24
「The Sea」 Journal of the Korean Society of Oceanography Vol. 21, No. 1, pp. 24─ 35, February 2016
http://dx.doi.org/10.7850/jkso.2016.21.1.24 Free Access
울산만 내측과 외측에서 계절적 환경요인의 변화에 의한 식물플랑크톤 성장 및 분포
이민지1,4 · 김동선2 · 김영옥1 · 손문호1,3 · 문창호4 · 백승호1,*
1한국해양과학기술원 남해특성연구센터
2한국해양과학기술원 환경기반연구센터
3국립수산과학원
4부경대학교 해양학과
Seasonal Phytoplankton Growth and Distribution Pattern by Environmental Factor Changes in Inner and Outer Bay of Ulsan, Korea
M
IN-J
IL
EE1,4, D
ONGSEONK
IM2, Y
OUNGO
KK
IM1, M
OONHOS
OHN1,3, C
HANG-H
OM
OON4ANDS
EUNGH
OB
AEK1,*
1South Sea Institute, KIOST, Geoje 656-830, Korea
2Marine Chemistry & Geochemistry Research Center, KIOST, Ansan 425-600, Korea
3Fishery and Ocean Information Division, NFRDI, Busan 619-705, Korea
4Departmaent of Oceanography, Pukyong National University, Busan 608-737, Korea
2014년 울산만 내, 외측 해역에서 해양환경의 계절 변화가 식물플랑크톤 군집의 공간 분포에 미치는 영향을 파악 하였다. 울산만을 태화강 하구에 위치한 내측과 외양 영향을 강하게 받는 외측으로 나누어, 내측과 외측의 환경 차 이를 t-test로 검증하였다. 수온은 동계(t = -5.833, p < 0.01)와 추계(p > 0.05)에는 외측이 높았고, 춘계(t = 4.247, p< 0.01)와 하계(t = 2.876, p < 0.05)에는 내측이 높았다. 염분은 추계를 제외한 모든 계절에 내측에서 외측보다 유 의하게 낮았다(p < 0.01). 동계에는 수층 혼합에 의해 전 수층에서 영양염 농도가 높았고, 하계에는 담수 유입으로 내측 표층에서 현저하게 높았다. 크기 분획된 Chl. a 양은 20 µm보다 작은 크기의 nano, pico식물플랑크톤의 양이 많았다. 극미소 식물플랑크톤은 영양염 농도가 낮은 외양 환경에서 적응력이 높은데 이러한 결과는 조사해역이 외 양의 영향을 강하게 받고 있음을 시사한다. 춘계와 하계에는 내측 정점을 중심으로 유글레나류 Eutreptiella gymnastica의 밀도가 특이적으로 높았다. 이는 강우 후 많은 담수가 유입되어 저염분 환경이 조성되고 DIN (nitrate+nitrite, ammonium)이 대량 공급되었기 때문이다. 하계에는 태화강으로부터 다량의 담수가 공급되어 담수미세 남조류 Oscillatoria sp., Microcystis sp.가 내측 정점(1-5)을 중심으로 높은 밀도로 출현하였다. 추계와 동계에는 규조류와 은편모류가 우점하였다. 높은 현존량을 보인 중심 규조류Chaetoceros 속은 성층 약화로 저층에서 공급되는 영양염 류의 영향을 받아 빠르게 성장하였고, 은편모조류는 다른 식물플랑크톤의 상대적인 성장 둔화로 기회적으로 우점하 였다. 결과적으로 울산만은 강우기인 춘계와 하계에 태화강의 영향을 강하게 받아 내측 정점을 중심으로 저염분, 고 영양염 환경이 유지되었고, 추계와 동계에는 만 외측 외양수의 영향을 크게 받았으며, 식물플랑크톤 군집은 크게 이 두가지 환경요인에 의해 계절적으로 제어되는 것으로 판단되었다.
To assess the relationship between environmental factors and seasonal phytoplankton community structure, we investigated abiotic and biotic factors in Ulsan Bay, Korea. We divided the bay into two areas based on geo- graphical characteristics and compared the difference in each factor between inner and outer bay with t-test sta- tistics. As a result, temperature in the outer bay was higher than that of the inner bay during winter (t = -5.833, p < 0.01) and autumn (p > 0.05). However, opposite trend was observed during spring (t = 4.247, p < 0.01) and summer (t = 2.876, p < 0.05). Salinity was significantly lower in the inner bay than in the outer bay in winter, spring, and
Received December 14, 2015; Revised February 4, 2016; Accepted February 14, 2016
*Corresponding author: [email protected]
summer (p < 0.01). However, the salinity was not significantly different between the inner and the outer bay in the autumn (p > 0.05). In particular, high nutrient concentration was observed in most stations during winter season due to vertical well mixing. The nutrient concentration was significantly higher in surface layers of inner bay after rainfall, particularly in the summer. The relative contribution (approximately 70%) of < 20 µm (nano and pico) size phytoplankton was increased in all seasons with continuously low nutrients from the offshore water due to their adaption to low nutrient without other large competitors. Interestingly, high population of Eutreptiella gymnastica was kept in the inner bay during the spring and summer associated with high DIN (nitrate+nitrite, ammonium) after river discharge following rainfall, suggesting that DIN supply might have triggered the increase of Eutrep- tiella gymnastica population. In addition, high density of freshwater species Oscillatoria sp. and Microcystis sp.
were found in several stations of the inner bay that were provided with large amounts of freshwater from the Tae-wha River. Diatom and cryptophyta species were found to be dominant species in the autumn and winter.
Of these, centric diatom Chaetoceros genus was occupied in the outer bay in the autumn. Cryptophyta species known as opportunistic micro-algae were found to have high biomass without competitors in the inner bay. Our results demonstrated that Ulsan Bay was strongly affected by freshwater from Tae-wha River during the rainy season and by the surface warm water current from the offshore of the bay during dry season. These two exter- nal factors might play important roles in regulating the seasonal phytoplankton community structures.
Key words: Seasonal trends, Water temperature, Salinity, Nutrients, Phytoplankton, Ulsan Bay
서 론
식물플랑크톤은 일차생산자로서 먹이흐름의 출발점에 있고, 그 들의 성장은 수온, 염분, 광량, 광주기, 영양염류 등과 같은 무생물 학적 환경요인과 더불어 종간 경쟁, 상위영양단계 생물의 포식, 박 테리아의 감염 등과 같은 생물학적 요인에 의하여 크게 영향을 받 는다(Thompson et al., 2008; Guinder et al., 2013). 또한 이들 환경 요인 중 영양염류의 공급과 제한은 식물플랑크톤 생물량의 증감과 종구성에 중요한 영향을 미칠 수 있고, 수계 내 영양염류의 분포 양상에 따라서 식물플랑크톤의 계절적 천이양상을 부분적으로 설명 가능할 수 있다(Anderson et al., 2002). 일반적으로 온난해역에서 식물플랑크톤의 대발생은 춘계와 추계에 일어나는 것으로 알려져 있다. 춘계의 대발생은 동계의 수층혼합으로 기인된 영양염류가 수계 내에서 지속적인 공급과 함께, 광량 및 일주기 증가가 중요 하게 작용하고, 추계의 발생은 하계에 고갈된 영양염류가 성층붕 괴와 더불어 유광층으로 공급되는 것이 중요하게 작용된다(Riley, 1942; Cushing, 1989). 특히 하계에는 수온증가와 더불어 계절적 수온약층으로 인하여 저층의 풍부한 영양염류가 유광층에 충분하게 공급되지 않아 식물플랑크톤의 성장에 부정적인 영향을 미칠 수 있다. 하지만, 우리나라는 몬순기후의 영향으로 6-7월의 장마철에 강우가 집중되고, 8-9월에는 산발적으로 접근하는 태풍의 영향으로 하계에도 불구하고, 육상기원의 영양염류가 연안해역으로 대량 유 입되어 식물플랑크톤 대발생이 빈번하게 발생하고 있다.
연안내만에서 영양염류의 공급원으로는 육상기원의 담수유입과 수계내에 있는 생물 증식에 의하여 생성되는 것과 더불어 분해과 정에서 기인하는 것으로 크게 구분할 수 있다. Dortch and Whitledge (1992)에 의하면, 해역의 영양염류 절대값 농도가 N < 1 μM, P < 0.2 μM, Si < 2 μM일 때 식물플랑크톤의 성장에 대한 영양염이 제한된다고 보고하였다. 따라서 특정해역에서 영양염류의 절대농도의 계절적 분포 특성을 파악하는 것은 식물플랑크톤의 계절적 군집조성 및 종천이 현상을 이해하는데 중요하다.
울산만은 한반도의 동남쪽연안에 위치하며 동해 쪽으로 열려있는
반폐쇄적인 만으로 해안선 전체 길이가 58 km, 항내수면적은 116 km2, 수심은 3~27 m로 상당히 규모가 크며, 옛날부터 어업이 성하여 동 해안의 장생포항과 방어진항을 중심으로 동해남부지역의 수산업 중심지로 발달되어 왔다. 하지만 1962년 특정공업지역으로 지정된 이후 중공업, 기계, 석유화학, 비철금속 산업 등이 급속도로 발달 되어 현재 울산만을 따라 울산석유화학단지와 온산공단과 같은 큰 산업단지가 다수 위치하고 있다. 특히 울산시과 온산시 인근 지역 에서 배출되는 막대한 양의 생활하수 및 공업폐수가 유입되어, 만 내 부영양화를 촉진시키는 요인으로 분석된다. 2000년부터 특별관 리해역으로 지정되어 환경개선을 위해 많은 국가예산이 투입되어 태화강 및 만 내측의 수질이 단계적으로 개선되고 있고, 연어가 회 귀하는 등 부분적으로 생태계 복원 효과가 나타나고 있다. 하지만, 외항강 하구의 수질개선 정도는 크지 않을 뿐만 아니라 유해화학 물질과 중금속에 의한 퇴적물 오염이 관측되고 있다(Koh et al., 2002). 울산만은 남동쪽으로는 쿠로시오의 지류인 대마난류의 영 향을 받아 외해수가 유입되고, 북쪽의 태화강과 서쪽 아래에는 고 사천과 외항강으로부터 담수가 유입된다. 집중강우 시에는 태화강을 중심으로 유입되는 많은 양의 담수는 만 전체에 걸쳐 저염분화와 더불어 영양염류의 공급을 촉진할 수 있다. 이와 같은 경로를 통 하여 유입된 다량의 영양염류로 인해 울산만은 식물플랑크톤의 대 증식이 빈번하게 발생할 수 있는 잠재적인 해역으로 판단된다. 울 산만 해역에서 식물플랑크톤의 군집조성 및 생체량의 계절적 변동은 Joo et al. (2011)과 더불어 Kwon and Kang (2013)의 연구로 국한 되어 있고, 영양염 분포 특성과 식물플랑크톤의 관계에 대한 연구는 거의 없는 실정이다. 따라서 본 연구는 2014년 울산만의 내측과 외측해역에서 계절에 따른 환경요인의 변화가 식물플랑크톤의 군 집구조에 미치는 영향을 파악하고자 하였다.
재료 및 방법
울산만의 내측 해양환경은 태화강과 함께 울산시, 온산시 인근 지역에서 배출되는 막대한 양의 생활하수 및 공업폐수가 유입되어,
26 이민지 · 김동선 · 김영옥 · 손문호 · 문창호 · 백승호
만 내측의 부영양화를 촉진시키는 요인으로 작용될 수 있다(Cloern, 2001; Park et al., 2010). 반면, 만 외측환경은 남동쪽으로는 쿠로 시오의 지류인 대마난류의 영향을 받아 외해수가 유입되어, 상대 적으로 빈영양상태가 유지될 수 있으나, 내측의 높은 영양염류가 외측의 빈영양수괴와 혼합 희석되어 일정의 영양염 농도로 유지될 수 있는 해역으로 판단된다. 따라서 본 연구에서는 이와 같은 울 산만의 지형 및 물리학적 특성에 기반하여 변화될 환경요인의 차 이에 따른 식물플랑크톤의 계절적·공간적 차이의 특성을 명확하게 구분하기 위하여 울산만 내측(1, 2, 3, 4, 5, 6, 10)과 외측(7, 8, 9, 11, 12, 13)을 나누어 평가하였다.
현장조사는 울산만 13개의 정점에서 2014년 동계(2월 12일), 춘 계(5월 7일), 하계(8월 6일), 추계(11월 5일)에 걸쳐 수행되었고, 표 층수는 버켓으로 저층수는 니스킨 채수기(5L PVC Niskin채수기;
General Oceanics, Miami, FL, USA)를 이용하여 각각 채수하였다 (Fig. 1). 수온과 염분은 CTD (Ocean Seven 319; Idronaut Co., Brugherio, Italy)를 이용하여 측정하였다. 영양염 분석을 위해 표층수와 저층수를 GF/F(25 mm, pore size 0.45 µm; Whatman, Middlesex, U.K.)필터를 이용하여 여과한 후 PE병에 넣고 HgCl2를 첨가한 후 냉동 보관하였다(Kattner, 1999). 냉동 보관한 영양염은 분석 전 해 동하여 암모니아, 질산염, 아질산염, 인산염, 규산염을 Parsons et
al. (1984)의 분석법에 따라 영영염 자동분석기(Autoanalyzer QuikChem 8000; Lachat Instruments, Loveland, CO, USA)를 이 용하여 분석하였고, 영양염 농도는 표준시약인 Brine solution (CSK Standard Solutions; Wako Pure Chemical Industries, Osaka, Japan)을 이용하여 보정하였다.
Chlorophyll-a (Chl. a) 는 현장에서 크기분획을 위해 20 µm와 3 µm polycarbonate filter로 여과한 후, 크기 별 Chl. a 측정을 위해 Whatman GF/F glass fiber filters (a 47-mm diameter; pore size 0.45 µm)를 이용하여 필터하였다(100~300 mL, 계절 및 생물 출현 량에 따라 차이를 둠). 필터는 15 ml 튜브에 넣어 분석 전까지 -20 °C 에서 냉동 보관하였으며, 이후 냉동 보관한 여과지를 90% Acetone에 넣어 냉암소에서 24시간 동안 색소를 추출한 후 형광측정기(Turner BioSystems, Sunnyvale, CA, USA)를 이용하여 분석하였다(Parsons et al., 1984).
식물플랑크톤 종조성과 정량분석을 하기 위해 500 ml의 표층수를 루골을 이용하여 최종농도0.5%가 되게 고정하였다. 이후 500 ml의 샘플을 50 ml로 농축시킨 후 Sedgewick-Rafter counting chamber에 샘플을 100~300 µl분주하여 200배 혹은 400배 배율로 동정 및 계 수하였다.
내측와 외측 환경 및 생물 요소의 차이를 확인하기 위해 t-test를 실 Fig. 1. Location of sampling stations in Ulsan Bay, Korea.
시하였으며, p < 0.05 일 때 통계적으로 유의하다고 판단하였다.
생물학적 요소와 무생물학적 요소의 계절별 정점별 차이는 One- way ANOVA를 통해 평가하였고, SPSS version 17.0 (SPSS, Inc., Chicago, IL, USA)를 이용하여 수행하였다. 또한, 식물플랑크톤과 환경인자간의 특성을 파악하기 위해 XLSTAT2012(AddinSoftTM)를 이용하여 주성분분석(PCA, principal component analysis)을 실시 하였다.
결 과
환경요인의 변화
2014년 울산만의 표층과 저층의 수온 및 염분 변화를 Table 1에 나 타내었다. 동계 표층 수온은 9.2~13.4 °C, 저층의 수온은 10.6~13.4 °C 로 큰 차이를 보이지 않는 것으로 보아, 수층의 혼합이 활발하게 이루어졌음을 알 수 있다. 반면, 상대적으로 내만 해역의 수온은 외만 해역보다 낮은 양상을 보였다. 춘계의 표층 수온은 13.6~15.0 °C로 변화하여 정점간의 큰 차이를 보이지 않았다. 저층에서는 12.4~14.1 °C 로 변화하였고, 표층과 저층에서 약 1 °C 정도의 온도차를 보였다.
하계에는 모든 정점에서 표층과 저층사이에서 성층이 뚜렷하게 형 성되었고, 각 정점의 수심에 따라서 표층과 저층의 온도차가 4.3~8.1 °C로 크게 나타났다. 표층수온의 변화는 19.1~24.5 °C로 저 층의 수온변화(11.7~20.2 °C)보다 적은 편차를 보였다. 추계 정점 간 수온변화 특성은 표층 18.4~19.3 °C로, 내측과 외측간의 차이는 보이지 않았다. 저층의 경우 정점 10까지는 17.1~19.3 °C로 표층과 거의 유사하였으며, 정점 11에서부터 점차적으로 하강하여 정점 13에서 12.1 °C로 가장 낮게 관찰되었다. 즉 내측 정점에서 외측 정점으로 갈수록 수온의 성층화 현상이 강하게 나타났다.
정점별 표층 염분은 동계 31.2~34.5 psu로, 춘계의 29.4~34.4 psu와 유사하게 나타났고, 특히 내측으로 갈수록 염분이 낮아지는 특징을 보였다. 하계의 표층 염분은 13.5~31.9 psu의 변동을 보였 고, 표층과 저층의 차이가 크게 나타났다. 그 중 정점 1에서 표층 (13.5 psu)과 저층(27.9 psu)의 차이가 14.4 psu로 컸다. 추계 표층 염분은 26.0~33.2 psu로 변화하였고, 정점 1에서 26.0 psu으로 현 저하게 낮은 값을 보였고, 그 외 정점에서는 약 32 psu전 후로 큰
변동을 보이지 않았다. 특히 저층은 32 psu이상으로 계절별·정점 별 큰 변동을 보이지 않았다.
영양염농도 변화
영양염 농도의 정점간 계절별 수평적 분포양상을 Fig. 2에 나타 내었다. 질산염 + 아질산염 농도는 3.0 µM~93.0 µM로 큰 변화폭을 보였고, 표층 평균 농도는 22.3 µM로(동계: 15.6 µM, 춘계: 18.2 µM, 하계: 42.8 µM, 추계: 12.5 µM), 저층 평균농도 10.4 µM (동계: 10.8 µM, 춘계: 7.8 µM, 하계: 13.9 µM, 추계: 9.2 µM)보다 높았다. 계 절적으로 하계에 매우 높게 나타났으며, 특히 강하구에 인접한 정 점에서 상대적으로 높게 나타나는 경향을 관찰하였다. 암모니아농 도는 0.01 µM~100.1 µM로 변동하였고, 모든 정점에서 표층과 저 층의 암모니아 평균농도는 하계(17.6 ± 22.4 µM) > 동계(10.0 ± 13.5 µM) > 추계(8.6 ± 19.9 µM) > 춘계(5.9 ± 9.3 µM)순으로 높게 나타났다. 인산염은 0.1 µM에서 5.6 µM의 농도범위를 보였고, 다 른 영양염과 유사하게 정점 1~4사이에서 높게 나타나는 경향을 관 찰하였다. 인산염의 평균농도는 표층에서 하계와 동계에 각각 1.9 µM과 1.3 µM 로 나타났고, 춘계와 추계에는 0.9 µM로 상대적으로 낮게 관찰되었다. 각 계절 표층과 저층 인산염의 차이는 평균 0.44 µM로 나타났고, 특히 하계의 표층과 저층의 차이가 컸다. 규산염 농도는 표층과 저층에서 7.0 µM~86.6 µM로 변동하였고, 전 계절 표층 평 균 농도는 22.9 µM로, 저층 농도15.4 µM 보다 약간 높게 관찰되 었다. 표층과 저층의 규산염농도의 계절별 평균은 하계(표층: 39.6 µM;
저층: 19.3 µM) > 추계(표층: 18.8 µM; 저층: 16.0 µM) > 춘계(표층:
18.3 µM; 저층: 12.9 µM) > 동계(표층: 15.0 µM; 저층: 13.2 µM) 순으로 높게 관찰되었다.
엽록소(Chl. a) 농도
Chl. a는 총량(Total Chl. a)과 크기분획(Pico: < 3 µm, Nano:
3~20 µm, Micro: > 20 µm)된 농도로 구분하여 평가하였고, 그 결 과를 Fig. 3에 나타내었다. 총 Chl. a량은 추계(0.7~4.1 µg L–1) >
춘계(0.4~1.8 µg L-1) > 하계(0.6~2.6 µg L-1) > 동계(0.4~1.0 µg L-1) 순으로 높게 나타났다. 상대적으로 총 Chl. a량은 추계 외측 정점에서 높은 농도로 관찰되었고, 특히 정점 12에서 4.1 µg L-1로 최고치에
Table 1. Seasonal ranges of temperature, salinity, nutrients (silicate, phosphate, and nitrate), chlorophyll-a, and the abundance of phytoplankton (diatoms, nanoplankton, dinoflagellates) in Ulsan Bay, Korea. Data represent as mean ± SD. Results are analyzed by ANOVA and Tukey’s post hoc test. Upper letters (a, b, and c) represent significant differences. N.S: not significant. ***p < 0.001, **p < 0.01, *p < 0.05
Surface (Bottom) Winter Spring Summer Autumn F-value
Temperature (°C) 11.4 ± 1.3a (11.9 ± 0.8a) 14.3 ± 0.4b (13.4 ± 0.5a,b) 20.9 ± 1.5d (14.4 ± 2.2b) 19.0 ± 0.3c (17.9 ± 2.1c) 226.1*** (31.97*** ) Salinity (psu) 33.3 ± 1.2a (34.1 ± 0.3) 32.9 ± 1.6a (34.1 ± 0.4) 27.4 ± 5.3b (33.7 ± 1.8) 31.8 ± 2.0a (33.2 ± 0.6) 10.33*** (2.44N.S) DIN (µM) 29.1 ± 28.3a (17.3 ± 10.0a) 27.2 ± 26.7 a (10.6 ± 4.3a) 73.0 ± 54.3b (18.9 ± 20.4a) 26.4 ± 39.0a (45.2 ± 39.02b) 7.14*** (5.90**) DIP (µM) 1.3 ± 1.4 (0.8 ± 0.3) 0.9 ± 0.5 (0.8 ± 0.2) 1.9 ± 1.2 (1.0 ± 0.5) 0.9 ± 1.1 (0.7 ± 0.4) 2.43N.S (1.36 N.S) DSi (µM) 15.0 ± 5.3 (13.2 ± 2.6a) 18.3 ± 14.5 (12.9 ± 2.2a) 39.6 ± 26.8 (19.3 ± 6.8b) 18.8 ± 15.7 (16.0 ± 7.0a,b) 5.50** (4.32**) Chlorophyll a (µg L-1) 0.6 ± 0.1a (0.6 ± 0.1a,b) 1.2 ± 0.4 a (0.9 ± 0.5b,c) 1.2 ± 0.6a (0.5 ± 0.2a) 2.0 ± 1.1b (1.1 ± 0.3c) 8.85*** (8.86***) Phytoplankton abundance (103 cells L-1)
Total abundance 96.6 ± 74.6a 245.0 ± 117.4a,b 298.9 ± 126.6b 204.3 ± 169.6a,b 3.17* Diatom 49.7 ± 34.9a 77.4 ± 43.6a,b 126.6 ± 786.2a,b 150.7 ± 157.6b 3.21* Dinoflagellate 1.2 ± 1.3 3.0 ± 3.5 3.1 ± 2.5 3.9 ± 3.4 2.26 N.S Cryptophyte 45.2 ± 52.1a 136.0 ± 116.7b 46.1 ± 33.8a 47.0 ± 16.6a 5.91**
Euglenophyte 0.6 ± 0.9 28.5 ± 51.2 11.7 ± 21.1 2.2 ± 2.2 2.78N.S
28 이민지 · 김동선 · 김영옥 · 손문호 · 문창호 · 백승호
도달하였다. 크기분획된 Chl. a농도를 살펴보면, 동계에는 대부분의 정점에서 pico, nano, micro 크기의 생물군이 30% 전후로 비슷한 양상을 보였다. 춘계 내측 1-4정점에서는 Nano와 Pico 크기의 생물 이 전체의 75%이상을 차지하였고, 5-11정점에서는 상대적으로 Micro크기의 생물이 30~40% 전후로 상대적으로 높은 점유율을 보였다. 하계에는 총 엽록소 농도가 1 µg L-1전후로 낮게 나타났고, 그 중 Nano와 Pico 크기의 생물이 80% 전후로 높은 비율로 관찰 되었다. 추계는 총 엽록소 농도가 높게 나타난 정점에서는 Micro 크기의 비율이 높게 나타났고, 상대적으로 엽록소 농도가 낮은 정 점에서는 pico크기가 높게 나타나는 특징을 보였다.
식물플랑크톤 군집구조
울산만 표층에서 관찰된 식물플랑크톤의 총 개체수와 각 분류군별 점유율을 Fig. 4에 나타내었다. 동계 식물플랑크톤 개체수는 평균 1.0 × 105 cell L-1로 매우 낮았으며, 규조류(60.7%; Chaetoceros socialis, Skeletonema costatum-like species, Thalassiosira pacifica 우점)와 은편모조류(37.0%; Cryptomonas spp. 우점)의 점유율이 높게 관찰되었다. 춘계 모든 정점의 평균 식물플랑크톤 개체수는 2.4 × 105 cell L-1로 나타났으며, 춘계 식물플랑크톤 군집중 규조류 (36.0%; Chaetoceros compressus, Leptocylindrus danicus 우점)와 은편모조류(50.2%)가 높은 비율을 차지했다. 그 중에서 정점 1과 Fig. 2. Seasonal and horizontal distribution of nutrients at sampling stations of Ulsan Bay during the four seasons. (a) silicate, (b) nitrate + nitrite, (c) ammonium, and (d) phosphate. Black and white bars are values from surface and bottom, respectively.
정점 4에서는 유글레나조류Eutreptiella gymnastica가 각각 51%와 63%로 높은 비율을 차지했다. 하계 평균 식물플랑크톤 개체수는 2.9 × 105 cell L-1로 나타났다. 특이적으로 정점 1, 2, 3, 5에서 담수 기원 남조류가 최소 33.8%에서 최대 75.5%의 변동 범위를 보였다.
특히 정점 1, 2, 3에서는 담수남조 Oscillatoria sp.가 우점하였고, 정점 5에서는 Microcystis spp.의 개체수가 7.7 × 105 cell L-1로 극 우점 하였다. 정점 4에서는Eutreptiella gymnastica가 39.2%를 차 지하였으며, 이 외의 정점에서는 규조류(Nitzschia spp., S. costatum-
like species 우점)가 71.8%를 차지했다. 추계 평균 식물플랑크톤 개체수는 2.0 × 105 cell L-1였으며, 그룹별 군집조성을 살펴보면 정점 1-4에서는 은편모조류가 66.5%로 높은 비율을 차지하였고, 이 외의 정점에서는 규조류가 74.9%(Chaetoceros debilis, Chaetoceros curvisetus, S. costatum-like species)로 대부분을 차지하였다.
토 의
울산만에서 내측과 외측의 환경인자를 구분하여 평가하면, 수온은 동계(t-test: t = -5.833, p < 0.01)와 춘계(t-test: t = 4.247, p < 0.01), 하계(t-test: t = 2.876, p < 0.05)에 유의한 차이를 보였고, 추계에는 유의한 차이를 보이지 않았다(p > 0.05). 쿠로시오 지류인 대마난 류는 추계 강하게 확장되어 대한해협으로의 유입량또한 최대에 달 하는 것으로 보고되고 있고(Ohwada and Ogawa, 1966; Cho and Choe, 1988), 본 연구에서도 추계 내측과 외측 수온에 대한 유의 한 차이를 확인할 수 없었던 것은 추계 성층이 약화되는 계절적인 특성과 함께 따뜻한 대마 난류의 영향이 외측에서부터 내측까지 일정하게 영향을 미친 결과로 판단된다. 염분은 외측 평균 33.1 psu
± 1.32 psu로 거의 변하지 않는 특징을 보였으며, 내측의 경우 전 반적으로 외측에 비해 낮은 값을 보였다. 특히 하계 집중강우로 인 한 태화강으로부터의 담수유입 영향으로 내측 염분이 최저 13.5 psu (정점 1)로 낮은 값을 기록하였으며, 추계를 제외하고 내측 염분이 통계적으로 유의하게 낮다는 것을 알 수 있었다( p < 0.05)(Fig. 5). 결 과적으로 울산만에서 내측과 외측의 수온 및 염분농도의 환경인자가 명확한 차이를 보이지 않은 계절은 추계로 나타났고, 이는 강우량이 점차적으로 줄어드는 추계의 계절적 특징과 함께, 대마난류 기원의 고온 고염 외양수의 영향이 크게 작용된 것으로 판단된다. 아울러, 내측과 외측 염분의 차이는 하계에 두드러지게 나타났고, 이는 춘 계와 하계에 집중되는 강우량이 울산만 내측에 지대하게 영향을 미친 것으로 판단된다.
식물플랑크톤의 성장에 중요하게 영향을 미치는 영양염류에 대한 내측과 외측의 비교는 내측해역에서 전반적으로 높게 나타는 것을 확인할 수 있었다. 구체적으로 살펴보면, DIN(nitrite, nitrate, ammonium)은 동계와 추계에 정점간의 편차가 커서 유의한 차이를 보이지 않았지만(p > 0.05), 춘계(t-test: t = 2.978, p < 0.05)와 하계 (t-test: t = 4.898, p < 0.01)에는 내만이 유의하게 높은 값을 보였다 (Fig. 5). 아울러, DIP농도에 관해서도 정점간의 농도차이가 큰 동 계를 제외하고, 춘계(t-test: t = 4.303, p < 0.01), 하계(t-test: t = 4.699, p < 0.01), 추계(t-test: t = 3.098, p < 0.05)에 유의하게 내측정점에서 높 았다. 규조류의 성장에 중요한 DSi농도는 모든 계절에 유의하게 내측이 높은 값을 보였고, 특히, 하계에 큰 차이를 보이는 것으로 나타났다(t-test: t = 4.071, p < 0.01). 연안 내만의 영양염 공급원은 주로 육상으로부터 담수기원과 해저 퇴적물에 의한 용출이 있을 수 있다. 동계 내측과 외측에서 DIN 및 DIP 영양염의 유의한 차이를 보이지 않은 것은 담수기원의 영양염의 공급보다는 수층혼합으로 인한 저층 영양염류 공급의 영향이 크다고 할 수 있다. 반면, 성층이 형성되는 춘계와 하계 영양염 공급원은 담수기원 영양염의 공급이 크다고 판단된다. 이와 같은 근거는 염분과 영양염과의 관계에서 명확하게 제시 할 수 있었고, 염분과 DIN의 관계는 R2=0.79로, 높은 음의 상관성을 보였고, 아울러 DIP는 R2= 0.51로, DSi는 R2= 0.76 Fig. 3. Seasonal and horizontal changes in Chl-a concentration at
the surface at sampling stations of Ulasn Bay. Percentages of total Chl-a concentration for each size fraction (< 3 µm, > 3 µm and <
20 µm, > 20 µm) are shown.
30 이민지 · 김동선 · 김영옥 · 손문호 · 문창호 · 백승호
Fig. 4. Seasonal and horizontal changes in total phytoplankton abundance and their relative contributions in Ulsan Bay. (a) winter, (b) spring, (c) summer, and (d) autumn.
로, 통계적으로 유의한 음의 상관성을 확인 할 수 있었다(p < 0.01).
특히, 저염분화된 해역에 주요 영양염류의 공급이 현저하게 높은 내만의 부영양화현상은 하계에 두드러지게 나타났다(Fig. 6). 결과 적으로, 울산만에서 식물플랑크톤의 성장에 중요한 영양염의 공급 원은 동계에는 수층혼합에 의한 퇴적물 용출에 의하여 전 수층에 걸쳐 일정의 높은 영양염 농도를 유지하는데 기여하였을 가능성이 높고, 하계에는 태화강, 울산 및 온산시의 도시하천수 유입에 의한 영양염공급이 만 내측의 표층에 크게 영향을 미치는 것으로 판단된다.
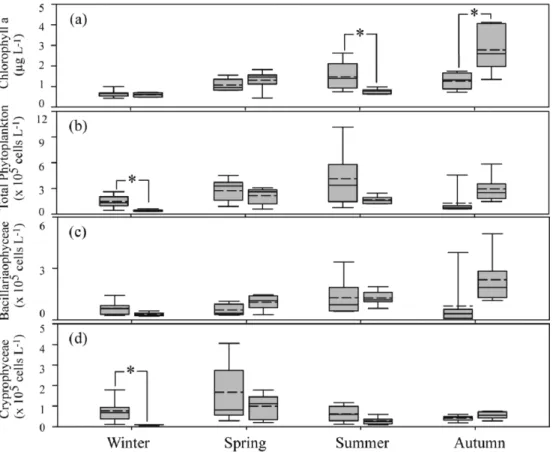
만 내측과 외측의 환경요인 차이에 따른 식물플랑크톤 현존량의 차이를 구체적으로 평가해 보았다. 먼저 식물플랑크톤에 기인한 총 Chl. a는 정점간 공간적인 차이를 보였고, 울산만 13개 정점의 평균 Chl. a의 계절적 변화양상은 추계> 춘계> 하계> 동계 순으로 높게 나타났다. 동계와 춘계에는 만 내측과 외측과의 Chl. a의 유의한 차이를 보이지 않았고, 상대적으로 낮은 값을 유지하였다. 하지만, 상기에서 언급된 바와 같이 영양염 농도가 만 내측해역에서 높게 관찰된 하계에는 외측해역보다 내측해역에서 유의하게 높은 값(t-test:
t = 3.402, p < 0.05)을 보였다. 반면, 추계에는 외측해역이 내측보다 유의하게 높은 값(t-test: t = -3.091, p < 0.05)이 관찰되었다(Fig. 7).
총 식물플랑크톤의 개체수는 동계, 춘계, 하계에는 전반적으로 내 측해역에서 높게 나타났지만, 유의한 차이를 보인 것은 동계에 국 한되었다. 반면, 추계에는 총 Chl. a 농도가 외측에서 높게 나타난 것과 같이 총 식물플랑크톤의 개체수 밀도 또한 외측에서 일정하게 높았지만, 통계적 유의한 차이를 보이진 않았다( p > 0.05). 울산만
에서 우점한 규조류의 만 내측과 외측의 비교 분석에서도 통계적 차이는 명확하지 않았으나, 전반적으로 수층혼합이 활발한 동계와 담수의 공급이 높게 나타나 하계에 내측해역에서 높게 나타났고, 춘계와 추계에는 외측해역에서 높게 나타나는 특성을 보였다. 아 울러 은편모조류의 비교에서도 전반적으로 동계(p < 0.01), 춘계(p >
0.05), 하계( p > 0.05)에 내측에서 높게 나타났고, 추계에는 내측과 외 측의 명확한 차이가 관찰되지 않았다. 결과적으로 하계에 내측에서 높은 영양염류가 나타났으나, Chl. a 농도가 1.5 µg L-1 전후로 높게 나타나지 않았다는 것은 하계에 공급된 영양염류가 조사 당일 기 준으로 식물플랑크톤의 증식에 직접적인 관여는 하지 않았을 가능 성이 높다. 따라서, 내측해역에서 식물플랑크톤에 의하여 영양염류의 소모되는 양상을 명확하게 파악하기 위해서는 강우 후 시계열의 자료의 축적이 중요할 것으로 판단된다.
울산만의 계절별 표층 환경인자와 식물플랑크톤 군집 자료를 바 탕으로 주성분분석(PCA)을 수행하였으며, 그 결과를 Table 2에 나 타내었다. 동계 제 1 주성분은 전체 기여율의 55.4%를 차지하였고, 식물플랑크톤 은편모조류의 성장에 영향을 미친 주요성분은 영양 염류 DIN, DIP, DSi로 나타났으며, 수온과 염분은 음의 (-)관계를 보였다. 아울러 제 2주성분(24.2%)을 포함한 누적 기여율은 79.9%로 나타나, 앞서 언급한 것과 같이 동계 식물플랑크톤의 성장은 수주 내 공급되는 영양염류의 대부분은 퇴적물 용출의 가능성이 높고, 이들 영양염류는 규조류와 은편모조류의 성장에 중요하게 작용하 였을 것으로 판단되었다. 춘계 주성분 분석에서는 제 1성분은 동
계와 유사하게 55.8%를 차지하였고, 그 중 유글레나류의 성장에 영향을 미친 요인으로는 수온과 함께 DIN, DIP, DSi로 판단되었다.
Palmer (1969)와 Kim and Boo (2000)의 보고에 의하면, 유글레나는 영양염류에 민감하게 반응하는 종이다. 주로 춘계 수온증가와 함 께 공급되는 영양염류에 반응하여 대증식을 일으키는 종으로 잘 알려져 있으며, 특히 본 종은 부영양화 또는 오염된 해역의 지표 및 지시종으로도 활용되고 있다. 결과적으로 수층혼합이 활발한 동 계에는 울산만에서 은편모조류가 우점할 수 있었고, 성층이 형성 되는 춘계로 갈수록 내측을 중심으로 유글레나조류가 영양염류에 민감하게 반응하여 일정의 높은 개체수를 유지할 수 있는 환경이 라는 것을 시사할 수 있다. 하계의 주성분 분석에서는 제 1주성분과 제 2성분의 누적 기여율은 77.1%로 나타났고, 그 중 제1성분의 기 여율은 57.3%로 나타났다. 제 1성분의 주요요인은 수온, DIN, DIP, DSi의 주요영양염류가 식물플랑크톤의 총 엽록소 Chl. a의 생성에
중요한 요인으로 판단되었다. 하지만, 특정 그룹의 주요성분의 분 석은 명확하게 파악할 수 없었다. 제2성분을 누적하여 평가하면, 특이적으로 남조류와의 상관성을 파악할 수 있었다. 즉 이는 담수 기원의 남조류(주로 Oscillatoria sp., Microcystis sp.)가 강우의 유 입과 함께 연안 내측해역에 영향을 미친 것으로 파악할 수 있다.
실제 이들 담수종은 높은 염분농도에 적응할 수 없어 시간의 경과와 더불어 사멸할 것으로 판단되어, 울산만의 1차 생산력에는 크게 기 여하지 못할 것으로 판단된다. Kim et al. (2011)의 진해만 보고에 의하면, 하계 DIN, DIP, DSi 영양염류와 Chl. a 양의 상관관계를, 염분과 높은 음의 상관성을 보여 담수 영향이 진해만의 식물플랑 크톤의 증식에 영향을 미치는 것으로 파악되었다. 울산만에서도, 하계 담수에서 공급되는 영양염류가 식물플랑크톤 성장에 중요한 작용될 것으로 파악된다. 하지만, 앞선 단락에서도 언급 하였는 것 과 같이 영양염류의 공급 된 후 곧바로 식물플랑크톤의 성장은 일 Fig. 5. Box plot of abiotic factors in the surface waters of Ulasn Bay during the four seasons. The median and average are presented by solid lines and dotted line, respectively (Significant at **p < 0.01; *p < 0.05).
32 이민지 · 김동선 · 김영옥 · 손문호 · 문창호 · 백승호
어나지 않고, 일정의 시간차(time lag) 두고 반응하기 때문에 주의 깊게 해석할 필요성은 있다. 추계의 주성분분석에서는 각 주요 우 점 분류군의 식물플랑크톤 현존량과 음(-)의 상관성이 관찰되었고, 결과적으로 추계의 주성분은 다른 3계절과 다른 양상에 의하여 주 요성분이 작용되었을 가능성이 높다. 추계의 제1성분과 제2성분의 누적 기여율은 73.5%로 다른 계절과 유사하게 나타났다. 앞서 언 급한 내측과 외측 해역의 차이에서도 추계에는 수온과 염분의 차 이가 명확하지 않아, 외부 물리적인 환경에 의하여 외측에서 내측 으로 영향을 미쳤을 것으로 사료되었고, 이와 같은 물리적인 작용이 주성분분석에서도 반영된 것으로 나타났다. 추후 시계열 자료를 근 거로 외부기인 미세조류의 번성에 의한 집적현상등과 같은 요인에 관해서도 심도있는 토의가 요구된다.
울산만에서 크기 분획된 Chl. a농도를 구체적으로 살펴보면, 동계 Micro 크기는 27.5%로, Nano 크기는 35.6%로, Pico-size가 36.9%로 나타나, Nano와 Pico 크기의 생물군이 전체70%이상을 차지하였 다. 이는 Kwon and Kang (2013)의 울산만 동계의 분포양상과 유 사하게 나타났다. 춘계의 크기 분획된 Chl. a농도는 Micro 크기가 30.7%, Nano 크기는 28.2%, Pico 크기는 41.1%로 나타났다. 내측 정점에서는 Nano와 Pico 크기의 생물이 전체의 75%이상으로 높게 나타났고, 외측 정점에서는 Micro 크기의 생물이 30-40%의 점유 율을 보였다. 특히 Nano와 Pico 크기 식물플랑크톤의 Chl. a가 전 체 Chl. a의 80.8%를 차지하였다. 일반적으로 Micro 크기 식물플 랑크톤은 영양염이 풍부한 상태에서 높게 분포하고, Pico 및 Nano 크기의 식물플랑크톤은 영양염 농도가 낮은 환경조건에서 효과적 으로 영양염류을 uptake하여 빠르게 성장한다고 알려져있다(Iriarte
Fig. 7. Box plot of biotic factors in Ulsan Bay during the four seasons. The median and average are presented by solid lines and dotted line, respectively (Significant at **p < 0.01; *p < 0.05).
Fig. 6. Relationships between salinity and nutrients (DIN, DIP, DSi).
and Purdie, 1994). 하지만 본 결과의 식물플랑크톤의 군집 조성에 서, 은편모조류가 외측과 내측에서 높은 밀도로 우점하여 높은 영 양염 조건에도 불구하고 춘계 내만에서 Pico 및 Nano 크기의 Chl. a의 농도가 나타난 것으로 판단된다. 하계의 Chl. a농도는 대부분의 정 점에서 상대적으로 낮게 나타났고, 크기 분획된 평균 Chl. a농도는 Pico 크기가 17.4%로, Nano 크기가 56.4%로, Micro 크기가 26.3%로, Nano 크기로 분획된 생물량이 가장 높게 나타났다. 특히 내만 정 점에서는 영양염 농도가 높게 나타났음에도 불구하고, Micro 크기 가 증식할 수 없었던 이유는 조사 3일전 심한 강우의 영향으로 일 시적으로 공급된 영양염이 조사당일 식물플랑크톤의 성장에는 그 다지 영향을 미치지 못한 결과로 사료된다. 결과적으로 하계에는 강우가 관측되기 전까지 울산만해역에 낮은 영양염이 유지되어, Pico와 Nano크기 분획이 높게 유지되었을 것으로 사료된다. 반면, 추계에는 상대적으로 다른 3계절과 비교하여 높은 Chl. a값이 관 찰되었다. 특히 Chl. a가 4 µg L-1 이상의 값을 보였던 정점7과 12 에서는 Micro 크기 분획된 Chl. a농도가 78%와 64%로 높게 나타 났다. 이들 두 정점을 제외하면, 춘계의 크기 분획된 결과와 유사 하게 나타났다. 결과적으로 2014년 울산만의 총 Chl. a농도는 Kwon and Kang (2013)이 보고한 결과보다 상대적으로 낮게 나타나, 해 역간의 공간적인 분포양상은 명확한 특징을 파악할 수 없었지만, 전반적으로 Pico와 Nano 크기로 분획된 Chl. a의 농도가 전반적으로 높게 나타나는 특색으로 보였다. 이는 울산만해역이 외양의 낮은 영양염 농도의 영향을 강하게 받고 있다는 것을 시사할 수 있다.
울산만의 계절적 식물플랑크톤의 군집구조를 살펴보면, 외측해 역을 중심으로 규조류가 우점하였고, 내측해역에서는 전반적으로 은편모조류가 우점하였다. Lee et al. (2005)의 아산만의 보고에 의 하면, 본 조사와 유사하게 내측해역에서는 은편모조류가 우점 출 현하였고 외측해역에서는 규조류가 높은 점유율을 보고하였다. 특 이적으로 울산만에서는 춘계(정점1~4)와 하계(정점4)에 Eutreptiella gymnastica가 내측 정점을 중심으로 높게 나타났다. 유글레나조류 E. gymnastica는 영양염류의 부하량이 높은 수계에서 대발생 하는 것으로 알려져 있다(Kant, 1989; Kim et al., 1998). Kim and Boo (2001)의 연구에 의하면, E. gymnastica와 같은 유글레나조류는 영
양염류의 상관 관계가 있고, 동계 대발생한 유글레나류는 인산염과 양의 상관를 보였고, 하계에 대발생한 유글레나류는 질산염과 양의 상관성을 보였다. 본 연구에서도 춘계 정점 1과 4의 DIN 농도가 83.4 µM와 80.2 µM로 다른 정점과 비교하여 매우 높은 값을 보 였으며, 하계 정점4에서도 DIN 농도가 106.1 µM로 극히 높은 값을 보였다. 결과적으로 E. gymnastica의 성장은 DIN농도가 높은 해 역에서 특이적으로 우점하였고, 이는 담수기원 의 높은 영양염이 갑작스럽게 공급될 경우, 기회적으로 빠르게 성장하여 대발생 할 것으로 판단된다. 아울러, 하계 내측정점 1, 2, 3, 5에서는 남조류 가 각각 44.3%, 33.9%, 33.8%, 75.5%로 높은 비율을 차지하였다.
특히 정점 1, 2, 3에서는 Oscillatoria sp., 정점 5에서는 Microcystis sp.가 극우점 하였다. PCA분석의 결과분석에서도 언급한 것과 같 이, 두 종은 담수에 서식하는 남조류로 태화강 상류에서 여름철 높은 수온으로 우점하여 대발생한 후 강우에 의하여 내측정점으로 유입 된 결과로 판단된다. 특히 하계 조사 전 3일간 누적 강수량은 98.6 mm로 높았고, 내측정점에서 최저 표층염분이 13.5 psu로 기 수역의 특성을 보였다(2014년 8월 기상청 일강수량 자료). 결과적 으로 하계 내측정점에서 담수종의 우점은 태화강의 영향을 크게 받아 나타난 일시적인 현상으로 파악되었다. 따라서 담수기원의 미 세조류의 개체수 밀도는 상대적으로 높게 나타났으나, 해수의 높 은 염분에 적응할 수 없기 때문에 엽록소 활성을 잃어 Chl. a농도 가 낮게 나타난 것으로 파악되었다. 추계와 동계의 식물플랑크톤 군집조성은 정점간의 점유비율의 차이는 있었으나, 전반적으로 규 조류와 더불어 은편모조류가 대부분을 차지하였다. 그 중 규조류의 우점종은 중심목의 Chaetoceros 속 Chaetoceros debilis, Chaetoceros curvisetus, Chaetoceros socialis로 나타났고, 추계 현존량에 가장 크게 기여한 종은 Chaetoceros debilis, Chaetoceros curvisetus로 나타났다. 이들 종은 수온이 낮은 추계에서 동계에 걸쳐 한국 연 안에서 우점종으로 출현하는 종으로 보고되어 왔다(Oh et al., 2008;
Park et al., 2009). 결과적으로 울산만에 추계에 우점한 Chaetoceros 속은 성층이 붕괴되고 난 후 공급되는 영양염류와 함께 수온의 하 강이 영향을 미친것으로 판단된다. 규조류 다음으로 계절적으로 높은 밀도로 출현한 종은 은편모조류로 나타났고, 그들은 동계와 추계에 Table 2. Loadings of parameters for the first two principal components in the surface waters of Ulsan Bay, Korea (Loadings > 0.6 are bolded).
Parameters Winter Spring Summer Autumn
PC1 PC2 PC1 PC2 PC1 PC2 PC1 PC2
Temperature -0.84 0.14 0.77 0.31 0.80 -0.04 -0.70 -0.20
Salinity -0.92 0.27 -0.93 -0.02 -0.94 0.09 -0.86 -0.47
DIN 0.91 -0.30 0.95 0.24 0.98 0.08 0.90 0.39
DIP 0.80 -0.31 0.96 -0.10 0.96 0.08 0.96 0.15
DSi 0.94 -0.30 0.83 0.33 0.99 -0.04 0.91 0.28
Chl. a 0.55 0.72 -0.52 0.67 0.92 -0.17 -0.61 0.55
Diatom 0.64 -0.02 -0.48 0.85 0.46 -0.51 -0.79 0.48
Dinoflagellate 0.43 0.75 -0.68 0.38 -0.52 0.70 -0.60 0.55
Cryptophyte 0.85 0.43 0.10 -0.12 0.58 0.65 -0.32 0.82
Euglenophyte 0.11 0.88 0.79 0.45 0.10 0.74 0.31 0.00
Cyanophyte 0.00 0.00 0.00 0.00 0.49 0.63 0.00 0.00
Eigenvalue 5.54 2.42 5.58 1.81 6.30 2.18 5.33 2.02
Variability (%) 55.39 24.21 55.75 18.05 57.31 19.80 53.31 20.15 Cumulative % 55.39 79.60 55.75 73.81 57.31 77.11 53.31 73.47
34 이민지 · 김동선 · 김영옥 · 손문호 · 문창호 · 백승호
전체 식물플랑크톤 군집 중 각각 59.6%와 57.4%로 높은 비율을 보였다. 특히 태화강의 영향을 강하게 받는 내측해역(정점 1~4)에서 은편모조류의 높은 점유율(>77%)을 보였다. Marshall and Lacouture (1986)의 의하면, Chesapeake Bay에서 은편모조류가 하 계와 추계에 높은 비율을 차지하였고, 동계에는 상대적으로 낮은 개체수를 보고하였다. Baek et al. (2015)의 광양만의 연구에서는 추계 Chl. a농도가 낮은 시기 은편모조류가 전체 군집의 >75%을 차지하였고, 동계에는 낮은 밀도를 유지하였다. Sommer (1985)와 Klaveness (1989)는 영양염에 대한 경쟁대상 생물이 존재하지 않는 수역에서 기회적으로 빠른 성장을 보이는 은편모조류가 우점할 수 있다고 보고하였다. 아울러, 은편모조류는 탁도가 높고 낮은 광조 건에서도 비교적 잘 적응하여 우점하는 종으로 알려져 있다(Barone and Naselli-Flores, 2003). Bae et al. (2014)은 광양만에서 2010~2012 년 추계에 은편모조류의 높은 개체수 밀도를 관찰하였고, 이는 추 계 성층파괴와 더불어 수층혼합으로 퇴적층의 미세입자가 재부유 되어 수계 내 투명도가 현저히 떨어져, 다른 경쟁종이 증식하지 않는 시기에 은편모조류가 기회적으로 우점한다고 보고하였다. 본 연구 에서도 선행된 연구결과와 유사하게 추계의 투명도가 현저하게 떨 어져, 다른 식물플랑크톤 군집이 우점하지 않는 시기에 기회적으로 우점하는 특색을 보였고, 이와 같은 경향은 수심이 낮은 태화강 하 구의 내만 정점에서 두드러지게 나타났다. 결과적으로 은편모조류는 추계와 동계의 다른 식물플랑크톤의 성장이 상대적으로 둔화되는 계절에 기회적으로 우점할 가능성이 높은 종으로 파악되었다. 요 약하면, 울산만은 내측과 외측의 계절적 환경요인의 차이가 뚜렷 하게 나타났고, 이는 식물플랑크톤의 군집조성의 공간적 분포 양 상에도 영향을 미친 것으로 판단되었다. 특히 전 계절에 걸쳐 외 측해역에는 규조류가 내측해역에는 은편모조류가 전반적으로 우점 하였으며, 춘계와 하계 내측정점에서 담수의 영향을 받아 유글레 나류와 남조류가 일시적으로 아주 높은 개체수를 보였다. 추계와 동계에는 규조류와 은편모조류가 대부분을 차지하였고, 그 중 중 심목 규조류 Chaetoceros 속이 추계 생물 현존량에 기여한 것으로 나타났다. 동계 엽록소가 낮은 시기에는 은편모조류가 상대적으로 높은 개체수를 보였는데, 이는 다른 종과의 경쟁이 상대적으로 적을 때 기회적으로 빠르게 성장하는 은편모조류의 생태전략으로 판단 된다.
사 사
본 연구는 한국해양과학기술원 주요사업(PE9931A) 및 해양수 산부 “독도의 지속가능한 이용연구(PG48910)”의 지원에 의하여 수행되었다.
참고문헌(References)
Anderson, D.M., P.M. Glibert and J.M. Burkholder, 2002. Harmful algal blooms and eutrophication: nutrient sources, composition, and consequences. Estuaries, 25: 704−726.
Bae, S.W., D. Kim, Y.O. Kim, C.H. Moon and S.H. Baek, 2014. The influences of additional nutrients on phytoplankton growth and horizontal phytoplankton community distribution during the autumn season in Gwangyang Bay, Korea. Korean J. Environ. Biol., 32:
35−48.
Baek, S.H., D. Kim, M. Son, S.M. Yun and Y.O. Kim, 2015. Sea- sonal distribution of phytoplankton assemblages and nutrient- enriched bioassays as indicators of nutrient limitation of phyto- plankton growth in Gwangyang Bay, Korea. Estuar. Coast. Shelf.
Sci., 163: 265−278.
Barone, R. and L. Naselli-Flores, 2003. Distribution and seasonal dynamics of cryptomonads in Sicilian water bodies. Hydrobio- logia, 502: 325−329.
Cho, K.-D. and Y.-K. Choe, 1988. Seasonal variation of the water type in the Tsushima Current. Korea. Fish. Aquatic. Sci., 21: 331−340.
Cloern, J.E., 2001. Our evolving conceptual model of the coastal eutrophication problem. Marine Ecology Progress Series, 210:
223−253.
Cushing, D.H., 1989. A difference in structure between ecosystems in strongly stratified waters and in those that are only weakly stratified.
J. Plankton Res., 11: 1−13.
Dortch, Q. and T.E. Whitledge, 1992. Does nitrogen or silicon limit phytoplankton production in the Mississippi River plume and nearby regions? Cont. Shelf Res., 12: 1293−1309.
Fisher, T., 1992. Nutrient limitation of phytoplankton in Chesapeake Bay. Mar. Ecol. Prog. Ser., 82: 51−63.
Goldman, J.C., J.J. McCarthy and D.G. Peavey, 1979. Growth rate influence on the chemical composition of phytoplankton in oce- anic waters. Nature, 279: 210−215.
Guinder, V.A., C.A. Popovich, J.C. Molinero and J. Marcovecchio, 2013. Phytoplankton summer bloom dynamics in the Bahía Blanca Estuary in relation to changing environmental conditions. Cont.
Shelf Res., 52: 150−158.
Iriarte, A. and D.A. Purdie, 1994. Size Distribution of chlorophyll-a biomass and primary production in a temperate estuary (South- ampton water) - the contribution of photosynthetic picoplankton.
Mar. Ecol. Prog. Ser., 115: 283−297.
Joo, H.M., J.H. Lee and S.W. Jung 2011. Correlations between cell abundance, bio-volume and chlorophyll a concentration of phy- toplankton communities in coastal waters of Incheon, Tongyeong and Ulsan of Korea. Korean J. Environ. Biol., 29: 312−320.
Kattner, G., 1999. Storage of dissolved inorganic nutrients in seawa- ter: poisoning with mercuric chloride. Mar. Chem., 67: 61−66.
Kim, D., H.-W. Choi, K.H. Kim, J H. Jeong, S.H. Baek and Y.-O. Kim, 2011. Statistical analysis on the quality of surface water in Jinhae Bay during winter and spring. Ocean Polar Res., 33: 291−301.
Kim, J.T. and S.M. Boo, 2001. Morphological variation and density of Euglena viridis (Euglenophyceae) related to environmental factors in the urban drainages. Korean J. Limnol., 34: 185−191.
Kim, J.T., S.M. Boo and B. Zakrys, 1998. Floristic and taxonomic accounts of the genus Euglena (Euglenophyceae) from Korean fresh waters. Algae, 13: 173−197.
Klaveness, D., 1989. Biology and ecology of the Cryptophyceae: status and challenges. Biol. Oceanogr., 6: 257−270.
Koh, C.H., J.S. Khim, D.L. Villeneuve, K. Kannan and J.P. Giesy, 2002. Analysis of trace organic contaminants in sediment, pore water, and water samples from Onsan Bay, Korea: instrumental analysis and in vitro gene expression assay. Environ. Toxicol. Chem., 21: 1796−1803.
Kwon, O.Y. and J.-H. Kang, 2013. Seasonal variation of physico- chemical factors and size-fractionated phytoplankton biomass at Ulsan seaport of East Sea in Korea. J. Korea. Acad. Industr. Coop.
Soc., 14: 6008−6014.
Lee, S., Y. Sin, S. Yang and C. Park, 2005. Seasonal characteristics of phytoplankton distribution in Asan Bay. Ocean Polar Res., 27:
149−159.
Marshall, H.G. and R. Lacouture, 1986. Seasonal patterns of growth and composition of phytoplankton in the lower Chesapeake Bay and vicinity. Cont. Shelf Res., 23: 115−130.
Oh, S.J., J.S. Lee, J.S. Park, I.H. Noh and Y.H. Yoon, 2008. Envi- ronmental factor on the succession of phytoplankton community in Jinju Bay, Korea. Korean Soc. Mar. Environ Eng., 11: 98−104.
Ohwada, M. and F. Ogawa, 1966. Plankton in the Japan Sea. Oceanogr.
Mag., 18: 39−42.
Palmer, C.M., 1969. A composite rating of algae tolerating organic pollution2. J. Phycol., 5: 78−82.
Park, J.S., Y.H. Yoon and S.J. Oh, 2009. Variational characteristics of phytoplankton community in the mouth parts of Gamak Bay, south- ern Korea. Korean J. Environ. Biol., 27: 205−215.
Park, S.-E., S.-J. Hong, W.-C. Lee, R.-H. Jung, Y.-S. Cho, H.-C. Kim and D.-M. Kim, 2010. Summer water quality management by eco- logical modelling in Ulsan Bay. Korean Soc. Mar. Environ. Saf., 16: 1−9.
Parsons, T.R., Y. Maita and C.M. Lalli, 1984. A Manual of Biolog- ical and Chemical Methods for Seawater Analysis. Publ. Perga- mon Press, Oxford: pp. 184.
Riley, G.A., 1942. The relationship of vertical turbulence and spring diatom flowerings. J. mar. Res., 5: 67−87.
Sommer, U., 1985. Comparison between steady-state and non-steady state competition - experiments with natural phytoplankton. Lim- nol. Oceanogr., 30: 335−346.
Thompson, P.A., P.I. Bonham and K.M. Swadling, 2008. Phytoplankton blooms in the Huon Estuary, Tasmania: top-down or bottom-up control?. J. Plankton Res., 30: 735−753.
2015년 12월 14일 원고접수 2016년 2월 4일 수정본 접수 2016년 2월 14일 수정본 채택 담당편집위원: 박종규