41
서 론
제1형 당뇨는 췌도 세포 내에서 인슐린을 생성하는 β-세 포의 자가면역 파괴에 의해서 야기된다.(1) 당뇨병의 치료 방법으로 주목 받고 있는 췌장 또는 췌도 세포의 이식은 인슐린 주사나 인슐린 펌프에 비해 보다 근본적이고 효율적 인 혈당조절 효과를 나타내지만, 기증되는 췌장의 한계로 제약을 받는다. 또한 이식 후 초기 염증반응은 기능적인 부 적합이나 세포 괴사 및 고사에 의한 췌도 세포의 파괴를 야 기시켜, 실제 필요한 췌도 세포보다 많은 양의 세포 이식을 요구한다.(10,11) β-세포는 T-세포나 대식세포와 같은 면역 세포와의 직접적인 접촉뿐 아니라 이들 세포에서 분비하는 tumor necrosis factor-α (TNF-α), interleukin-1β (IL-1β), γ- interferon (IFN-γ)과 같은 다양한 cytokine의 자극에 의하여 파괴된다.(2-5,22) 다량의 cytokine은 췌도 세포에서 iNOS에 의한 NO 생성을 유도하는데, 이는 Krebs cycle에서 aconitase 를 억제하여, 글루코스의 산화와 ATP 생성, 그리고 인슐린 의 생성을 감소시킨다.(6-9) Prostagrandin E2 (PGE2)는 췌도 세포 또는 β-세포주에서 글루코스에 의해 유도되는 인슐린 의 분비를 억제하는 것으로 보고 되었으며, IL-1β에 의해서 생성이 촉진된다고 알려져 있다.(23) 이러한 염증성 cytokine 의 제어는 이식 초기 췌도 세포의 안정성을 유도하여 보다 소량의 세포로 그 효과를 기대할 수 있어 이식 성공률을 향상시킬 수 있을 것으로 보여진다.
Alpha-Melanocyte Stimulating Hormone Protects Pancreatic Islet Dysfunction by Peripheral Blood Mononuclear Cells in vitro
Eun-Jung Jung1, Duck-Jong Han, Sung-Ho Chang1, Dong-Gyun Lim, Yu-Mee Wee1, Jin-Hee Kim1, Yang-Hee Kim1, Sung-Kyung Koo1, Monica Choi1, Kwan Tae Park1 and Song-Cheol Kim
Department of Surgery, Ulsan University College of Medicine &
Asan Medical Center, 1Asan Institute for Life Science, Seoul, Korea
Purpose: The α-melanocyte-stimulating hormone (α-MSH) has been shown to interact with various cells of the immune and inflammatory system and down-regulate either the pro- duction or the action of the pro-inflammatory cytokines. In this study, we investigated the potential of α-MSH on preventing pancreatic islet cell from death and dysfunction by inflammatory cytokines released from peripheral blood mononuclear cells (PBMCs) in rat. Methods: Rat pancreatic islets were co-cultured with PBMCs, stimulated by phorbol myrstic acid and ionomycin. α-MSH was treated to PBMCs for 2 hours before co-culture. Viability and apoptosis of islets were observed by MTT and FACS. Inflammatory cytokines and nitric oxide (NO) were measured. Insulin release from islet co-cultured with mononuclear cells was checked for the islet function. Results: In comparison to control group, viability of islets with α-MSH treated mononuclear cells was increased and apoptosis was reduced significantly. Inflam- matory cytokines such as TNF-α and IL-1β were reduced in α-MSH-treated group. NO production in α-MSH-treated group was decreased. Insulin secretory function of islet was recovered in condition of α-MSH treatment. Conclusion:
This study demonstrates that α-MSH protects cell death and preserves the secretory function of pancreatic islet cells from the pro-inflammatory reaction of mononuclear cells, and may have the potential to improve the graft survival in clinical islet transplantation. (J Korean Soc Transplant 2006;20:41-48) ꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏ Key Words: Alpha-melanocyte-stimulating hormone, Pancre-
atic islet, Blood mononuclear cell, Anti-inflam- matory reaction
중심 단어: α-MSH, 췌장도세포, 혈액단핵세포, 항염 증반응
책임저자:김송철, 서울시 송파구 풍납2동 388-1번지 울산대학교 의과대학 외과학교실, 138-736 Tel: 02-3010-3936, Fax: 02-474-9027 E-mail: [email protected]
본 연구는 보건복지부 보건의료기술진흥사업의 지원에 의하여 이루어진 것임(과제고유번호; A050238).
혈액 단핵 세포에 의한 췌도 세포의 손상에 대한 Alpha-Melanocyte Stimulating Hormone의 세포 보호 효과
울산대학교 의과대학 외과학교실, 1서울아산병원 아산생명과학연구소
정은정1․한덕종․장성호1․임동균․위유미1․김진희1․김양희1․구선경1․최모니카1․박관태1․김송철
사람과 설치류에서 다양한 면역 반응 조절에 관여하고 있는 것으로 알려진 neuropeptides 중 하나인 α-melanocyte- stimulating hormone (α-MSH)은 pro-opiomelanocortin에서 전 이된 tridecapeptide로서 뇌하수체와 뇌, 피부, 순환기 조직에 서 발현한다.(12-14) 이는 다양한 면역 및 염증 체계에 관여 하는 세포들과 상호작용을 하여 전염증성 cytokine과 항염 증성 cytokine의 작용을 조절한다. α-MSH의 수용체(recep- tor)는 다양한 세포들에서 발현되는데, 특히 α-MSH의 주된 수용체인 melanocortin receptors I (MC1R)은 α-MSH의 항염 증 반응을 매개한다.(12,15) 그러나 아직까지 세포적 신호전 달 기전은 구체적으로 밝혀지지 않았다.
본 연구에서는 neuropeptieds인 α-MSH를 혈액 단핵 세포 에 작용시켜 췌도 세포 이식 시 발생할 수 있는 혈액 단핵 세포에 의한 전염증성 cytokines에 의한 췌도 세포의 파괴와 기능 손실에 미치는 영향을 밝히고자 하였다.
방 법
1) 췌도 세포와 혈액 단핵 세포의 분리 및 혼합 배양 췌도 세포와 혈액 단핵 세포를 얻기 위해 무게 300 g 전후 의 Lewis 백서를 사용하였다. 췌도 세포를 얻기 위해 백서 의 췌장을 분리하여 1,000 U/mL의 collagenase XI (Sigma, St.
Louis, MO, USA)로 37oC에서 20분간 조직을 소화하였다.
세포는 27%의 FicollⓇ-400 (Sigma) 용액에 혼합한 후 18%와 13%의 Ficoll 용액을 중첩하여 2,000 xg에서 20분간 원심 분 리하여 췌도 세포만을 분리하였다. 분리된 도세포는 10%
fetal bovine serum (FBS; Invitrogen, Grand Island, NY, USA) dmf 포함한 RPMI-1640 (Sigma) 배지에서 배양하였다. 혈액 단핵 세포를 얻기 위하여 백서의 간문맥에서 10 mL의 혈액 을 채취하여 HistopaqueⓇ-1077 (Density=1.077±0.001, Sigma)에 중첩하여 1,800 rpm에서 25분간 원심 분리하여 단핵 세포만 을 분리하였다.
두 세포는 TranswellTM (Corning, New York, USA)을 사용 하여 물리적 접촉 없이 배지만을 공유하는 혼합 배양을 수행 하였다.(11) 24-well plate에 Trypsin-EDTA (Invitrogen)를 처리 하여 단일 세포로 분리한 도세포를 5×105개 넣고 TranswellTM 을 넣은 후 여기에 혈액 단핵 세포를 5×105개 첨가하여 배 양하였다. 배지는 10% FBS를 포함한 RPMI1640을 사용하 였으며, 혈액 단핵 세포의 활성화를 유도하기 위하여 10 ng/
mL phorbol myristic acid (PMA; Sigma)와 1μg/mL ionomycin (Sigma)을 처리하였다.(11) 실험군에는 50 nM의 α-MSH를 단핵 세포에 2시간 동안 선처리한 후 혼합 배양하였고, 대 조군은 췌도 세포만 단독 배양하였다.
2) 췌도 세포 및 혈액 단핵 세포에서 α-MSH의 작용 에 관여하는 MC1R의 발현
췌도 세포와 혈액 단핵 세포를 배양하여 각각을 PBS
(phosphate buffered saline)로 세척하여 1 mL의 TRIzol (Isothio- cyanate and phenol, Invitrogen) 용액을 처리하여 세포를 파괴 하였다. 여기에 200μL의 Chloroform을 처리하여 12,000 xg 에서 15분간 원심분리하여 phenol 성분을 제거하고, 상층의 RNA만은 조심스럽게 따냈다. 여기에 500μL의 Isopro- phylalcohol과 75% Ethyl acohol을 단계별로 처리하여 원심 분리 세척하고 20μL의 DEPC (Diethyl pyrocarbonate, Sigma) 물에 부유시켜 총 RNA를 분리하였다. 분리된 0.5∼1μg의 total RNA는 ImProm-IITM Reverse Transcriptase (Promega, Madison, WI)를 이용하여 first strand cDNA를 합성하였다.
중합효소연쇄반응(polymerase chain reaction, PCR)은 1μL의 cDNA, 0.2μM의 각 primer, 1×PCR buffer (Takara, Otsu Shiga, Japan), 10 mM dNTP mix (Takara)와 1 unit Ex Taq DNA polymerase (Takara)를 혼합하여 PCR system (MJ Research, SanFrancisco, CA, USA)에서 시행하였다. 증폭 프로그램은 94oC에서 45초, 53oC에서 45, 72oC에서 1분으로 30 cycle 수 행하였다. 최종 PCR 산물은 1.8% agarose gel에 전기 영동 한 후 Ethidium bromide 염색하여 확인하였다. 반응에 이용 된 MC1R primer는 5'-TCTGGGGAATGTCAGGACC-3' for- ward)과 5'-TAGTGGTCTCCAGCACGATG-3' (reverse)으로, GAPDH primer는 5'-TCATGACCACAGTCCAGCCCA-3' for- ward)과 5'-GGGAFTTGCTGTTGAAGTCAC-3' (reverse)으로 제작해서 반응을 수행했다.
3) 췌도 세포의 고사(apoptosis)
췌도 세포의 고사율을 관찰하기 위하여 Annexin V로 pho- spholipids phosphatidylserine (PS)을 검출하였다. 세포 고사 가 일어날 때 PS는 extracellular surface에 축적되고, 여기에 서 형광 물질이 결합된 annexin V가 결합하여 검출된다. 혼 합 배양에서 췌도 세포만을 분리하여 5μL의 Annexin V (BD Bioscience Pharmingen, San Jose, CA, USA)와 10μL의 Propidium Iodide (PI; Sigma)를 처리하였다.(1) 15분간 암소 에 보관한 후 fluorescence activated cell sorter (FACS; Becton Dickinson Facs caliber, San Jose, California, USA)로 세포 고 사율을 분석하였다.
4) 췌도 세포의 생존율
혼합 배양에 따른 췌도 세포의 생존율을 관찰하기 위하 여 MTT assay를 수행하였다. 혼합 배양에서 췌도 세포만을 분리하여 0.5 mg/mL의 MTT (3-(4,5-dimethylthiazol-2-yl)-,5- diphenyletrazolium bromide; Sigma)를 처리하여 2∼4시간 정 도 배양한 후, 배지를 제거하고, 100μL의 DMSO (Sigma)와 glycin buffer를 처리하여 570 nm에서 흡광도를 측정(Spectra MAS PLUS, Molecular devices)하여 상대적인 생존율을 비 교하였다.
5) Nitric Oxide (NO) 생성
세포에서 NO 생성을 확인하기 위하여 혼합 배양한 세포 상청액을 96-well plate에 넣고, Griess Reagent (p-amino- benzene sulfonamide N-(1-naphthyl)ethylene-diamine in O-phos- phoic acid; Promega)를 처리하였다. 10분간 암소에서 반응 을 유도한 후 540 nm에서 흡광도를 측정하여 NO 생성량을 비교 분석하였다.
6) 염증성 cytokine의 분비
염증성 cytokine의 생성에 미치는 α-MSH의 효과를 분석 하기 위하여 혼합 배양한 세포 상청액을 모아 TNF-α, IL-1β 의 분비량을 측정하였다. ELISA를 수행하기 위하여 각 cytokine에 대한 ELISA kit (R&Dsystems Inc., Minneapolis, MN, USA)를 사용하였다. 각 cytokine에 대한 항체가 부착된 96-well plate에 세포 배양액과 표준 용액 50μL를 넣어 2시 간 동안 반응시켜 4회 세척하고, 100μL의 HRP가 결합된 2차 항체를 첨가하여 1시간 동안 상온에 배양하였다. 동일 한 세척 과정을 수행하고 100μL의 기질을 처리하여 30분 간 발색을 유도하였다. 최종적으로 50μL의 정지액(1M H2SO4)을 처리해서 450 nm에서 흡광도를 측정하여 그룹별 cytokine 분비능을 비교 분석하였다.
7) PGE2의 생성
PGE2 생성에 미치는 α-MSH의 효과를 분석하기 위하여 혼합 배양한 세포 상청액을 모아 PGE2의 분비량을 측정하 였다. 본 연구에서는 PGE2 ELISA kit (R&Dsystems)를 사용 하였다. PGE2에 대한 항체가 부착된 96-well plate에 세포 배 양액과 표준 용액을 50μL씩 넣어 2시간 동안 반응시켜 4회 세척하고, 100μL의 HRP가 결합된 2차 항체를 첨가하여 1 시간 동안 상온에 배양하였다. 동일한 세척 과정을 수행하 고 100μL의 기질을 처리하여 30분간 발색을 유도하였다.
최종적으로 50μL의 정지액(1M H2SO4)을 처리해서 450 nm 에서 흡광도를 측정하여 그룹별 PGE2 분비능을 비교 분석 하였다.
8) 췌도 세포 인슐린 분비능 분석
세포에 α-MSH를 처리한 그룹과 그렇지 않은 그룹을 구 분하여 세포 배양 상청액을 모아 췌도 세포의 인슐린 분비 능을 측정하기 위하여 Insulin assay ELISA kit (SHIBAYAGI co., ltd., Japan)를 사용하였다. 배양 상청액은 단계별로 희석 (1/10, 1/50, 1/100, 1/500)하여 최적의 분석 농도를 결정하였 다. 96-well plate에 biotin-conjugated anti-insulin을 처리한 후 10μL의 세포 배양액과 표준 용액을 넣어 2시간 반응시켰 다. Plate를 세척액으로 4회 세척하여 물기를 완전히 제거한 후 100μL의 HRP가 결합된 streptavidin 용액을 넣고 30분간 배양하였다. 같은 방법으로 세척하여 100μL의 substrate
chromogen reagent를 30분간 처리하고, 100μL의 정지액 (1M H2SO4)을 이용해 반응을 정지시키고 450 nm에서 흡광 도를 측정하였다. 표준화 곡선을 이용하여 각 그룹별로 생 성된 인슐린 양을 정량 비교하였다.
9) 통계학적 분석
결과의 통계학적 분석은 Student's t-test를 수행하였으며, P<0.05로 통계학적 의의를 부여하였다.
결 과 1) 세포에서 MC1R의 발현
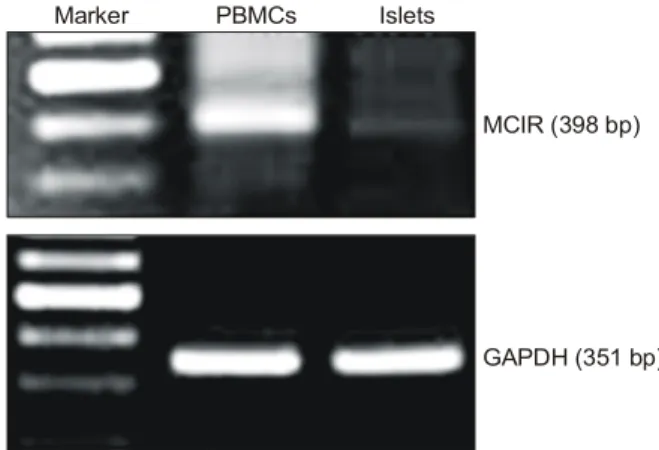
백서의 췌장세포와 혈액 단핵 세포에서의 MC1R의 발현을 확인하기 위하여 RT-PCR을 수행하였다. 그 결과, 두 세포 모 두에서 MC1R의 발현이 관찰되었다. 그러나 혈액 단핵 세포 의 MC1R 발현 정도가 췌도 세포에서 발현하는 MC1R보다 현저하게 많아 두 세포 사이에 차이를 나타냈다(Fig. 1).
2) 최적 효과를 얻기 위한 α-MSH의 농도
본 연구에서는 최적의 α-MSH 농도를 결정하기 위하여 10 nM, 50 nM, 100 nM, 500 nM 농도의 α-MSH를 처리하여 그 효과를 비교하였다. 췌도 세포의 생존율에 영향을 미치 지 않는 조건에서 α-MSH가 NO생성을 억제하는 효과를 비 교한 결과, 10 nM에서는 16% 정도 감소하여, 효과가 미미하 였으나, 50 nM에서 500 nM까지의 농도에서는 30∼50% 정
Fig. 1. Expression of Melanocortin receptor I (MC1R) on pancre- atic islet and peripheral blood mononuclear cells (PBMCs).
Total RNA was purified from cultured pancreatic islets and PBMCs and each samples were carried out RT-PCR for MC1R. GAPDH was used as positive and internal control.
Expression of MC1R was detected on islet and PBMCs.
But, the band intensity of them was different. MC1R expression of pancreatic islet was very weak compared with PBMCs. Data were representative of the triplicate of three independent experiments.
Marker PBMCs Islets
MCIR (398 bp)
GAPDH (351 bp)
도 NO형성 억제 효과를 나타내었다. NO 억제제로서 1mM 의 Nω-Nitro-L-arginine methyl ester (L-NAME; Calbiocam, Merck KGaA, Germany)를 처리하여 37.4%의 억제 효과를 관찰하였다(Fig. 2).
3) α-MSH에 의한 췌도 세포의 생존율 및 고사율의 변화
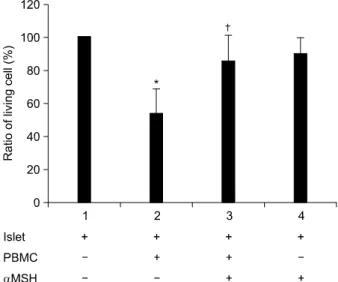
α-MSH 처리 후 세포의 생존율을 MTT를 이용하여 비교 한 결과, 췌도 세포 단독배양을 기준으로 하여, 단핵 세포와 혼합 배양한 췌도 세포의 경우 53.8%로 생존율이 현저하게 감소하였으며, α-MSH를 처리한 경우 생존율이 85.8%로 증 가하여 단핵 세포에 의한 췌도 세포의 파괴를 감소시키는 것으로 나타났다(P<0.05)(Fig. 3). 또한 췌도 세포 고사 정 도를 분석한 결과, α-MSH를 처리한 그룹에서 단핵 세포와 의 혼합 배양으로 인한 췌도 세포의 고사율이 감소하였다 (Fig. 4). 췌도 세포만 단독 배양했을때 38.2%였으나, PMA/
Ionomycin으로 자극한 단핵 세포를 함께 배양한 경우 췌도 세포의 고사율은 56.8%로 증가하였다. 여기에 α-MSH처리 한 경우 43.2%로 고사율이 감소하였다(P<0.05).
4) α-MSH에 의한 췌도 세포에서 생성되는 NO 생성 량의 변화
췌도 세포 단독 배양한 대조군의 nitrite 생성량은 3.1±
Fig. 3. Islet viability by PMA/Ionomycin treated PBMCs. Pan- creatic islets were cultured with PMA/I treated PBMCs with or without the α-MSH for 48 hr. Islets were repre- sented on fresh 96-well plate for MTT assay. 10μL of MTT solution (5 mg/mL) was added to each well. It was incubated for 2 hr and treated DMSO and glycin buffer.
Optical density was detected. Viability of co-cultured islets was decreased. Viability of α-MSH treated group was recovered compared to co-cultured islets. Each experiment was performed in triplicate and repeated three times for reproducibility. *P<0.05 vs. lane 1, †P<0.05 vs. lane 2.
Fig. 2. Inhibition of NO production as the concentration of α- MSH. Different concentrations of α-MSH were treated to PBMCs before co-culture with pancreatic islet. Islet cells were co-cultured with pretreated PBMCs and the culture supernatants were used for detection of NO production.
30∼40% of NO production was inhibited by treatment of 50 nM to 500 nM concentration of α-MSH. ω-Nitro-L- arginine methyl ester (L-NAME) was used for inhibitor of NO production.
Fig. 4. Apoptosis of pancreatic islet by PMA/I treated PBMCs.
Pancreatic islets were cultured with PMA/I treated PBMCs on TranswellTM for 48 hr. Islets were collected and washed with PBS. Apoptotic islets were stained with Annexin V
& PI. They were analyzed with FACS. Apoptotic cell ratio of co-cultured islets was recovered after α-MSH treat- ment. Each experiment was performed in triplicate and repeated three times for reproducibility. *P<0.05 vs. lane 1, †P<0.05 vs. lane 2.
0.34μM이었으나, 단핵 세포와의 혼합 배양에서 7.6±0.7μM 로 급격한 증가량을 나타내었다. 그러나, α-MSH 처리 그룹 의 경우 4.2±0.4μM로 감소하였다(P<0.05). 췌도 세포에 α-MSH만을 처리한 경우는 3.3±0.09μM로 대조군과 통계 학적 차이를 나타내지 않았다(Fig. 5). α-MSH 단독으로는 췌도 세포의 nitrite 생성에 영향을 미치지 않음을 알 수 있 었다.
5) α-MSH에 의한 배양액에 분비된 염증성 cytokine 의 변화
전염증성 cytokine인 TNF-α의 변화를 관찰한 결과, 대조 군의 경우 70.2±5.9 pg/mL였으나, 단핵 세포와 혼합 배양한 경우 122.5±8.4 pg/mL로 급격히 증가하였다(Fig. 6). α- MSH를 처리한 그룹에서는 77.6±12.7 pg/mL로 대조군과 유사한 양으로 감소하였다(P<0.05). 또한 IL-1β의 경우, 대 조군에서 18.1±7.6 pg/mL이었으나, 혼합 배양에서 185.4±
43.6 pg/mL로 증가하였다. α-MSH를 처리한 경우 IL-1β의 생성량이 130.9±26.1 pg/mL로 소폭 감소하였다(Fig. 7). 본 결과는 α-MSH가 다양한 염증성 cytokine의 생성을 억제하 는 것으로 나타났다.
6) 혼합 배양에서 분비된 PGE2량에 미치는 α-MSH 의 효과
세포 배양 상청액의 PGE2 생성량을 관찰한 결과, 대조군 에서 1382±353.3 pg/mL이었으며, 혼합 배양시 2523.8±213
pg/mL로 증가하였으며(P<0.05), α-MSH 존재시에 2065±
265.4 pg/mL로 소량 감소하는 듯 하지만 통계학적으로 의 의가 없었다(P>0.05)(Fig. 8).
Fig. 6. TNF-α release from the islet/PBMCs co-culture. Pancreatic islets were cultured with PMA/I treated PBMCs. Con- centration of TNF-α on culture supernatant was measured by ELISA. In comparison to control, TNF-α release was increased in co-cultured group. And it was decreased significantly in α-MSH treated group. Each experiment was performed in triplicate and repeated three times for reproducibility. *P<0.05 vs. lane 1, †P<0.05 vs. lane 2.
Fig. 7. IL-1β release in the islet/PBMCs co-culture. Pancreatic islets were cultured with PMA/I treated PBMCs on TranswellTM for 48 hr. Quantity of IL-1β was measured by ELISA. In comparison to control, concentration of IL-1β was increased in co-cultured group significantly. And it was recovered in α-MSH treated group. Each experiment was performed in triplicate and repeated three times for reproducibility. *P<0.05 vs. lane 1, †P<0.1 vs. lane 2.
Fig. 5. Nitrite production from the islet/PBMCs co-culture. Pancre- atic islets were cultured with PMA/I treated PBMCs for 48 hr. The supernatant was collected from each well. Pro- duction of nitrite was calculated with Griess reagent.
Concentration of nitrite in co-cultured group was decreased after α-MSH treatment. Each experiment was performed in triplicate and repeated three times for reproducibility. *P<
0.05 vs. lane 1, †P<0.05 vs. lane 2.
7) 인슐린 분비능의 변화
췌도 세포의 기능 변화를 관찰하기 위하여 인슐린 분비 량을 비교 분석한 결과, 대조군의 경우 522.6±85 ng/mL이 었던 인슐린 생성량이 단핵 세포와 혼합 배양으로 인하여 277.5±18.6 ng/mL로 감소하였다(Fig. 9). 그러나 α-MSH의 존재하에서 췌도 세포의 인슐린 분비량은 471.0±62.9 ng/mL로 증가하였다(P<0.05). α-MSH만을 처리한 그룹에 서는 인슐린 분비량이 509.5±21.0 ng/mL로 췌도 세포 단독 배양과는 통계학적으로 의의가 없었다. 이 결과는 α-MSH 의 처리는 단핵 세포와의 혼합 배양으로 손상된 췌도 세포 의 인슐린 분비 기능의 손상이 보호되는 것으로 관찰되었 다.
고 찰
α-MSH는 백혈구, 림프구 등과 같은 면역 세포에서 생성 되는 TNF-α, IL-1β와 같은 염증성 cytokine에 의한 세포 파 괴를 억제하는 항염증 효과를 가지고 있어 염증 반응을 조 절하는 역할을 하는 것으로 보고되고 있다.(12-16) 본 연구 에서는 이러한 α-MSH의 항염증 효과를 이용하여, 췌도 세 포 이식 시 발생하는 염증에 의한 췌도 세포의 파괴 및 기 능 상실을 억제하고자 하였다.
α-MSH가 세포에 반응하는 기전에는 5종의 melanocortin receptor (MC1R-MC5R)들이 관여하고 있는데, 이 중 MC1R 은 다양한 염증 및 면역세포의 표면에 발현하여 α-MSH의
항염증 반응을 조절하는 데 중요한 역할을 한다.(12-16) 현 재까지 인체와 생쥐의 췌도 세포에서 MC1R의 발현이 보고 되었으나, 구체적인 α-MSH와 췌도 세포와의 관계를 규명 하는 보고는 없었다. 본 연구에서 백서의 단핵 세포와 췌도 세포에서의 MC1R의 발현을 관찰하였다. 두 세포에서 모두 MC1R의 발현이 확인되었으나, 이들의 발현양에는 다소 차 이를 보였다. 혈액 단핵 세포에서 발현하는 MC1R에 비해 췌도 세포에서 발현하는 양이 작은 것으로 나타났는데, 이 는 췌도 세포가 혈액 단핵 세포에 비해 α-MSH에 대한 작 용이 미약할 수 있음을 암시하였다. 이를 확인하기 위하여 췌도 세포에 직접 α-MSH를 처리하여 항염증 및 NO 억제 여부를 관찰한 결과, α-MSH의 효과가 거의 나타나지 않았 다. Hill 등(15)은 인체 피부 섬유아세포가 다른 멜라닌 세포 들에 비해 MC1R의 발현이 낮아 α-MSH에 의한 세포 보호 효과가 다른 세포들에 비해 낮다고 보고하였다. Sarkar 등 (14)은 Jurkat (T-cell line) 세포와 대식세포, PMA로 자극한 THP-1 (macrophage cell line) 세포, U-937 (monocyte cell line) 세포, 호중구(neutrophil), H4 (glioma cell line), HeLa 세포, NIH 3T3 (fibroblast) 세포에서 MC1R 특이 항체를 이용한 α-MSH 결합 정도를 분석하였다. 그 결과에 따르면 면역작 용에 관여하는 대식세포와 호중구, T-세포 등에서의 결합률 이 높은 것으로 관찰되었다. 이는 세포마다 존재하는 MC1R 에 차이가 있고 이로 인해 α-MSH의 작용에도 차이가 나는 것으로 보고하였다. 본 연구에서도 췌도 세포에서 발현되 Fig. 8. PGE2 release in the islet/PBMC co-culture with TranswellTM.
Pancreatic islets were cultured with PMA/I treated PBMCs for 48 hr. In comparison to control, PGE2 release was increased in co-cultured group significantly. It was not recovered in α-MSH treated group. Each experiment was performed in triplicate and repeated three times for reproducibility. *P<0.05 vs. lane 1, †P>0.05 vs. lane 2.
Fig. 9. Insulin release from the islet/PBMCs co-culture with TranswellTM. The supernatant was diluted with dilution buffer as the ratio of 1/500. Concentration of insulin was measured by ELISA. In comparison to control, insulin release was decreased in co-cultured group significantly.
And it was progressed in α-MSH treated group. Each experiment was performed in triplicate and repeated three times for reproducibility. *P<0.05 vs. lane 1, †P<0.05 vs. lane 2.
는 MC1R의 양이 미약하여 α-MSH에 효과적으로 반응하지 못하는 것으로 여겨진다. 그러므로, α-MSH를 면역세포에 선 처리하여 췌도 세포의 보호를 유도하는 것이 보다 효과 적임을 알 수 있다.
췌도 세포 이식에서 발생할 수 있는 다양한 염증 세포와 이들에서 분비하는 독성 물질에 의한 췌도 세포의 손상을 유도하는 in vitro 환경을 만들기 위하여 TranswellTM을 사용 하였다. 두 세포가 한 well에 존재하는 직접 접촉법(direct contact method)으로 배양한 경우, 보다 간편한 방법이나 다 양한 요소가 배양 환경에 존재하여 용해성 물질(souble factor)이 미치는 영향을 분석하기 어렵고, 혼합 배양에서 순 수한 췌도 세포만의 생존율을 분석하기가 용이하지 않았 다. TranswellTM의 사용은 물리적인 접촉을 배제하고, 배지 만을 공유하도록 하여 단핵 세포에서 분비된 용해성 물질 에 의한 췌도 세포의 손상에 대한 연구를 수행하기 위한 최적의 환경을 제공하였다. 이 TranswellTM 시스템(17)은 새 로운 염증 억제제의 연구와 다양한 염증 세포와 췌도 세포 의 상호작용 연구 등에 다양하게 응용할 수 있을 것으로 생각한다.
최근 여러 논문들이 α-MSH는 nanomole의 농도 범위에 서 항염증 효과가 최대라고 보고하고 있다.(12-16) 본 연구 에서는 최적의 α-MSH의 농도를 정하기 위하여, 다양한 nanomole 농도의 α-MSH를 처리하여 혼합 배양에서 그 효 과를 비교하였다. 그 결과 50 nM에서 500 nM까지의 농도에 서는 30∼50% 정도 NO형성 억제 효과를 나타내었다. 또한, 이 농도 범위에서 α-MSH만으로는 췌도 세포 생존율과 NO 생성에 어떠한 영향도 미치지 않았다. 같은 양의 α-MSH를 처리하여 혼합 배양하였을 때, 단핵 세포에 의해 손상되는 췌 도 세포의 생존율은 증가시키고, 다양한 염증 관련 cytokine의 생성은 효과적으로 억제되었다. 이는 다양한 면역 세포가 존재하는 혈액 단핵 세포와의 혼합 배양에서 이들에 의한 췌도 세포의 파괴와 기능 상실을 염증 세포에 α-MSH를 선 처리하는 것을 통하여 일부 예방할 수 있음을 보여주었다.
본 결과에서 대표적인 염증 관련 cytokine으로 알려진 TNF- α와 IL-1β, 그리고 NO는 혼합 배양에 의해서 각각의 생성 량이 현저하게 증가하였고, α-MSH의 처리에 따라 감소하 여 α-MSH가 이들의 생성을 억제하는 것을 확인할 수 있었 다. TNF-α는 췌도 세포의 고사에 직접적으로 관여하 고,(18) 이식 부위에서 adhesion molecule을 증가시켜 이식 거부 반응을 유도한다. IL-1β는 T-cell을 자극하여 면역 거 부의 주요 원인이 되는 IL-2를 포함한 다양한 면역 관련 인 자들의 작용을 촉진한다. 또한 NO도 대식세포를 활성화시 키고, 이들 자체가 세포에 독성 요인으로 작용한다.(4,8,18-20) 따라서 이들의 조절은 췌도 이식 시 췌도세포의 염증성 및 면역학적 손상을 최소화할 수 있을 것으로 생각한다.
또한, 염증 반응 시 증가되는 것으로 알려진 PGE2의 생성 량을 분석한 결과 혼합 배양에 의해 PGE2의 생성량이 증가
하였으나, α-MSH의 처리에 따른 변화는 나타나지 않았다.
이는 혈액 단핵 세포에 의해서 나타나는 췌도 세포의 손상 은 PGE2의 생성과 다른 기전이 관여하는 것으로 보여진다.
Heitmeier 등(2)의 연구에 따르면 다양한 세포에서 NO나 cytokines에 의한 cyclooxygenase-2 (COX-2)의 활성은 PGE2
의 생성을 증가시키지만, 이러한 PGE2의 생성이나 COX2 의 작용이 췌도 세포의 파괴나 기능 손상에 관여하지는 않 는 것으로 보고하였다. 이에 대해서는 여러가지 다양한 보 고(21)가 있으나 그 기작이 정확하게 밝혀지지 않아 지속적 인 연구가 필요하다.
이렇듯 혼합 배양에 의해 분비되는 다양한 독성 인자들 에 의해 췌도 세포가 손상을 입게 되면 췌도 세포에 존재하 는 β-세포의 인슐린 분비능에도 문제가 발생한다. 세포수 의 감소와 인슐린 분비를 억제하는 cytokine의 작용에 따라 감소된 인슐린 분비량이 α-MSH의 처리 그룹에서 회복되 는 양상을 나타내었다. 이는 α-MSH에 의한 면역 억제 반응 이 췌도 세포를 보호하기 때문이다.(15,16) 본 결과들은 α- MSH가 염증 세포에서 생성되는 다양한 독성 물질의 생성 을 억제하여 췌도 세포 이식 시 췌도 세포의 손실을 방지하 여 이식 효율을 향상시킬 것으로 생각한다. 이를 위하여 α- MSH에 대한 구체적인 기전을 밝히기 위한 연구가 수행되 어야 하며, 더불어 당뇨 동물에서의 α-MSH의 효과를 밝히 는 전 임상 연구가 수행되어야 할 것으로 생각한다.
결 론
본 연구에서 α-MSH는 혈액 단핵 세포에서 분비되는 다 양한 염증성 cytokine과 NO 등에 의한 췌도 세포의 파괴 및 인슐린 분비능의 손상을 방지하였다. 이는 궁극적으로 췌 도 세포 이식 시 발생하는 염증 반응 및 이에 수반되는 면 역 작용을 감소시켜 췌도 세포 이식 효율을 향상시킬 수 있을 것으로 생각한다.
REFERENCES
1) Yang Z, Chen M, Ellett JD, Carter JD, Brayman KL, Nadler JL. Inflammatory blockade improves human pancreatic islet function and viability. Am J Transplant 2005;5:475-83.
2) Heitmeier MR, Kelly CB, Ensor NJ, Gibson KA, Mullis KG, Corbett JA, Maziasz TJ. Role of cyclooxygenase-2 in cytokine- induced beta-cell dysfunction and damage by isolated rat and human islets. J Biol Chem 2004;279:53145-51.
3) Han X, Sun Y, Scott S, Bleich D. Tissue inhibitor of metallo- proteinase-1 prevents cytokine-mediated dysfunction and cyto- toxicity in pancreatic islets and beta-cells. Diabetes 2001;
50:1047-55.
4) Kuttler B, Steveling A, Kloting N, Morgenstern O, Wanka H.
Aminoguanidine downregulates expression of cytokine-induced
Fas and inducible nitric oxide synthase but not cytokine- enhanced surface antigens of rat islet cells. Biochem Pharmacol 2003;66:2437-48.
5) Bregenholt S, Moldrup A, Blume N, Karlsen AE, Nissen Friedrichsen B, Tornhave D, Knudsen LB, Petersen JS. The long-acting glucagon-like peptide-1 analogue, liraglutide, in- hibits beta-cell apoptosis in vitro. Biochem Biophys Res Commun 2005;330:577-84.
6) Eizirik DL, Pavlovic D. Is there a role for nitric oxide in beta-cell dysfunction and damage in IDDM? Diabetes Metab Rev 1997;13:293-307.
7) Kang MK, Yoon YE, Yang JY, Kwon KB, Park JW, Jhee EC.
Protective effect of retinoic acid on interleukin-1 beta-induced cytotoxicity of pancreatic beta-cells. Mech Ageing Dev 2004;
125:483-90.
8) Baker MS, Chen X, Cao XC, Kaufman DB. Expression of a dominant negative inhibitor of NF-kappaB protects MIN6 beta- cells from cytokine-induced apoptosis. J Surg Res 2001;97:
117-22.
9) Corbett JA, McDaniel ML. Reversibility of interleukin-1 beta- induced islet destruction and dysfunction by the inhibition of nitric oxide synthase. Biochem J 1994;299:719-24.
10) Choi SE, Choi KM, Yoon IH, Shin JY, Kim JS, Park WY, Han DJ, Kim SC, Ahn C, Kim JY, Hwang ES, Cha CY, Szot GL, Yoon KH, Park CG. IL-6 protects pancreatic islet beta cells from pro-inflammatory cytokines-induced cell death and functional impairment in vitro and in vivo. Transpl Immunol 2004;13:43-53.
11) Tran VV, Chen G, Newgard CB, Hohmeier HE. Discrete and complementary mechanisms of protection of beta-cells against cytokine-induced and oxidative damage achieved by bcl-2 overexpression and a cytokine selection strategy. Diabetes 2003;
52:1423-32.
12) Yoon SW, Goh SH, Chun JS, Cho EW, Lee MK, Kim KL, Kim JJ, Kim CJ, Poo H. alpha-Melanocyte-stimulating hor- mone inhibits lipopolysaccharide-induced tumor necrosis factor- alpha production in leukocytes by modulating protein kinase A, p38 kinase, and nuclear factor kappa B signaling pathways.
J Biol Chem 2003;278:32914-20.
13) Lipton JM, Zhao H, Ichiyama T, Barsh GS, Catania A. Mecha-
nisms of antiinflammatory action of alpha-MSH peptides. In vivo and in vitro evidence. Ann N Y Acad Sci 1999;885:
173-82.
14) Sarkar A, Sreenivasan Y, Manna SK. alpha-Melanocyte-stimu- lating hormone inhibits lipopolysaccharide-induced biological responses by downregulating CD14 from macrophages. FEBS Lett 2003;553:286-94.
15) Hill RP, Wheeler R, Macneil S, Haycock JW. Alpha-melano- cyte-stimulating hormone cytoprotective biology in human dermal fibroblast cells. Peptides 2005;26:1150-8.
16) Sarkar A, Sreenivasan Y, Manna SK. Alpha-melanocyte-stimu- lating hormone induces cell death in mast cells: involvement of NF-kappaB. FEBS Lett 2003;549:87-93.
17) Hamaguchi K, Utsunomiya N, Takaki R, Yoshimatsu H, Sakata T. Cellular interaction between mouse pancreatic alpha-cell and beta-cell lines: possible contact-dependent inhibition of insulin secretion. Exp Biol Med (Maywood) 2003;228:1227- 33.
18) Chang I, Kim S, Kim JY, Cho N, Kim YH, Kim HS, Lee MK, Kim KW, Lee MS. Nuclear factor kappaB protects pancreatic beta-cells from tumor necrosis factor-alpha-medi- ated apoptosis. Diabetes 2003;52:1169-75.
19) Scarim AL, Heitmeier MR, Corbett JA. Heat shock inhibits cytokine-induced nitric oxide synthase expression by rat and human islets. Endocrinology 1998;139:5050-7.
20) Jang JY, Lee DY, Park SJ, Byun Y. Immune reactions of lymphocytes and macrophages against PEG-grafted pancreatic islets. Biomaterials 2004;25:3663-9.
21) Matsuda T, Omori K, Vuong T, Pascual M, Valiente L, Ferreri K, Todorov I, Kuroda Y, Smith CV, Kandeel F, Mullen Y.
Inhibition of p38 pathway suppresses human islet production of pro-inflammatory cytokines and improves islet graft func- tion. Am J Transplant 2005;5:484-93.
22) Yang Z, Chen M, Ellett JD, Fialkow LB, Carter JD, Nadler JL. The novel anti-inflammatory agent lisofylline prevents autoimmune diabetic recurrence after islet transplantation.
Transplantation 2004;77:55-60.
23) Andersson AK, Thorvaldson L, Carlsson C, Sandler S. cytokine- induced PGE(2) formation is reduced from iNOS deficient murine islets. Mol Cell Endocrinol 2004;220:21-9.