Copyrightⓒ2009, The Microbiological Society of Korea
324
능이버섯(Sarcodon aspratus) 무름병소에서 분리한 MK1 균주의 최적 성장조건과 정제된 균체외다당류의 특성
유정은·이영남*
충북대학교 자연과학대학 미생물학과
능이버섯(Sarcodon aspratus)의 무름병소에서 분리한 MK1 균주는 절대호기성 타가영양 세균으로 거식세포의 활성을 유도하는 균체외다당류를 다량 생산한다. MK1의 최적 성장 조건과 MK1이 생산하는 균체외다당류의 몇 가지 물리적 성질을 알아보았다. MK1은 glucose, galactose, fructose, sucrose를 탄소원으로 잘 이용하나 MK1의 lactose 이용은 다소 저조하였다. 한편 potato starch와 dextrin을 이용하지 못하였다. MK1은 pH 7.0, 30oC, 200 rpm, 2% glucose, 0.05~0.2% (NH4)2SO4에서 최적 성장을 보였다. 최적 성장조건에서 얻은 균체외다당류 (EPSopt)는 탄소(37.1%), 질소(2.2%), 산소(49.3%), 수소(6.4%)로 구성되었으며, 황(S)은 검출되지 않았다. 그리고 아미노당과 산성 당을 함유한 이종다당류로 여겨지며, SDS-polyacrylamide gel elctrophoresis로 분석한 EPSopt의 분자량은 14.8~47.9 kDa였다. Glucose 배지에서 생산된 균체외다당류(EPSglu)의 점도를 측정한 바, pH 6.0과 7.5에 서 얻은 것들(0.1 g/ml)의 상대 점도(´ηrel)는 각각 1.23과 1.39이었다. 또한 형태 또는 질감에도 차이가 있었는데, 배 지의 pH가 낮을수록 미세결정형을, pH가 높아질수록 끈끈한 질감을 보였다. 따라서 배지 pH가 균체외다당류의 물리적 성질에 영향을 주는 것으로 사료된다.
Key words□ exopolysaccharide, heteropolysaccharide, MK1 strain, Neungee (Sarcodon aspratus), optimal culture, rotten tissue
우리나라에서 능이버섯[Sarcodon aspratus (Berk) S. Ito]은 송 이(Tricholoma matsutake), 표고(Lentinula edodes)와 더불어 선호 도가 높은 식용 버섯으로 토양이 비옥하고 습한 산림에 자생하 는 능이는 맛과 향이 뛰어나 향버섯이라고도 한다(17, 47). 담자 균강 민주름버섯목 굴뚝버섯과에 속하는 능이는 참나무 수종 (Quercus mongolica, Q. dentata)의 뿌리에 공생하는 균근성 버섯 (ectomycorrhizal mushroom)으로 아직 노지재배조차 되지 않는다.
한국과 일본 등에서 수요가 큰 능이는 생산량이 부족하고 노동 집약적 채집으로 고가이다(20). 예로부터 민간에서 육류를 먹고 체했을 때 능이버섯의 끓인 물을 복용하였으며, 근래 다른 식용 버섯보다 능이의 단백분해효소 활성이 탁월하다는 연구도 있다 (46, 58). 뿐만 아니라, 능이버섯에 면역조절물질(37), 항산화물질 (19), 세포 사멸 유도활성(37), 혈중 콜레스테롤 저하 물질 등 다 양한 생리활성이 있음이 보고되었다(47, 62).
능이버섯은 수확 후 수일이면 버섯의 뒷면 갓이 점액성 물질 로 뒤덮여(무름병) 식용가치를 잃어버린다. 무름병 원인을 규명 하고자 무름병소에서 미생물을 탐색하였는데, 광합성 세균 한 균 주, 타가 영양균 네 균주를 분리·동정하였다(27). 타가영양세균 중 성장속도가 매우 빠른 MK1 균주는 신종의 세균으로 추정되 며 나머지 타가영양성 세균은 Burkholderia cepacia complex나
Methylobacter sp.로 동정되었다. 그람 음성 간균 절대호기성 세 균인 MK1 균주는 점액성 물질을 생산하여 능이의 무름병을 주 도하는 것으로 사료되며(27), 따라서 능이 무름병의 방제 방안도 보고되었다(41).
MK1 균체를 둘러싸고 있는 유백색 점성물질은 균체외다당류 (exopolysaccharides, EPS)로 여겨진다. 미생물의 체외다당류 물질 은 세포벽에 단단히 결합된 협막 다당류(capsular polysaccharide) 와 세포 표면에 느슨하게 부착되어 쉽게 균체 밖으로 유리되는 점액성 다당류(slime polysaccharide)로 구분된다(6, 25, 52). 미생 물 균체외다당류는 자연생태계에서 생물막 형성이나 항생물질, 독성 물질, 삼투압, 건조, 자외선으로부터 미생물 세포를 보호하 는 등 생존에 매우 중요하며(50, 56), 동·식물에 병독성 인자로 작용하기도 한다(45). 한편, 균체외다당류는 안정제, 유화제, 현탁 제, 점증제, 겔 형성제, 교결제, 응집제, 피막제, 수분흡수제, 접착 제 등 여러 용도가 있어 식품, 의료 제약, 향장, 농업, 화학, 기 계 등 각종 분야에 산업 소재로 높은 상업적 가치를 지닌다(1, 21, 40). 예를 들면, 1950년대 Leuconostoc mensenteroides가 생 산하는 덱스트란이 혈장 대용제로 개발된 것을 위시하여 식품의 맛이나 질감의 향상을 위하여 첨가하는 Lactobacillus sp.가 생산 하는 균체외다당류(57), 중금속 흡착력이 우수하여 자연환경의 생치유에 활용되는 Alteromonas의 균체외다당류(29), 화장품의 보습물질로 들어가는 Streptococcus equi의 히아눌론산(49) 등이 있다. 뿐만 아니라 Bacillus licheniformis에서 얻은 균체외다당류 는 면역조절기능과 항바이러스활성을(1), Gluconoacetobacter
*To whom correspondence should be addressed.
Tel: 82-43-261-2301, Fax: 82-43-264-9600 E-mail: [email protected]
xylinus, Rhanella aquatilis, Zymomonas mobilis 등 세균이 생산 하는 레반은 항종양활성을 지닌 것으로 보고되었다(61).
거식세포의 활성화(macrophage activation)로 개시되는 면역계 는 감염 또는 암세포 증식에 대항하는 매우 중요한 숙주의 방어 기작인데(63), 거식세포가 항원물질, 감염체 등에 의하여 활성화 되면 nitric oxide와 cytokine 같은 면역조절물질의 생산이 증가한 다. 거식세포 활성인자로 식물과 미생물의 균체외다당류가 주목 을 받는데(1, 7, 59), Lactobacillus delbrueckii subsp. bulgaricus 의 균체외다당류, B. licheniformis에서 얻은 균체외다당류 등이 현저하게 거식세포 활성을 유도한다는 보고가 있다(22, 39).
우리는 예비실험에서 MK1에서 얻은 균체외다당류가 쥐의 거 식세포에서 nitric oxide, TNF-α, IL-1β 생산을 현저하게 증가시 키는 것을 관찰하였다(28). 이에 MK1의 균체외다당류가 유용한 생리활성물질로 개발 될 수 있다고 사료되어, 균체외다당류의 생 산, 물성, 화학 구조 및 생리-생화학 성질에 관심을 갖게 되었다.
본 논문에서는 우선 MK1 균주의 최적 성장 조건을 알아보고 최적 성장 환경에서 얻은 균체외다당류의 몇 가지 물리화학적 성상을 조사하였다.
재료 및 방법 사용 균주 및 배양
MK1 균주의 증식에는 LB 배지(Difco)를 사용하였다. 기본 무 기염 배지는 1 L DW에 Na2HPO4 4.8 g, KH2PO4 4.4 g, MgSO4·7H2O 500 mg, FeCl3 50 mg, CaCl2 5 mg을 넣어 조성하였 고, Na2HPO4, KH2PO4를 넣어 배지의 pH를 6.8±0.2로 맞추고 탄 소원과 질소원을 보충하였다(23). 탄소원으로 glucose, galactose, fructose, sucrose, lactose, dextrin, potato starch 등을, 질소원으로 는 NH4Cl, (NH4)2SO4, (NH4)2HPO4, yeast extract를 사용하였다.
최적 C/N (w/w) 비를 구할 때는 2% glucose에 각기 다른 농도 의 (NH4)2SO4를 첨가하여 조성하였다. MK1의 최적 성장에 요구 되는 pH, 교반 속도, 온도는 2%의 glucose가 들어있고 탄소/질 소비가 10/1로 조성된 무기염 배지에서 조사하였다. 하루 밤 LB 배양액에서 자란 MK1 균액 10 ml을 탄소, 질소원 함유 무기염 화학조성 배지 500 ml에 접종한 후 30oC에서 3일간 진탕 배양 (150 rpm)하면서 배양액의 흡광도(OD600)를 측정하고 균의 성장 을 specific growth rate로 표시하였다. 배양액의 pH는 전극이 장 치된 디지털 pH 미터(Model 7359, Istek, Korea)로 측정하였다.
시약
배지 성분, 당 분석에 필요한 aniline, phthalic acid, ninhydrine 및 표준 분자량 물질 등은 Sigma-Aldrich Inc. (USA)에서, 균체외 다당류의 정제 및 glucose 정량에 각기 필요한 에탄올과 황산 등 은 Merck 사(Thailand)에서, 산성 당(acidic sugar) 정량에 필요한 alcian blue 8GX는 Aldrich Chemical Co.에서, 균체외다당류 염색 용 PowerStain™ Silver Stain kit는 ELPIS Biotech (EDP1051, Korea)에서 구입하였다.
균체외다당류 정제
고속원심분리 (25,000×g, 15분)로 균 배양액으로부터 상등액을 얻은 후 상등액에 냉 무수에탄올을 3배(v/v) 첨가하고 4oC에서 하루 밤 정치하였다. 침전된 균체외다당류를 고속원심분리 (8,870×g, 15분)로 모은 후 DW 2 ml에 녹였다. 균체외다당류 액 을 -50oC에서 2일간 동결건조(Ilsin®, Korea)하였다(24, 35). 건조 된 균체외다당류를 다시 DW에 용해하고 2일간 15 L의 DW에서 투석(분자량 1,000 Da 이하는 통과)한 후, 균체외다당류를 냉 에 탄올로 재침전하고 동결 건조하였다. 이렇게 동결 건조한 균체외 다당류로 형태 관찰, 종이 크로마토그라피, 점도 측정 및 분자량 을 계측하였다.
균체외다당류의 구성 원소와 종이 크로마토그라피에 의한 구성 당 분석
균체외다당류의 구성 원소는 충북대학교 공동실험실습관에서 C, H, N, S, O 원소분석기(EA 1110, CE Instruments, Italy)로 분석하였다. 질소로 충전한 밀폐된 유리관에서 건조된 균체외다 당류를 6 N HCl로 16시간 동안 100oC에서 처리하여 가수분해물 을 얻었다. 왓트만 3 mm 종이(12×30 cm)에 DW에 녹인 가수분 해산물을 약 200 µg을 점적한 후 n-butanol:acetic acid:DW (3:1:1, v/v/v) 액에서 전개하였다. 비교표준물질로 5% (w/v) glucose, galactose, fructose, sucrose, rhamnose, glucuronic acid 용액을 사용했다. 100oC의 건열기에서 15분쯤 종이를 말린 뒤 당 발색 용액인 n-butanol/phthalic acid/aniline (26 ml:0.43 g:235 µl) 용 액에 잠시 담갔다가 꺼낸 후 건열기에서 말려 당 spot을 발색시켰 다. 균체외다당류의 분해산물에 아미노당(amino sugar)의 존재를 알기 위하여 새로 만든 0.2% (w/v) ninhydrine-ethanol액을 살포한 후 15분간 100oC에서 건조하였다(60). 아미노기의 표준물질로 5% (w/v) glycine과 glutamic acid액을 사용하였다.
균체외다당류의 분자량 측정
균체외다당류의 분자량을 SDS-polyacrylamide gel electrophoresis 로 측정하였다(15). 분자량 표준물질로 aprotinin (6.5 kDa), bovine α-lactalbumin (14.2 kDa), soybean trypsin inhibitor (20.1 kDa), bovine carbonic anhydrase (29 kDa), rabbit glyceraldehyde-3- phosphate dehydrogenase (36 kDa), chicken ovalbumin (45 kDa) (Sigma-Aldrich Co.)을 사용하였다. 전기영동 후 겔에 있는 균체 외다당류와 단백을 우선 50% 메탄올-12% 초산 용액에서 고정하 고 0.2% sodium thiosulfate 액에서의 전처리 과정을 거친 후 은 염색하였다(18). 균체외다당류와 단백질 표준분자의 밴드 간의 상대 이동비(Rf 값)에 근거하여 분자량을 구하였다.
균체외다당류 점도 측정과 형태 관찰
균체외다당류의 점도는 Ubbelohde 모세관 점도계(No. 30, Schott-Duran, Daihan, Korea)로 25oC±0.3의 수조에서 측정하였 다(60). DW와 균체외다당류 액 10 ml의 유속 시간(second)을 각 기 측정한 후 상대점도(η´rel = 균체외다당류 액의 유속 시간/
DW의 유속 시간)를 구하였다. 동결 건조한 균체외다당류를 디지
털 사진기, 광학 현미경, 주사전자현미경(S-2500C, Hitachi, Japan)으로 관찰하였다.
탄소원 정량
균 배양액의 잔여 탄소원 가운데 aldo-당은 DNS법으로, keto- 당은 Seliwanoff법으로 정량하였다(48). 배양액 1 ml과 DNS 3 ml (DNS 구성: 3,5-dinitrosalicylic acid 7.49 g, NaOH 13.98 g, Rochelle salt 216.1 g, sodium metabisulfite 5.68 g, phenol 5.73 ml, DW를 첨가하여 최종 부피가 1 L가 되게 함)을 혼합 후 100oC 수조에서 5분간 가열, 실온에서 냉각 후 잔여 glucose, galactose, lactose 같은 환원당이 3,5-dinitrosalicylic acid (DNS) 를 환원시켜 생성되는 주황색의 3-amino-5-nitrosalicylic acid의 농도를 OD550에서 측정하였다. Glucose, galactose, lactose로 작 성한 표준 곡선을 활용하여 당의 농도를 계량하였다. Fructose 농도는 Seliwanoff법으로 측정하였는데, 배양액 1 ml과 resorcinol [0.5% (w/v) in 3 N HCl] 5 ml을 잘 섞어 100oC 수조에서 1~2 분간 정치하면 형성되는 진홍의 furfural 복합체의 흡광도(OD480) 를 재었다. 농도를 달리한 fructose액으로 표준 곡선을 만들었다.
Glucose 상응 균체외다당류 및 산성 균체외다당류의 정량 Glucose 상응 균체외다당류는 phenol-sulfuric acid법으로 정량 하였다(31, 35). 스크류 캡 유리 시험관에 DW에 녹인 균체외다 당류 액 500 µl와 phenol 액(5%, v/v) 300 µl를 넣고 혼합하였다.
이에 sulfuric acid 1.5 ml을 천천히 첨가한 후 시험관 뚜껑을 닫 고 5초 동안 와동(vortexing)하였다. 그런 후 110oC 항온유조에서 15분간 가열한 반응액을 실온에서 식힌 후 반응액의 흡광도 (OD490)를 측정하였다. Glucose 용액으로 표준 곡선을 작성하여 glucose 상응 균체외다당류 정량에 참조하였다. 시료마다 세벌씩 분석하여 평균치를 구하였다. 산성 균체외다당류는 alcian blue 결합 분석법으로 정량하였다(24, 35, 44). 체외다당류 액 200 ml, acetic acid (7%, v/v)에 녹인 alcian blue 8GX 액(0.0015%) 400µl, acetic acid (7%) 400 µl로 구성된 반응액을 처음 5분 정 도는 완만히 가열, 진탕하고 나중에는 실온에서 2시간 진탕하였 다가 30분간 정치한 후 원심분리(9,800×g, 5분)로 얻은 상등액의 흡광도(OD600)를 측정하였다. 균체외다당류의 카복시 음전하와 반에스터 황산이 alcian blue의 양이온과 결합하여 불용성 침전 물을 형성하는데 이때 청색 상등액의 흡광도는 반응액 중 산성 체외다당류 량과 반비례한다. DW에 30분간 완만히 가열하여 녹 인 xanthan gum (0.5 mg/ml)을 4oC에 보관하였다가 표준 곡선 만드는데 사용하였다.
β-Galactosidase 활성 측정
Lactose 기본 배지에서 자란 MK1 균주의 β-galactosidase 활성 은 ONPG (o-nitrophenyl-β-D-galactoside)를 기질로 사용하여 Miller법으로 측정하였다(36). 균 배양액의 흡광도(OD600) 및 효 소 반응액의 흡광도(OD420, OD550)를 기록하여 수식에 따라 β- galactosidase unit를 구하였다. E. coli KCTC 25923를 β- galactosidase 활성 측정에 대조균으로 사용하였다.
결과 및 고찰 MK1 균주의 최적 성장
탄소원 및 질소원
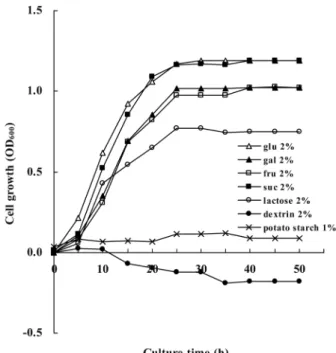
능이버섯의 무름병소에서 분리한 MK1 균주는 타가영양세균으 로 PDA (potato dextrose agar)에서 길렀을 때 균체외다당류를 많이 생산한다. 이 균주의 세균학적 특성은 보고되었지만(27), 생 리적 특성은 연구된 바 없다. 기본 무기염 배지에 각종의 탄소원 을 가하고 균을 길러보니 glucose, galactose, fructose, sucrose 첨 가 시는 균의 성장이 좋으나 lactose가 첨가되면 성장이 다소 저 조하였다(Fig. 1). 균의 성장 곡선과 배지 속의 잔여 탄소원 농도 는 서로 반비례하였다(자료 미제시). Glucose나 sucrose 배지에서 성장률(growth rate, 0.121 h-1)이 galactose나 fructose 배지 (0.095~0.092 h-1)보다 조금 좋았다. Potato starch나 dextrin를 함 유 배지에서는 균이 자라지 않아 탄소원으로 다당류를 사용하지 못함을 알 수 있다. Lactose 최소배지에서 24시간 자란 MK1 균 체에서는 β-galactosidase 활성이 전혀 검출되지 않았음으로(대조 군인 E. coli의 경우 600 U/ml 배양액), MK1의 lactose 분해 경 로는 lactose permease와 β-galactosidase에 의한 E. coli의 lactose 대사와는 다를 것으로 추정할 수 있다(3). 한편 β-galactosidase를 갖고 있지 않은 MK1 균주가 lactose 배지에서 자란 것이 이용 가능한 오염된 탄소원에 의한 것일 수도 있으나, 균의 성장에 반 비례하여 배양액의 lactose가 감소하는 것으로 보아 lactose가 균 의 성장에 이용되었다고 사료된다. 무기염 배지에 glucose량을
Fig. 1. Growth of MK1 cultured in basal minimal salt medium containing different carbon sources. Overnight grown MK1 strain in LB broth at 30oC was inoculated into basal minimal salt medium containing different carbon sources to give an initial OD600=0.05. Each value is an average of three measurements.
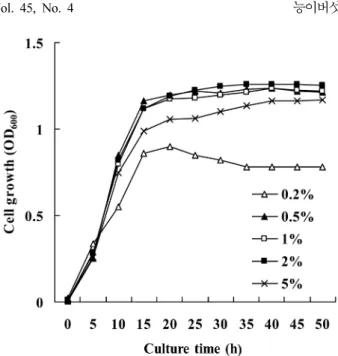
달리하여 균을 배양한 결과(Fig. 2), 0.5~2% glucose에서는 농도 의 구애 없이 균이 잘 생장하나 5%의 glucose에서는 MK1 생장 이 다소 줄었다. 이는 고농도 기질에 의한 성장 저해로써 사료되 는데 이러한 현상은 Azotobacter vinelandii MTCC 2460 (55), Bacillus polymyxa KS-1 (23), Methylomonas organophilum (8) 등에서 이미 보고된 바 있다. 질소원으로 NH4Cl, (NH4)2SO4, (NH4)2HPO4, yeast extract를 넣은 glucose (2.0%) 최소 배지에서 균을 배양하였을 때 yeast extract 첨가 배지에서 성장이 가장 빨 랐으며(0.230 h-1), (NH4)2HPO4에서는 0.17 h-1, (NH4)2SO4와 NH4Cl에서는 0.16 h-1로 다소 차이를 보였다. Yeast extract 첨가 배지에서 빠른 성장을 보이지만, yeast extract에는 유기 질소 이 외에 비타민 등 다른 성장 인자도 들어 있을 뿐만 아니라, 당을 정량할 때 감도가 떨어질 수 있으며, (NH4)2HPO4는 배지의 pH 유지에 영향을 줄 수 있어 구하기 쉬운 (NH4)2SO4를 질소원으 로 사용하는 것이 무난하다고 사료된다.
탄소/질소 비
Glucose (2.0%) 최소 배지에 (NH4)2SO4의 농도를 다양하게 조 정하여 탄소/질소의 비를 10~80:1로 조절한 후 균을 배양하였더 니 탄소/질소의 비가 10~40:1일 때는 MK1의 생장속도에 별 차 이가 없었으나(0.110 h-1), 탄소/질소의 비가 80일 때는 현저하게 줄었다(0.055 h-1).
pH, 온도 및 교반속도
MK1 생장에 대한 pH의 영향을 살펴본 결과, 지수기의 생장률 및 배양액의 생장수율 모두가 pH 7.0 에서 가장 좋았다. 일반적 으로 균체외다당류를 생산하는 세균은 중성 환경에서 최적 성장
을 보이는데 B. polymyxa (23), H. chejuensis (26), M.
organophilum (8)의 보고에서 비슷한 예를 찾을 수 있다. MK1은 37oC에서는 생장하지 않고 20oC 보다는 25~30oC에서 더 잘 자 랐다. 따라서 배양기의 온도 조절이 보다 용이한 30oC를 최적 성장 온도로 삼았다. MK1 균주는 절대 호기성 세균이라 교반속 도가 높을수록 성장도 좋고 균체외다당류의 생산성도 높을 것이 다. 배양액의 교반속도가 200 rpm일 때 균의 성장이 더 좋았다 (Table 1). 그러나 MK1의 균체외다당류 회수 할 때 거품이 지장 을 줄 수 있어 균 성장과 균체외다당류의 생산성 양면을 모두 고려하여 150 rpm이 적절하다고 본다. 해양 세균인 Zoogloea sp.
나 Lactobacillus sp.의 균체외다당류 생산성의 보고에서도 비슷한 점을 제기하였다(16, 42). 따라서 MK1 균주의 최적 배양 조건은 glucose 무기염 배지[2.0% glucose, 0.05~0.2% (NH4)2SO4, Na2HPO4 4.8 g, KH2PO4 4.4 g, MgSO4·7H2O 500 mg, FeCl3 50 mg, CaCl2 5 mg, l L DW, pH 7.0]를 사용하며 배양 온도 30oC, 교반속도 150 rpm 으로 정리된다.
MK1 균체외다당류의 특성
Glucose-무기염 배지에서 최적 조건을 맞추어 배양한 균 상등액 에서 냉 ethanol 침전으로 얻은 균체외다당류(EPSopt)의 구성 원소 는 탄소(37.1%), 수소(6.4%), 산소(49.3%), 질소(2.2%)였다. L.
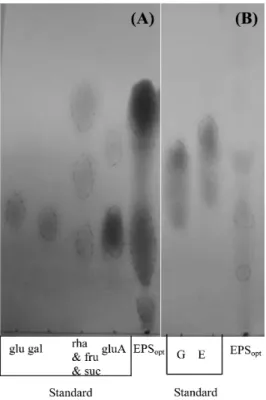
delbrueckii subsp. bulgaricus (21)나 Halomonas sp.의 균체외다당 류에는 황(sulfur)이 있는 것으로 보고되었는데(2, 32), MK1 균체 외다당류에서 황은 검출되지 않았다. EPSopt을 염산 가수분해하여 paper chromatography한 바 glucose, rhamnose, glucuronic acid 등 의 당과(Fig. 3A) glucose spot에서 amino기가 검출되어(Fig. 3B), EPSopt는 amino sugar와 acidic carbohydrate를 가진 이종다당류로 추정된다. EPSopt의 상세한 구조의 연구는 liquid chromatograph, NMR으로 진행 중이다. EPSopt의 분자량은 14.8~47.9 kDa으로 분 Fig. 2. Growth of MK1 at different concentrations of glucose.
Overnight grown MK1 strain in LB broth at 30oC was inoculated to minimal salt medium containing different glucose concentration to give an initial OD600=0.05. Each value is an average of three measurements.
Table 1. Specific growth rates of MK1 at various conditions parameter Specific growth rate (h-1) Remark
pH 6.0 0.138 30oC, 150 rpm
6.5 0.145 30oC, 150 rpm 7.0 0.191 30oC, 150 rpm 7.5 0.164 30oC, 150 rpm Temperature
(oC) 20 0.159 pH 7.0, 150 rpm
25 0.208 pH 7.0, 150 rpm
30 0.208 pH 7.0, 150 rpm
37 0.025 pH 7.0, 150 rpm
Agitation speed
(rpm) 100 0.143 30oC, pH 7.0 150 0.203 30oC, pH 7.0 200 0.270 30oC, pH 7.0 Minimal salt medium contained 2% glucose and 0.2% (NH4)2SO4 with C/N ratio of 10.
자량이 서로 다른 것이 혼재하고 있음을 알 수 있었다(Fig. 4). 분 자량이 다른 것들이 혼재하는 것은 균체외다당류가 효소의 작용으 로 분해된 결과로 추정된다(11, 21, 52). MK1 EPSopt의 분자량은 Halomonas maura (2), H. chejuensis (60), L. delbrueckii subsp.
bulgaricus (21), R. aquatillis (34), Streptococcus themophilus ST111 (54), S. themophilus Rs (13), S. themophilus LY03 (10) 등에서 보 고된 균체외다당류(1.5~5.0 mDa) 보다 훨씬 작았다.
pH에 따른 균체외다당류의 점성 및 형태 변화
Glucose-무기염 배지의 pH를 달리하여 배양한 균에서 얻은 균 체외다당류(EPSglu)의 물성이 상이하였다. pH 7.0의 배지에서 수 확된 균체외다당류(0.1 g/ml)의 상대 점도(η´rel)는 1.35, pH 6.0에 서 얻은 것은 1.23, pH 7.5의 것은 1.39로 배지의 pH가 높을수 록 균체외다당류의 점성이 높았다. Petry 등(43)은 균체외다당류 의 분자 크기가 클수록 점도가 높다고 하였는데, MK1 EPSopt (분 자량 14.8~47.9 kDa)의 점도가 H. chejuensis EPS-R (분자량 2.2 mDa)에 비하여 500~1,000배 낮은 것은 분자 크기의 현저한 차이 때문 일수도 있다(60). MK1 균체외다당류 물성의 차이를 광학현미경, 디지털 사진기 및 주사전자현미경으로도 알 수 있는 데(Fig. 5), 배지의 pH 6.0과 6.5에서 수확한 균체외다당류는 수 용성이며 고운 결정성 물성인데, pH 7.0과 7.5에서 얻은 균체외 다당류는 수용성이 줄고 점착성을 보였다. 이러한 결과에서 배지 Fig. 3. Paper chromatography of EPSopt-acid hydrolysates. (A)
Chromatograms for neutral and acidic sugar. Standard sugars: glucose (glu), galactose (gal), fructose (fru), sucrose (suc), rhamnose (rha), glucuronic acid (gluA). (B) Chromatograms fo amino sugar. Standard amino acids: glycine (G), glutamic acid (E).
Fig. 4. Molecular weight determination of EPS by SDS-PAGE. A mixture of protein markers (0.6µg) as well as EPS (300 µg) was resolved on 0.1 % SDS-15% PAGE. Protein size markers: aprotinin (6.5 kDa), bovine α-lactalbumin (14.2 kDa), soybean trypsin inhibitor (20.1 kDa) bovine carbonic anhydrase (29 kDa), rabbit glyceraldehyde- 3-phosphate dehydrogenase (36 kDa), chicken ovalbumin (45 kDa).
Fig. 5. Photos of freeze dried EPSs obtained from MK1 culture broth.
EPS was purified from culture broth of MK1 at log phase. Cells were grown in 2% glucose minimal medium whose initial pH was differently adjusted. EPS purification procedure was described in
‘Materials and Methods’.
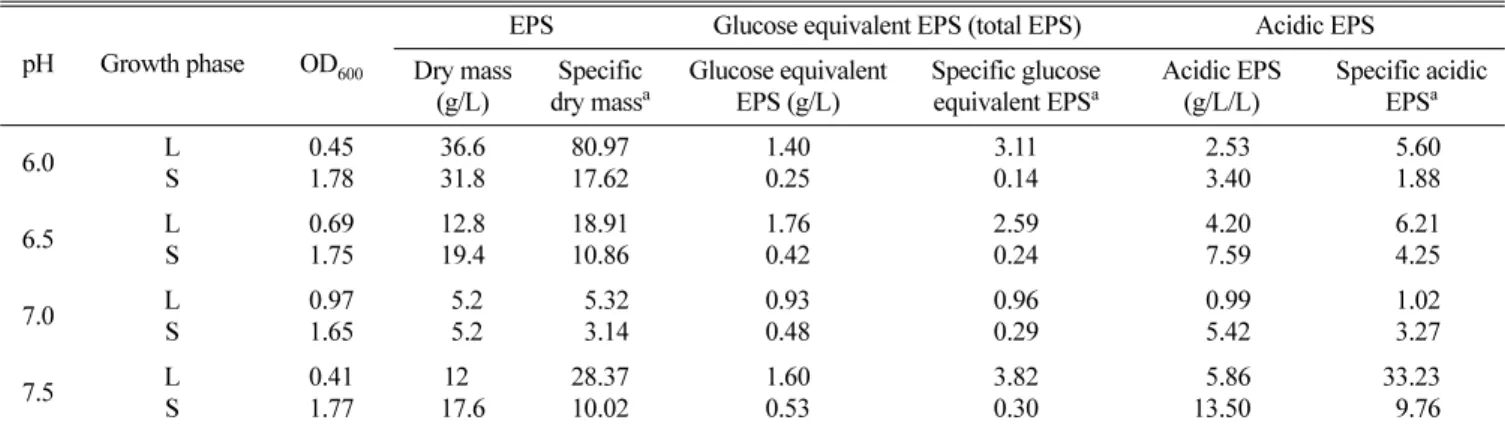
의 pH에 따라 생산된 균체외다당류의 물성이 달라짐을 추정할 수 있다. MK1의 균체외다당류는 세포에 느슨하게 결합하여 쉽 게 유리되는 유출 다당류(released polysaccharide, RPS)로(27), 세포에 단단히 결합된 협막 다당류(capsular polysaccharide, CPS)와는 달리 배양액으로부터 용이하게 얻을 수 있다(6, 33, 34). 미생물 균체외다당류에는 세포 보호 기능이 있어 성장에 부 적합한 환경이면 균체외다당류의 합성이 우세한데(5, 14, 30, 35, 53, 55), 그 이유는 세포벽 및 균체외다당류 합성대사가 isoprenoid glycosyl lipid 운반체에 대하여 서로 경쟁적이기 때문 인 것으로 생각된다(12, 38, 50). 이와는 달리 S. thermophilus는 호성장 환경에서 균체외다당류를 더 많이 생산한다는 보고가 있 다(9). 다양한 pH에서 MK1 균주를 배양하여 균체외다당류의 생 산성을 살펴 보면(Table 2), 배양액의 pH가 최적인 7.0보다 높거 나 낮으면 균체외다당류의 생산이 높아지는 추세이다. 이것을 근 거로 현재 균 생장 환경 변화에 따른 균체외다당류 생산성 변화 를 살피고 있다. L. delbrueckii subsp. bulgaricus의 phospho- polysaccharide (21, 22), Ulva rigida의 sulfur-polysaccharide (4) 나 식물에서 얻은 mannose 함유 다당류처럼(7, 59) 특정 성분을 지닌 균체외다당류는 면역자극활성이 현저하다는 보고가 있다.
우리 실험실에서도 MK1 균주의 균체외다당류가 쥐과 동물의 거 식세포를 자극하여 nitric oxide, TNF-α, IL-1β 생산성이 증가한 다고 보고한 바 있다(28). 이와 관련하여 MK1 균주의 균체외다 당류의 구조를 분석하여 신물질 인지 여부를 밝히는 연구를 수 행 중이다.
감사의 말
이 논문은 2008년도 충북대학교 학술연구지원사업의 연구비로 연구되었음.
참고문헌
1. Arena, A., T.L. Maugeri, B. Pavone, D. Iannello, C. Gugliandolo, and G. Bisignano. 2006. Antiviral and immunoregulatory effect of
a novel exopolysaccharide from a marine thermotolerant Bacillus licheniformis. Int. Immunopharm. 6, 8-13.
2. Arisa, S., A. del Moral, M.R. Ferrer, R. Tallon, E. Quesada, and V.
Bejar. 2003. Mauran, an exopolysaccharide produced by the halo- philic bacterium Halomonas maura, with a novel composition and interesting properties for biotechnology. Extremophiles 7, 319- 326.
3. Cadwell, D.R. 1995. Microbial physiology and metabolism, pp.102- 103. Wm. C. Brown Communications, Inc. Dubuque, Iowa, USA.
4. Castro, R., M.C. Piazzon, I. Zarra, J. Leiro, M. Noya, and J.
Lamas. 2006. Stimulation of turbot phagocytes by Ulva rigida C.
Agardh polysaccharides. Aquaculture 254, 9-20.
5. Cerning, T., C. Bouillanne, and M. Landon. 1992. Isolation and characterization of exopolysaccharides from slime-forming meso- philic lactic acid bacteria. J. Dairy Sci. 75, 692-699.
6. Chang, M.W., Y.S. Kang, J.W. Hong, J.D. Kim, and J.Y. Kong.
1995. Production conditions of two polysaccharides from marine bacterium Zoogloea sp. Kor. J. Biothechnol. Bioeng. 10, 518-524.
7. Chen, N.Y., T.H. Hsu, F.Y. Lin, H.H. Lai, and J.Y. Wu. 2006.
Effects on cytokine-stimulating activities of EPS from Tremella mesenterica with various carbon sources. Food Chem. 99, 92-97.
8. Choi, J.H., S.Y. Kim, D.K. Oh, and J.H. Kim. 1998. Optimization of culture conditions for production of a high viscosity polysac- charide, methylan, by Methylomonas organophilum from metha- nol. Kor. J. Appl. Microbiol. Biotechnol. 26, 244-249.
9. De Vuyst, L., F. Vanderveken, S. Van de Ven, and B. Degeest.
1998. Production and isolation of exopolysaccharides from Strep- tococcus thermophilus grown in milk medium and evidence for their growth-associated biosynthesis. J. Appl. Microbiol. 84, 1095- 1098.
10. Degeest, B. and L. De Vuyst. 1999. Indication that the nitrogen source influences both amount and size and modeling of the bac- terial growth and exopolysaccharide production in a complex medium. Appl. Environ. Microbiol. 65, 2863-2870.
11. Degeest, B., F. Mozzi, and L. De Vuyst. 2002. Effect of medium composition, temperature, and pH changes on exopolysaccharide yield and stability during Streptococcus thermophilus LY03 fer- mentations. Int. J. Food Microbiol. 79, 161-174.
12. Escalante, A., C. Axcher-Rodarte, M. Garcia-Garibay, and A.
Ferres. 1998. Enzymes involved in carbohydrate metabolism and their role on exopolysaccharide production in Streptococcus ther- mophilus. J. Appl. Microbiol. 84, 108-114.
Table 2. Effect of medium pH on production of EPSglu by MK1 strain
pH Growth phase OD600
EPS Glucose equivalent EPS (total EPS) Acidic EPS Dry mass
(g/L)
Specific dry massa
Glucose equivalent EPS (g/L)
Specific glucose equivalent EPSa
Acidic EPS (g/L/L)
Specific acidic EPSa
6.0 L
S
0.45 1.78
36.6 31.8
80.97 17.62
1.40 0.25
3.11 0.14
02.53 03.40
05.60 01.88
6.5 L
S
0.69 1.75
12.8 19.4
18.91 10.86
1.76 0.42
2.59 0.24
04.20 07.59
06.21 04.25
7.0 L
S
0.97 1.65
5.2 5.2
05.32 03.14
0.93 0.48
0.96 0.29
00.99 05.42
01.02 03.27
7.5 L
S
0.41 1.77
120 17.6
28.37 10.02
1.60 0.53
3.82 0.30
05.86 13.50
33.23 09.76 Minimal salts medium contained 2% glucose and 0.2% (NH4)2SO4 with C/N ratio of 10. L: log phase, S: stationary phase. aSpecific dry mass: Dry mass/ OD600, Specific glucose equivalent EPS: glucose equivalent EPS/ OD600, Specific acidic EPS: acidic EPS/ OD600.
13. Faber, E.J., P. Zoon, J.P. Kamerling, and J.F.G. Vliegenthart. 1998.
The exopolysaccharides produced by Streptococcus thermophilus Rs and Sts have the same repeating unit but differ in viscosity of their milk cultures. Carbohydr. Res. 310, 269-276.
14. Gancel, F. and G. Novel. 1994. Exopolysaccharide production by Streptococcus salivarius subsp. thermophilus cultures. 1. Condi- tions of production. J. Dairy Sci. 77, 685-688.
15. Gersten, D.M. 1996. Gel elecrtrophoresis of proteins: essential techniques, pp. 84-85, Wiley & Sons, Ltd., Chichester, UK.
16. Jang, J.H., S.K. Bae, B.J. Kim, S.D. Ha, and J.Y. Kang. 1998.
Effects of fermentation condition on the production of the usual polysaccharides from marine bacterium Zoogloea sp. Kor. J. Bio- technol. Bioeng. 13, 303-307.
17. Jeong, O.J., H.S. Yoon, and Y.K. Min. 2001. Aroma characteristics of Neungee (Sarcodon aspratus). Kor. J. Food Sci. Technol. 22, 307-312.
18. Kim, J.B. and J.H. Ahn. 1993. The modification of the silver stain method in sodium dodecyl sulfate-polyamide gels for detecting lipopolysaccharides. J. Kor. Soc. Microbiol. 28, 193-198.
19. Kim, J.W., B.S. Moon, Y.M. Park, N.H. Yoo, I.J. Ryoo, N.T.
Chinh, I.D. Yoo, and J.P. Kim. 2005. Structures and antioxidant activity of diketopiperazines isolated from the mushroom Sar- codon aspratus. J. Kor. Soc. Appl. Biol. Chem. 48, 93-97.
20. Kim, H.E., J.N. Park, W.S. Shin, J.S. Kim, S.C. Shin, and C.D.
Koo. 2000. Suggestions for developing the cultivation technology of mycorrihizal mushrooms, pp. 58-62. Proc. 2nd Int. Sym. New Horizon of Biosci. Forest Product Field, Cheongju, Korea.
21. Kitazawa, H., T. Harata, J. Uemura, T. Saito, T. Kaneko, and T.
Itoh. 1998. Phosphate group requirement for mitogenic activation of lymphocytes by an extracellular phosphopolysaccharide from Lactobacillus delbrueckii subsp. bulgaricus. Int. J. Food Micro- biol. 40, 169-175.
22. Kitazawa, H., Y. Ishii, J. Uemura, Y. Kawai, T. Saito, T. Kaneko, K. Noda, and T. Itoh. 2000. Argumentation of macrophage func- tions by an extracellular phosphopolysaccharide from Lactobacil- lus delbrueckii subsp. bulgaricus. Food Microbiol. 17, 109-118.
23. Kwon, G.S., B.D. Yoon, and H.K. Joo. 1995. Cultural conditions of exopolysaccahride KS-1 produced by Bacillus polymyxa KS-1.
Kor. J. Biotechnol. Bioeng. 10, 441-448.
24. Kumar, C.G., H.S. Joo, J.W. Choi, Y.M. Koo, and C.S Chang.
2004. Purification and characterization of an extracellular polysaccharide from haloalkalophilic Bacillus sp. I-450. Enzyme Microb. Technol. 34, 673-681.
25. Kumar, A.S., K. Mody, and B. Jha. 2007. Bacterial exopolysac- chrides - a perception. J. Basic Microbiol. 47, 103-117.
26. Lee, H.K., J. Chun, E.Y. Moon, S.H. Ko, D.S. Lee, H.S. Lee, and K.S. Bae. 2001. Hahella chejuensis gen. nov., sp. nov., and extra- cellular-polysaccharide-producing marine bacterium. Int. J. Sys.
Evol. Microbiol. 51, 661-666.
27. Lee, Y.N. and C.D. Koo. 2007. Identification of bacterial isolated from diseased Neungee mushroom, Sarcodon aspratus. J. Basic Microbiol. 47, 31-39.
28. Lee, Y.N., C.K. Lee, S.A. Im, and J.E. Ryu. Exopolysaccharide produced by MK1 strain isolated from diseased Neung-ee mush- room. Abstr. C005, p. 177, Proc. 2007 Int. Meet. Microbiol. Soc.
Kor.
29. Loaëc, M., R. Olier, and J. Guezennec. 1997. Uptake of lead, cad- mium, and zinc by a novel bacterial exopolysaccharide. Wat. Res.
31, 1171-1179.
30. Looijesteijn, P.J. and J. Hugenholtz. 1999. Uncoupling of growth
and exopolysaccharide production by Lactococcus lactis subsp.
cremoris NIZO B40 and optimization of its synthesis. J. Biosci.
Bioeng. 88, 178-182.
31. Masuko, T., A. Minami, N. Iwasaki, T. Majima, S.I. Nishimura, and Y.C. Lee. 2005. Carbohydrate analysis by a phenol-sulfuric and method in microplate format. Anal. Biochem. 339, 69-72.
32. Mata, J.A., V. Bejar, I. Llams, S. Aris, P. Bressollier, R. Tallon, M.C. Uradaci, and E. Quesada. 2006. Exopolysaccharides pro- duced by the recently described halophilic bacteria Halomonas ventosae and Halomonas anticariensis. Res. Microbiol. 157, 827- 835.
33. Matsuyama, H., T. Kamesaki, R. Sasaki, H. Minami, and I.
Yumato. 2003. Production of two types of exopolysaccaride by Novosphingobium rosa. J. Biosci. Bioeng. 95, 152-156.
34. Matsuyama, H., R. Sasaki, K. Kawasaki, and I. Yumoto. 1999.
Production of a novel exopolysaccharide by Rahnella aquatilis. J.
Biosci. Bioeng. 87, 180-183.
35. McKellar, R.C., J. van Geest, and W. Cui. 2003. Influence of cul- ture and environmental conditions on the composition of exopolysaccharide produced by Agrobacterium radiobacter. Food Hydrocolloides 17, 429-437.
36. Miller, J.H. 1972. Experiments in Molecular Genetics. pp. 352- 355. Cold Spring Harbor Lab. Press, New York, USA.
37. Mizuno, M., Y. Shiomi, K. Minato, S. Kawakami, H. Ashida, and H. Tsuchida. 2000. Fucogalactan isolated from Sarcodon aspratus elicits release of tumor necrosis factor-a and nitric oxide from murine macrophages. Immunopham. 46, 113-121.
38. Mozzi, F., G. Rollan, G.S. de Glori, and G.F. de Valdez. 2001.
Effect of galactose and glucose on the exopolysaccharide produc- tion and the activities of biosynthetic enzymes in Lactobacillus casei CRL 87. J. Appl. Microbiol. 91, 160-167.
39. Nishimura-Uemura, J., H. Kitazawa, Y. Kawai, T. Itoh, M. Oda, and T. Saito. 2003. Functional alteration of murine macrophages stimulated with extracellular polysaccharides from Lactobacillus delbrueckii subsp. bulgaricus OLL1073R-1. Food Microbiol. 20, 267-273.
40. Ozdemir, G., N. Ceyhan, and E. Manav. 2005. Utilization of an exopolysaccharide produced by Chryseomonas luteola TEM05 in alginate beads for adsorption of cadmium and cobalt ions. Biores.
Technol. 96, 1677-1682.
41. Park, B.S., C.D. Koo, K.H. Ka, and Y.N. Lee. 2008. Effect of chi- tosan acetate on bacteria occurring on Neungee mushrooms, Sar- codon aspratus. Mycobiology 36, 249-254.
42. Petry, S., S. Furlan, M.J. Crepeau, J. Cerning, and M. Des- mazeaud. 2000. Factors affecting exocellular polysaccharide pro- duction by Lactobacillus delbrueckii subsp. bulgaricus grown in a chemically defined medium. Appl. Environ. Microbiol. 66, 3427- 3431.
43. Petry, S., S. Furlan, E. Waghorne, L. Saulnier, J. Cerning, and E.
Maguin. 2003. Comparison of the thickening properties of four Lactobacillus delbrueckii subsp. bulgaricus strains and physico- chemical characterization of their exopolysaccharides. FEMS Microbiol. Lett. 221, 285-291.
44. Ramus, J. 1977. Alcian blue: A quantitative aqueous assay for algal and sulfated polysaccharides. J. Phycol. 13, 345-348.
45. Silva, F.R., A.L. Vettore, E.L. Kemper, A. Leite, and P. Arruda.
2001. Fastidian gum: the Xylella fastidiosa exopolysaccharide possibly involved in bacterial pathogenicity. FEMS Microbiol.
Lett. 203, 165-171.
46. Shin, H.G., Y.M. Choi, H.K. Kim, Y.C. Ryu, S.H. Lee, and B.C.
Kim. 2007. Tenderization and fragmentation of myofibrillar pro- teins in bovine longissimus dorsi muscle using proteolytic extract from Sarcodon aspratus. Food Sci. Technol. 21, 1-7.
47. Song, J.H., H.S. Lee, J.K. Hwang, J.W. Han, J.G. Ro, D.H. Keum, and K.M. Park. 2003. Physiological activity of Sarcodon aspratus extracts. Kor. J. Food Sci. Ani. Resour. 23, 172-179.
48. Stenesh, J. 1984. Experimental biochemistry, pp. 237-240. Allyn and Bacon Inc., Boston, MA, USA.
49. Sutherland, I.W. 1998. Novel and established applications of microbial polysaccharides. Tibtech. 16, 41-46.
50. Sutherland, l.W. 2001. The biofilm matrix an immobilized but dynamic microbial environment. Trends Microbiol. 9, 222-227.
51. Takei, T., M. Yoshida, M. Ohnishi-Kameyama, and M. Kobori.
2005. Ergosterol peroxide, and apoptosis-inducing component iso- lated from Sarcodon aspratus (Berk.) S. Ito. Biosci. Biotechnol.
Biochem. 69, 212-215.
52. Tallon, R., P. Bressollier, and M.C. Uraci. 2003. Isolation and char- acterization of two exopolysaccharides produced by Lactobacillus plantarum EP56. Res. Microbiol. 154, 705-712.
53. Toda, K., Y. Gotoh, T. Asakura, I. Yabe, and H. Furuse. 2000.
Fluid viscosity of Aureobasidium pullulans cultures obtained at two different initial pH values. J. Biosci. Bioeng. 89, 258-261.
54. Vaningelgem, F., R.V. Meulen, M. Zamfir, J. Adriany, A.P. Laws, and L. de Vuyst. 2004. Streptococcus thermophilus ST111 pro- duces a stable high-molecular-mass exopolysaccharides in milk- based medium. Int. Dairy J. 14, 857-864.
55. Vermani, M.V., S.M. Kelkar, and M.Y. Kamat. 1995. Production and optimization of certain growth parameters for an exopolysac- charide from Azotobacter vinelandii MTCC 2460 isolated from a plant rhizosphere. J. Ferment. Bioeng. 80, 599-602.
56. Wang, H., X. Jiang, H. Mu, X. Liang, and H. Guan. 2007. Struc- ture and protective effect of exopolysaccharide from Pantoea agglomerans strain KFS-9 against UV radiation. Microbiol. Res.
162, 124-129.
57. Welman, A.D. and I.S. Maddox. 2003. Exopolysaccharides from lactic acid bacteria: perspectives and challenges. Trends Biotech- nol. 21, 269-274.
58. Yang, J.H., J.S. Eun, and J.D. Her. 1989. Studies on proteolytic enzyme preparation using Sarcodon aspratus extracts. J. Kor.
Pharm. Sci. 19, 203-212.
59. Yang, T., M. Jia, J. Meng, H. Wu, and Q. Mei. 2006. Immunomod- ulatory activity of polysaccharide isolated from Angelica sinensis.
Int. J. Biol. Macromol. 39, 179-184.
60. Yim, J.H., S.J. Kim, S.H. An, and H.K. Lee. 2004. Physicochem- ical and rheological properties of a novel emulsifier, EPS-R, pro- duced by the marine bacterium Hahella chejuensis. Biotechnol Bioprocess Eng. 9, 405-413.
61. Yoo, S.H., E.J. Yoon, J. Cha, and H.G. Lee. 2004. Antitumor activ- ity of levan polysaccharides from selected microorganisms. Int. J.
Biol. Macromol. 34, 37-41.
62. Zhang, M., S.W. Cui, P.C. Cheung, and Q. Wang. 2007. Antitumor polysaccharides from mushrooms: A review on their isolation pro- cess, structural characteristics and antitumor activity. Food Sci.
Technol. 18, 4-19.
63. Zirk, N.M., S.F. Hashmi, and H.K. Ziegler. 1999. The polysaccha- ride portion of lipopolysaccharide regulates antigen-specific T- cell activation via effects on macrophage-mediated antigen pro- cessing. Infect. Immun. 67, 319-326.
(Received August 17, 2009/Accepted October 30, 2009)
ABSTRACT : Optimal Culture Conditions for MK1 Strain Isolated from Soft-Rotten Tissue of Neungee Mushroom (Sarcodon aspratus) and the Physico-Chemical Properties of the Purified Exopolysaccharide of MK1
Jeong Eun Ryu and Young Nam Lee* (Department of Microbiology, Chungbuk National Uni- versity, Cheongju 363-761, Republic of Korea)
MK1 strain, an obligate aerobic heterotrophic bacterium isolated from the rotten tissue of Neungee mushroom (Sarcodon aspratus), produces a copious amount of exopolysaccharide (EPS), which could evoke macrophage activation. Investigations on optimal culture conditions of MK1 and physical properties of MK1 EPS were made. Glucose, galactose, fructose, and sucrose supported well growth of MK1, but potato starch and dextrin did not. However, lactose seemed to be a less favorable carbon source. Optimal growth of MK1 was obtained at pH 7.0, 30oC, and 200 rpm with 2% glucose, and 0.2~0.05% (NH4)2SO4. EPSopt obtained from an optimal growth condition constituted of carbon (37.1%), nitrogen (2.2%), oxygen (49.3%), and hydrogen (6.4%), but no sulfur. Paper chrogromatogram of the acid-hydrolysate of EPSopt suggested that MK1 EPS seemed to be hetropolysaccharide composed of a few number of monosaccharides including amino- and acidic-sugars. Its molecular mass determined by SDS-polyacrylamide gel electrophoresis varied from 14.8 to 47.9 kDa. Physical properties of EPSglu obtained from cell grown in glucose medium, such as relative viscosity (rel) and crystalline morphology were rather affected by pH of the growth medium. Relative viscosity (rel) of exopolysaccaride (0.1 g/ml) harvested from cells grown at medium pH ranging from 6.0 and 7.5 was 1.23 and 1.39, respectively. The freeze-dried exopolysaccharide obtained at low pH (6.0 and 6.5) was fine crystaloid and water-soluble, whereas those obtained at high pH (7.0 and 7.5) was rather gluey and less water-soluble.