40 µL씩 처리 후 50분 동안 incubator에 배양하였다. PBS를 well에 1 mL 분주하여 washing 후, DMSO를 well에 150 µL씩 넣어 주었다. 96 well plate에 100 µL씩 취해 옮겨준 다음 FLUOstar Omega (BMG labtech, Germany)를 이용하여 595 nm 파장에서 흡광도를 측정하였다.
2.4 Observation of A549 cell morphological change 6 well plate 에 A549 세포를 well에 1×10
4cells/mL 로 분주하 여 24시간 동안 배양한 후 PTE를 농도별 (50, 75, 100 µg/mL) 로 처리하였다. 24시간 동안 incubator에서 배양한 후 A549 세포에서 일어난 형태 변화를 Inverted microscope (Zeiss, Oberkochen, Germany)로 관찰하였다.
2.5 Flow cytometry
Apoptosis를 정량적으로 분석하기 위해 Muse
TMAnnexin V
& Dead Cell Kit (Merck Millipore, USA)를 사용하였다.
A549 cell을 6 well plate에 1x10
5cells/mL로 seeding 후, 5%
CO
2, 37
oC 조건하에 incubator에서 24시간 배양하였다. 그 후 PTE 를 농도별 (50, 75, 100 µL/mL)로 처리해준 후 5% CO
2, 37
oC 조건하에 incubator에서 24시간 배양하였다. 그 후 Trypsin- EDTA(HyClone Laboratories Inc.)를 이용하여 세포를 부유 시킨 후 원심분리하여 모아주었다. 모아준 세포는 PBS를 이 용하여 washing 후 Muse
TMAnnexin V & Dead Cell Kit (Merck Millipore, USA) 를 처리하여 20분 반응시켰다. 반응 후 PB4455ENEU Muse
TMCell Analyzer (Merck Millipore, USA) 를 사용하여 분석하였다.
2.6 Western blotting
6 well plate 에 A549 세포를 well에 1×10
4cells/mL 로 분주하 여 5% CO
2, 37
oC 조건하에 incubator에서 24시간 배양하였다.
그 후 PTE를 농도별 (50, 75, 100 µg/mL)로 처리한 후 5%
CO
2, 37
oC 조건하에 incubator에서 24시간 배양하였다.
LY294002 (20 mM)과 Compound C (10 mM)는 PTE를 처리 하기 1시간 전에 처리하였다. 그 후 세포에 RIPA lysis buffer [25 mM Tris-Cl(pH 7.4), 1% NP40, 0.5% sodium deoxycholate, 150 mM NaCl, 1 mM PMSF]를 well에 150 µL씩 첨가하고 scrapper 를 이용하여 단백질을 분리하였다. 그런 다음 14,000 rpm, 4
oC에서 20분 동안 원심분리하여 상등액을 취하였다.
얻어진 상등액은 FLUOstar Omega (BMG labtech, Germany) 를 이용하여 595 nm에서 흡광도를 측정하여 단백질을 정량 하였다. 전기영동은 8%, 12% acrylamide gel을 이용하여 실 시하였고 nitro-cellulose membrane에 transfer 하였다. 그런 다음 4% Bovine serum albumin (BSA)을 이용하여 상온에서 1시간 30분 동안 blocking 하였다. 1차 항체는 4
oC에서 over night 하여 단백질에 결합시켰다. 다음날 2차 항체를 1-2시간 동안 반응 시킨 다음 Blue X-ray film에 감광하여 실험 결과 를 측정하였다.
2.7 통계처리
통계 프로그램인 SPSS (Statistical Package for the Social Science) 22.0 프로그램을 사용하였고 실험 설계에 대한 분산 분석은 독립 표본 t-test(Independent sample t-test)를 통하여 검정하였다. 각 자료는 최소 3번 이상의 반복된 실험을 통하 여 얻어진 결과로 검정하였고 p<0.05인 경우 통계적으로 유 의한 것으로 판정하였다.
3. RESULTS AND DISCUSSION
3.1 소나무 담쟁이덩굴 추출물이 A549 폐암 세포의 증식에 미치는 효과 확인
Apoptosis 는 생명체의 정상적인 발달과 분화에 관여하며 DNA의 손상이나 산화적, 대사적 스트레스로 인해 세포가 더 이상 생존할 수 없을 때 비정상적인 세포들을 제거하는 프로그
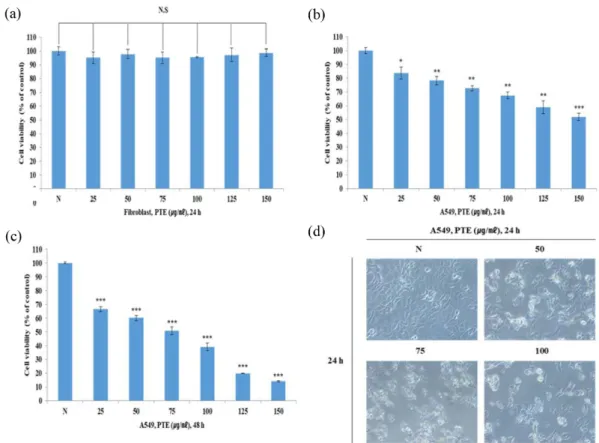
램된 세포의 죽음이다 [5,14]. 또한 정상 세포는 핵이나 세포질 의 형태가 일정하고 세포의 이동과 관련된 lamellipoia와 filopodia가 잘 관찰되지만, 세포사멸 시에는 세포 골격 단백 질의 변화로 인하여 세포질이 응축되고 핵이 분절되면서, lamellipodia와 filopodia의 소실과 같은 특이적인 형태학적 변화가 관찰된다 [15]. 본 연구에서는 PTE가 A549 폐암 세포 에 특이적으로 세포독성을 나타내는지 확인하기 위해 PTE 를 농도별 (25, 50, 75, 100, 125, 150 µg/mL)로 처리하여 MTT assay 를 통해 세포독성을 측정하였다. 그 결과 정상세 포인 Fibroblast 세포에서는 control 대비 모든 농도 군에서 생 존율이 90% 이상으로 유지되며 유의하지 않은 것으로 보아 PTE 가 정상세포에서 독성이 없음을 확인했다 (Fig. 1(a)). 반 면 A549 폐암 세포주에 PTE를 처리했을 때에는 control 대비 농도와 시간 의존적으로 세포의 증식이 유의하게 억제되는 것을 확인하였다 (Fig. 1(b), 1(c)). 또한 A549 세포에 PTE를 농도별(50, 75, 100 µg/mL)로 처리하였을 때 밀도가 높고 균 일하며 부착이 잘 되어있는 정상적인 A549 세포의 형태를 보이는 control 군에 비해 농도 의존적으로 세포의 밀도가 현 저히 감소하며 세포사멸에 있어 세포의 응축 같은 형태학적 인 변화가 일어난 것을 확인하였다 (Fig. 1(d)). 결과적으로 PTE는 정상 섬유아세포인 Fibroblast의 증식에는 영향을 미 치지 않으며 A549 폐암 세포주에 있어 농도와 시간 의존적 으로 세포의 증식을 특이적으로 억제하는 것을 확인하였다.
이러한 세포증식 억제효과가 Apoptosis에 의한 것이지 연구하 기 위해 위 결과를 기반으로 주요 농도를 50, 75, 100 µg/mL로 결정하여 실험에 활용하였다.
3.2 소나무 담쟁이덩굴 추출물에 의한 A549 폐암 세포의 apoptosis 유도 효과 확인
프로그램된 세포의 죽음인 apoptosis는 인지질 이중층 안쪽 에 존재하던 phosphatidylserine(ps)이 표면으로 노출되는 원형질막 구조의 변화가 동반되며 시간적 진행에 따라 phospatidylserine의 translocation이 발생은 하지만 세포막의 손상은 일어나지 않은 early apoptosis와 phospatidylserine의 translocation과 함께 세포막의 손상이 함께 일어나는 late apoptosis로 나누어진다 [16,17]. 앞서 PTE 처리에 의한 A549 세포의 증식 억제 효과에 대해 알아보았는데, 이러한 증식 억제 효과가 necrosis에 의한 것인지 apoptosis 유도에 의한 것인지를 확인하기 위해 Muse
TMAnnexin V & Dead Cell Kit 로 염색 후 flow cytometry을 실시하였다. 그 결과 PTE를 농 도별(50, 75, 100 µg/mL)로 처리한 후 24시간 동안 반응시켰 을 때 control군의 Apoptosis 유발 빈도는 약 9.6%로 매우 낮 았으나 소나무담쟁이 덩굴 추출물을 농도별 (50, 75, 100 µg/
mL)로 처리한 군의 경우 각각 Apoptosis 유발 빈도가 46.65%, 59.2%, 74.8% 로 소나무담쟁이 덩굴 추출물의 농도 가 증가할수록 apoptosis가 많이 유도된 것을 확인했다 (Fig.
2). 이러한 결과는 PTE 처리로 인한 A549 폐암 세포 특이적
인 생존율 및 증식 억제는 necrosis에 의한 것이 아니라
apoptosis에 기인한 결과였음을 시사한다.
3.3 소나무 담쟁이덩굴 추출물에 의한 intrinsic pathway 관련 apoptosis 단백질 발현 확인
Akt 는 암세포의 대사와 성장, 그리고 생존에 중요한 역할을
하고, mammalian target of rapamycin (mTOR)나 종양 억제인 자인 p53과 같은 중요 신호 분자들을 조절한다 [12,18]. 또한 AMPK 의 활성화는 mTOR의 발현을 억제시키며 이러한 Fig. 1. Anti-proliferation and morphological change effects of PTE in A549 cells. (a) Fibroblast cells were treated with variable concentrations of PTE (25~150 μg/mL) for 24 h. Cell viability was measured by MTT assay. (b) A549 cells were treated with variable concentrations of PTE (25~150 μg/mL) for 24 h. Cell viability was measured by MTT assay. (c) A549 cells were treated with variable concentrations of PTE (25~150 μg/mL) for 48 h. Cell viability was measured by MTT assay. (d) The morphology of A549 cells, Cells were treated with PTE (50, 75, 100μg/mL) for 24 h. Original magnification x200.Each figure is representative of an experiment that was repeated at least three times. The statistical analysis of the data was carried out by use of an independent sample t-test.
*p<0.05, **p<0.01 and ***p<0.001, NS is not significant (each experiment, n=3).
Fig. 2. PTE treatment induces apoptotic effect in A549 cells. (a) A549 Cells were treated with PTE (50, 75, 100 μg/mL) for 24 h.
apoptotic effect was measured by flow cytometry. (b) The graph of cells ratio at each groups.
AMPK의 활성화와 mTOR의 하향조절은 종양 억제 단백질 인 p53을 활성화하여 내인성 경로를 통한 apoptosis를 유도 한다 [13]. 이러한 상위 단백질들의 조절은 미토콘드리아에 서 세포질로 cytochrome c가 방출되어 이루어지는 intrinsic pathway에서 중요한 Bcl-2 family 단백질들을 조절한다 [19].
Bcl-2 family 단백질은 기능적으로 구분되는데 anti-apoptotic 단백질인 Bcl-2, Bcl-XL 등과 pro-apoptotic 단백질인 Bax가 heterodimer를 형성하거나 미토콘드리아의 기능을 조절한다 [20,21]. 또한 Bax, Bak, Bad 같은 pro-apoptotic 단백질들은 미토콘드리아로 이동되어 cytochrome c를 방출 할 수 있는 pore를 형성한다 [22]. 이렇게 미토콘드리아에서 apoptosis 과정이 활성화됨에 따라 caspase-3가 활성화되면서 DNA repair나 유전자 안정성을 유지시키는 PARP 단백질을 절단 하여 PARP 단백질이 역할을 할 수 없는 cleaved-PARP가 생 성되고 그 결과 특징적인 apoptosis의 형태를 생성한다 [23].
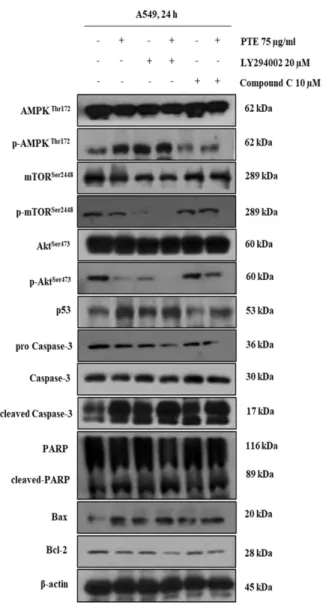
앞서 소나무 담쟁이덩굴 추출물이 apoptosis 룰 유도하는 것 을 확인하였고, 이러한 apoptosis가 intrinsic pathway 관련 단 백질을 어떻게 조절하여 유도되는지 알아보기 위해 상위 조 절 단백질인 Akt, AMPK, mTOR, p53 단백질뿐만 아니라 이 들로 인해 영향을 받는 하위 조절단백질의 발현 정도를 확인 하기 위해 A549 폐암 세포주에 소나무 담쟁이덩굴 추출물을 농도별 (50, 75, 100 µg/mL)로 처리한 후 24시간 반응시킨 후 western blotting을 실시하였다. 그 결과 암세포의 증식에 관 여하는 단백질들인 p-Akt, p-mTOR의 감소를 확인할 수 있으 며 p-AMPK는 증가하는 것을 볼 수 있다. 또한 p-AMPK의 증가는 종양 억제 단백질인 p53의 증가로 나타났다. 이러한 상위 단백질들의 조절을 받아 Bcl-2 family 단백질 중 anti- apoptotic 단백질인 bcl-2단백질은 감소하고 pro-apoptotic 단 백질인 Bax는 증가하는 것을 볼 수 있다. 또한 pro-caspase3 의 감소를 볼 수 있었으며 cleaved-caspase3의 증가를 볼 수 있었다. 이러한 경향은 PARP 단백질의 절단을 이끌어내어 cleaved-PARP 의 증가로 나타났다 (Fig. 3). 이러한 결과를 바 탕으로 PTE가 A549 폐암 세포에서 apoptosis 관련 단백질들 의 발현을 조절하여 intrinsic pathway를 통해 apoptosis를 유 도하는 것을 확인했다.
3.4 Akt, AMPK 저해제가 A549 폐암 세포 증식과 apoptosis 조절단백질 발현에 미치는 영향
본 실험에서는 PTE를 농도별 (50, 75, 100 µg/mL)로 처리한 후 실시한 western blot의 결과를 기반으로 소나무 담쟁이덩 굴 추출물이 Akt와 AMPK를 경유하는 signaling pathway를 통 해 apoptosis를 유도한다는 가설을 세우게 되었다. apoptosis가 일어나는 일련의 신호 경로를 확인하기 위해 각각 상위 단백 질인 Akt와 AMPK의 활성을 억제하는 ly294002와 Compound C 를 A549 세포에 처리하여 MTT assy와 western blotting을 시행하였다. 그 결과 MTT assay에서는 control 군과 비교했 을 때 PTE (75 µg/mL) 단독 처리 군의 경우 72.8% 정도의 생 존율을 보였으며 ly294002를 단독처리한 군은 세포 생장과 증식에 관여하는 Akt의 활성이 억제되어 74.3% 정도의 생존
율을 보였다. 또한 PTE와 ly294002를 병행 처리한 군의 경우 PTE나 ly294002를 단독 처리한 군보다 더 적은 59.4%의 생 존율을 보였다. 또한 control 군과 비교하여 AMPK 저해제인 Compound C를 처리한 군의 경우 96.1%의 생존율을 보여 control 군과 유의한 차이가 없었으며, PTE와 Compound C를 병행 처리한 군의 경우 PTE만을 단독 처리한 군보다 높은 84%의 생존율을 보였다 (Fig. 4). 이 결과는 AKT의 저해가 A549 폐암 세포의 증식 억제에 있어 중요한 역할을 하며 AMPK의 저해가 A549 폐암 세포의 사멸을 억제하는 것을 나타낸다. 다음으로 Akt, AMPK 저해제와 PTE(75 µg/mL) Fig. 3. PTE treatment regulates the expression of intrinsic pathway–
related apoptosis proteins in A549 cells. PTE effects on (p)mTOR,
(p)Akt, (p)AMPK, p53, pro caspase-3, cleaved caspase-3, Bcl-2,
cleaved-PARP, Bax and β-actin. Cells were treated with PTE (50,
75, 100 μg/mL) for 24 h. Protein levels were determined by
Western blotting. The β-actin probe served as protein -loading
control. Each protein is representative of an experiment that was
repeated at least three times
처리에 따른 apoptosis 단백질의 발현을 보면 p-Akt와 p- mTOR는 control 군 대비 PTE나 Akt 저해제를 단독처리한 군 의 경우 발현이 감소함을 볼 수 있었으며 병행 처리하였을 때 더 많은 감소를 보였다. 또한 AMPK 저해제를 단독처리 한 군의 경우 control 군과 비슷한 발현을 보였으며 AMPK 저해제와 물질을 병행 처리하였을 때에는 약간의 감소 경향 을 보였다. p-AMPK와 p53의 경우에는 control 군 대비 PTE 나 Akt 저해제를 단독처리한 군의 경우 발현이 증가함을 볼 수 있었으며 Akt 저해제와 물질을 병행 처리하였을 때에는 더 많은 증가를 보였다. AMPK 저해제를 단독으로 처리한 군에서는 control 군과 비슷한 경향을 보였고 AMPK 저해제 와 물질을 병행 처리 하였을 때에는 조금 증가하는 경향을 확인했다. 이러한 상위 단백질들의 영향을 받아 하위 단백질 인 Bax, cleaved-caspase3, cleaved-PARP는 control 군 대비 PTE나 Akt 저해제를 단독으로 처리한 군의 경우 발현이 증 가하는 것을 볼 수 있었으며 병행 처리하였을 때 단독처리 군들에 비해 더 많이 발현이 증가하는 것을 볼 수 있었다.
AMPK 저해제를 단독으로 처리하였을 때에는 control 군과 유사한 단백질의 발현을 보였고 PTE와 AMPK 저해제를 병 행 처리하였을 때에는 발현이 약간 증가하는 것을 볼 수 있 었다. 또한 Bcl-2와 pro-caspase3의 경우에는 control 군 대비 PTE나 Akt 저해제를 단독으로 처리한 군의 경우 발현이 감 소하는 것을 볼 수 있었으며 병행 처리하였을 때에는 단독 처리했을 때 보다 더 많은 발현의 감소를 볼 수 있었다. 또한 AMPK 저해제를 단독으로 처리하였을 때에는 control 군과 비슷한 발현을 보였고 AMPK 저해제와 물질을 병행 처리하
였을 때에는 발현이 약간 감소하는 것을 확인했다 (Fig. 5).
이와 같은 결과를 기반으로 PTE는 Akt를 억제하고 AMPK를 활성화시켜서 이를 경유하는 signaling pathway를 통해 apoptosis를 유도한다는 가설을 증명하였다.
4. CONCLUSION
본 연구에서는 PTE를 A549 폐암 세포에 처리함으로 인해 나 타나는 항암 효과가 Intrinsic pathway를 통한 Apoptosis 임을 Fig. 4. PTE, LY294002, and compound C affect the proliferation
of A549 cells. A549 Cells were pre-treated with 20μM LY294002 or 10μM compound C for 30 min and co-treated with 75μg/mL PTE for 24 h. Cell viability was measured by MTT assay. This figure is representative of an experiment that was repeated at least three times. The statistical analysis of the data was carried out by use of an independent sample t-test.
*p<0.05, **p<0.01 and ***p<0.001, NS is not significant (each experiment, n=3).
Fig. 5. PTE induces apoptosis through the Akt- and AMPK-
dependent pathway. PTE, LY294002 and compound C effects on
(p)mTOR, (p)Akt, (p)AMPK, p53, procaspase-3, cleaved caspase-
3, Bcl-2, cleaved-PARP, Bax and β-actin. A549 Cells were pre-
treated with 20 μM LY294002 or 10 μM compound C for 30 min
and co-treated with 75 μg/mL PTE for 24 h. Protein levels were
determined by Western blotting. The β-actin probe served as
protein-loading control. Each protein is representative of an
experiment that was repeated at least three times.
확인하였다. A549 폐암 세포에 PTE를 농도별 (25, 50, 75, 100, 125, 150 µg/mL)로 처리하였을 때 농도와 시간 의존적 으로 증식률이 감소함을 확인하였으며 정상 섬유아세포인 Fibroblast 에 PTE를 농도별 (25, 50, 75, 100, 125, 150 µg/mL) 로 처리하였을 때에는 control 군 대비 세포증식률에 영향을 주지 않는 것을 확인했다. 이는 PTE가 A549 폐암 세포에 특 이적으로 영향을 주는 것을 나타낸다. 또한 이러한 세포 증 식률의 감소는 Necrosis에 의한 것이 아니라 Apoptosis에 의한 것임을 Flow cytometry를 통해 확인하였다. 이러한 Apoptosis가 어떠한 단백질을 조절하여 일어나는지 확인하기 위해 PTE 를 농도별 (50, 75, 100 µg/mL)로 처리하여 Western blotting 을 실시한 결과 AMPK/Akt/mTOR 연관 단백질들을 농도 의 존적으로 조절함을 확인했다. 마지막으로 Akt와 AMPK 저 해제인 ly294002와 Compound C를 처리하여 세포의 증식률 과 AMPK/Akt/mTOR 연관 단백질들의 발현을 확인한 결과 PTE가 Akt를 억제하고 AMPK를 활성화시킴으로 인해 이를 경유하는 단백질들을 조절함으로써 Apoptosis를 유도하는 것을 확인하였다. 이러한 결과를 바탕으로 소나무 담쟁이덩 굴 추출물은 폐암 세포에 대해 새로운 치료법을 열어 줄 것 으로 기대되며, 치료물질로의 사용 가능성을 확인하기 위해 서는 다양한 추가 연구가 수행되어야 할 것이다.
Acknowledgements
본 연구는 2019년도 (주)포바이오의 용역 사업에 의해 수행 되었음.
REFERENCES
1. Choi, J. H., H. I. Kim, and I. S. Lee (2009) Effect of Rosmarinus officinalis L. on growth inhibition and apoptosis induction in can- cer cells. J. Korean Soc. Food Sci. Nutr. 38: 1008-1015.
2. Cleeland, C. S., T. R. Mendoza, X. S. Wang, C. Chou, M. T. Harle, et al. (2000) Assessing symptom distress in cancer patients: The MD anderson symptom inventory. Cancer: Interdisciplinary Inter.
J. Am. Cancer Soc. 89: 1634-1646.
3. Park, Y. J., M. H. Kim, and S. J. Bae (2002) Anticarcinogenic effects of Allium tuberosum on human cancer cells. Korean J.
Food Sci. Tech. 34: 688-693.
4. Lee, S. H., G. T. Kim, J. I. Kim, E. G. Lim, I. S. Kim, and Y. M.
Kim (2013) The Extract from Lysimachia foenum-graecum induces apoptosis in MCF-7 breast cancer cells. KSBB J. 28: 303- 309.
5. Park, C., C. Y. Jin, T. H. Choi, S. H. Hong, and Y. H. Choi (2013) Effect of proapoptotic Bcl-2 on naringenin-induced apoptosis in human leukemia U937 cells. J. Life Sci. 23: 1118-1125.
6. Kerr, J. F., A. H. Wyllie, and A. R. Currie (1972) Apoptosis: a basic biological phenomenon with wideranging implications in tis- sue kinetics. Br. J. Cancer 6: 239-257.
7. Zimmermann, K. C., C. Bonzon, and D. R. Green (2001) The
machinery of programmed cell death. Pharm.Ther. 92: 57-70.
8. Lim, E. G., G. T. Kim, B. M. Kim, E. J. Kim, S. H. Ha, et al.
(2016) Apoptotic effects and cell cycle arrest effects of extracts from Cnidium monnieri (L.) Cusson through regulating Akt/
mTOR/GSK-3β signaling pathways in HCT116 colon cancer cells. J. Life Sci. 26: 663-672.
9. Schuler, M., E. Bossy-Wetzel, J. C. Goldstein, P. Fitzgerald, and D.
R. Green (2000) p53 induces apoptosis by caspase activation through mitochondrial cytochrome c release. J. Biol. Chem. 275:
7337-7342.
10. Lee, J. W., Y. H. Soung, S. W. Nam, J. Y. Lee, N. J. Yoo, and S. H.
Lee (2006) Mutational analysis of pro-apoptotic BAD gene in non-small cell lung cancer. J. Lung Cancer 5: 35-38.
11. Jeong, Y. J. and K. J. Kang (2011) Effect of Angelica keiskei extract on apoptosis of MDA-MB-231 human breast cancer cells. J.Korean Soc. Food Sci. Nutr. 40: 1654-1661.
12. Kitagishi, Y., M. Kobayashi, K. Kikuta, and S. Matsuda (2012) Roles of PI3K/AKT/GSK3/mTOR pathway in cell signaling of mental illnesses. Depress. Res. Treat. 2012: 752563, DOI: 10.1155/
2012/752563.
13. Jing, K., K. S. Song, S. Shin, N. Kim, S. Jeong, et al. (2011) Doco- sahexaenoic acid induces autophagy through p53/AMPK/mTOR signaling and promotes apoptosis in human cancer cells harboring wild-type p53. Autophagy 7: 1348-1358.
14. Lavrik, I. N. (2010) Systems biology of apoptosis signaling networks. Curr. Opin. Biotechnol. 21: 551-555.
15. Kim, C. H. (2010) New Conversing Technology; Plasma Medi- cine. Korean J. Otorhinolaryngol-Head Neck Surg. 53: 593-602.
16. Yoo, G. S., J. M. Lee, C. H. Lee, J. B. Jang, and K. S. Lee (2011) Study of apoptosis by scirpi tuber in Hela cell and MCF-7 cell. J.
Korean Obstetrics Gynecol. 24: 1-13.
17. Van Engeland, M., L. J. Nieland, F. C. Ramaekers, B. Schutte, and C. P. Reutelingsperger (1998) Annexin V-affinity assay: a review on an apoptosis detection system based on phosphatidylserine exposure. Cytometry: J. Inter. Soc. Anal. Cytol. 31: 1-9.
18. Sheppard, K., K. M. Kinross, B. Solomon, R. B. Pearson, and W.
A. Phillips (2012) Targeting PI3 kinase/AKT/mTOR signaling in cancer. Crit. Rev. Oncog. 17: 69-95.
19. Park, S. Y., S. H. Lee, O. J. Park, and Y. M. Kim (2011) Apoptotic effects of curcumin and EGCG via Akt-p53 signaling pathway in HCT116 colon cancer cells. J. Life Sci. 21: 89-95.
20. Gross, A., J. M. McDonnell, and S. J. Korsmeyer (1999) BCL-2 family members and the mitochondria in apoptosis. Genes Dev.
13: 1899-1911.
21. Vander Heiden, M. G., N. S. Chandel, E. K. Williamson, P. T.
Schumacker, and C. B. Thompson (1997) Bcl-xL regulates the membrane potential and volume homeostasis of mitochondria.
Cell 91: 627-637.
22. Park, C., Y. J. Lee, Y. K. Kim, Y. C. Choi, J. H. Shin, et al. (2002) 5-FU induces apoptosis of Fas (+), HepG2 cells via activation of Fas-mediated caspase and mitochondria dysfunction. Cancer Res.
Treat. 34: 128-138.
23. Kwon, K. B., S. J. Yoo, D. G. Ryu, J. Y. Yang, H. W. Rho, et al.
(2002) Induction of apoptosis by diallyl disulfide through activa- tion of caspase-3 in human leukemia HL-60 cells. Biochem. Pharm.
63: 41-47.