Article
여름철 및 겨울철 수괴에 따른 대한해협의 동물플랑크톤 분포 양상
장민철·백승호·장풍국·이우진·신경순 * 한국해양연구원 남해분원
(656-830) 경상남도 거제시 장목면 장목1길 41
Patterns of Zooplankton Distribution as Related to Water Masses in the Korea Strait during Winter and Summer
Min-Chul Jang, Seung Ho Baek, Pung-Guk Jang, Woo-Jin Lee, and Kyoungsoon Shin * South Sea Branch, KORDI
Geoje 656-830, Korea
Abstract : We investigated the distribution and species composition of zooplankton in relation to hydrographical characteristics in the Korea Strait during the winter (February) and summer (July) of 2009.
Satellite images of sea surface temperatures and in situ CTD data showed that the southeastern water zone (St3-5) off Jeju Island was strongly influenced by the Tsushima Current during both the winter and summer, whereas the Changjiang Diluted Water, characterized as water with relatively low salinity, was evident in the coastal waters of Jeju Island during the summer. During winter, zooplankton abundance was significantly higher than in the summer, with dominance by copepods, ostracods, siphonophorans, appendicularians, and nauplii. In both seasons, copepods represented >70% of the total zooplankton population. Calanus sinicus, a large calanoid copepod, was dominant in near the coast, and that may be associated with the intrusion of low salinity water (i.e., the Changjiang Diluted Water) along the coast. The abundance of P. parvus s.l. and A. omorii, known as neritic copepods, was mainly associated with the Korea Southern Coastal Water. Foraminiferans, Ostracods, O. plumifera, and P. aculeatus were concentrated in the southeastern water off Jeju Island during both seasons; showing their association with the Tsushima Current, which is characterized warm, high salinity water. Our results suggest that the distribution, abundance, and species composition of zooplankton are highly influenced by different water masses in the Korea Strait.
Key words : zooplankton, Korea Strait, water mass, Tsushima Current, Changjiang Diluted Water, Korea Southern Coastal Water
1. 서 론
동물플랑크톤은 기초생산자인 식물플랑크톤을 섭식하 여 보다 상위영양 소비자에 속하는 어류, 해파리 등에 에 너지를 전달함으로써 해양생태계를 유지하는 중요한 중간 고리 역할을 담당하고 있다. 운동 능력이 제한적인 동물플
랑크톤의 분포 양상, 개체수 변동, 종 천이 등은 계절에 따 른 수괴의 변화에 매우 민감하게 반응한다(Uye et al.
2000; Tseng et al. 2008). 이러한 특성으로 동물플랑크톤 의 여러 군집(특히, 요각류)은 수괴 및 해류의 변화와 분 포 상태를 나타내는 지시자로 이용되어 왔다(Grant 1991;
Hwang and Wong 2005; Hwang et al. 2006; Tseng et al.
2008). 전 지구적인 기후 변화의 상황에서, 여러 해양생태 계에서 수온, 염분과 같은 물리적인 요인들의 급격한 변화
*Corresponding author. E-mail : [email protected]
가 예상되며(Chen and Folt 2002), 이러한 변화는 고유한 수괴에도 변화의 가능성을 암시한다. 이러한 맥락에서 수 괴와 관련된 동물플랑크톤의 계절적인 변동 특성에 대한 연구는 현재 해양생태계의 상태에 대한 심도 깊은 이해를 제공할 수 있으며, 향후 전 지구적 또는 특정 해역에 대한 기후변화 및 해양환경 변화와 관련된 예측에 도움이 되는 자료로 이용될 수 있다.
우리나라 남해와 일본열도 사이에 위치한 대한해협 (Korea Strait) 은 동중국해와 동해를 연결하는 좁은 대륙붕 해역으로 중앙에 위치한 수심 220 m의 해곡을 제외하면 수심 130 m 미만의 평탄한 지형으로 이루어져 있다. 대한 해협은 북태평양의 서부를 따라 북상하는 쿠로시오 해류 (Kuroshio Current) 로부터 분지되어 동중국해에서 동해로 흐르는 대마 난류(Tsushima Warm Current)에 의해 크게 지배된다. 대마 난류는 중국 연안수와 동중국해에서 혼합 되어 제주도 인근 해역을 거쳐 대한해협을 통과한 후 동 한 난류를 형성하며 동해로 유입됨으로써 우리나라 남, 동 해에 영양염과 다양한 물질을 수송함과 더불어 해수 순환 을 조절하는 중요한 역할을 한다(Moriyasu 1972; Yanagi 2002; Onitsuka et al. 2007; Morimoto et al. 2009). 대한 해협에는 대마 난류 이외에도 남해 연안수(Korea Southern Coastal Water), 장강 희석수(Changjiang Diluted Water) 등의 다양한 수괴가 존재하며 계절에 따라 그 확장범위를 달리하며 남해 연안까지 영향을 끼쳐 계절변화가 크다(승 1992; 조와 김 1994). 특히, 겨울철 남해 연안수는 북서계 절풍과 수표면 열 이동 등의 영향에 의해 대한해협 권역 의 외해역으로 확장되어 대마 난류와의 사이에 강한 연안 전선을 형성한다(나 등 1990; 양 1994). 여름철은 제주도 북서쪽을 경유한 장강 희석수로 대표되는 중국 대륙연안 수의 유입에 의해 남해 및 대한해협 권역에는 저염화 현 상과 함께 염전선이 발달한다(조와 김 1994; 조 등 1995).
각 수괴 사이에 형성된 전선의 생성, 이동, 소멸은 수괴의 시, 공간적인 변동을 유도하며, 결국 동물플랑크톤의 종조 성, 개체수, 다양도 등에 영향을 미침으로써 분포 양상과 군집 구조를 변화시키는 주요한 요인으로 작용한다(Smith and Vidal 1986; 박 등 1991; 박 등 1998).
그 동안 대한해협 인근 해역에서 수행된 동물플랑크톤 에 대한 연구는 요각류의 계절적 수직분포 특성(이 등 1999), 요각류 및 모악류의 분포와 수괴 특성(박 1973a;
박 등 1990; Kang and Hong 1995), 동물플랑크톤의 양적 분포(김 1972; 박 1973b) 등을 다룬 연구가 진행되었다.
그러나 선행된 연구는 대부분 요각류와 모악류의 분포와 물리적 환경특성을 이해하는데 중점을 두거나 자원량 관 점의 동물플랑크톤의 양적 분포 조사에 치우쳐 있어 대한 해협의 전체 동물플랑크톤을 대상으로 수괴 특성과 군집 구조를 연구한 사례는 매우 미흡하다.
본 연구의 목적은 대한해협 및 남해에서 2월과 7월의 동물플랑크톤 군집 구조와 공간적 분포 특성을 이해하는 것이다. 이를 위해 해역의 수리학적인 수괴 특성을 파악하 고, 이를 동물플랑크톤의 집괴분석 결과와 비교·분석하였 다. 또한, 정준대응분석(Canonical Correspondence Analysis:
CCA)을 통하여 수괴의 변화에 민감하게 반응하는 calanoid 요각류의 연안종(neritic species)과 외양성 난류 종(oceanic warm-water species)의 분포 양상과 환경요인 들과의 상관성을 이해하였다.
2. 재료 및 방법
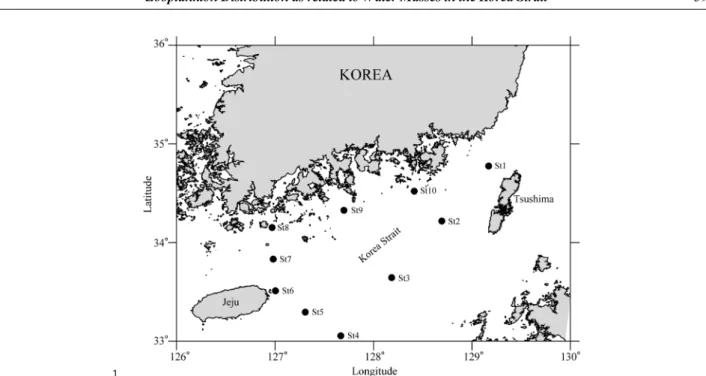
대한해협을 포함한 남해권역에 대한 동물플랑크톤의 조 사는 겨울철(2009년 2월 10-12일)과 여름철(2009년 7월 16-18 일)에 해양조사선 이어도호(R/V Eardo)를 이용하여 부산 외양의 대한해협 상단에서 배타적 경제수역 (Exclusive Economic Zone; EEZ)의 경계선을 따라 제주 도 남동 해역으로 이어지는 라인(St. 1-4), 제주도 동쪽의 남단에서 북단으로 연결되는 라인(St. 5-7), 보길도 인근해 역에서 남해 연안을 따라 거제도 외해역까지 이어지는 라 인(St. 8-10) 등 총 10개 정점에서 수행하였다(Fig. 1). 각 조사정점에 대한 위치, 조사일 및 시간, 수심 정보는 Table 1 에 나타냈다.
환경요인 및 동물플랑크톤
대한해협 권역에서 조사기간이 포함된 표층수온(SST) 의 주간 평균 자료는 국립수산과학원(NFRDA)의 위성해 양정보시스템(http://portal.nfrdi.re.kr/sois/)에서 제공하는 자료를 사용하였으며, 이 자료는 NOAA의 AVHRR (Advanced Very High Resolution Radiometer) 위성자료를 바탕으로 분석되었다. 각 조사 정점의 수온과 염분의 수 직 프로파일은 SBE 911plus CTD(Sea-Bird Electronics) 를 이용하여 측정하였고, 수심과 좌표(위도, 경도)는 조사 선에 장착된 장비로부터 수집하였다. 엽록소-a 농도를 측 정하기 위한 해수 채수는 로젯멀티 샘플러(Rosette Multi- sampler)에 부착된 Niskin 채수기를 이용하였고, 각 정점 의 수심 정도에 따라 0, 5, 10, 20, 30, 50, 75, 100 m와 Bottom 을 기준으로 채수하였다. 각 수심에서 채수된 해수 의 1-3 l를 선상에서 GF/F 필터지(47 mm Whatman glass fiber filters) 로 여과하여, 15 ml tube에 넣어 냉동 보관하 였고, 실험실에서 90% 아세톤 10 ml를 주입하여 24시간 동안 냉암소에서 색소를 추출한 후에 3000 rpm으로 5분 간 원심 분리시켜 상등액을 취하여 형광 광도계(Turner designs 10-AU)를 사용하여 측정하였다(Parsons et al.
1984).
동물플랑크톤 채집은 표준형 네트(망구 60 cm, 망목
200 µm)를 이용하여 바닥 5 m 위에서 표층까지 주야 구 분 없이 0.5-1 m s
−1의 속도로 수직 예인하였다. 채집된 시료는 선상에서 즉시 중성포르말린으로 최종 농도가 5%
가 되도록 고정하여 실험실로 운반하였다. 정량분석을 위 해 네트 입구에 유량계(Hydro-Bios model 438115)를 부 착하여 여과된 해수량을 측정하였다. 실험실에서 우점종 의 개체수가 200개체 이상 계수될 정도의 시료를 취하여 Bogorove 계수판에서 해부현미경(Zeiss Stemi SV11)과 광학현미경(Zeiss Axioskop) 하에서 동정 및 계수하여 단 위체적 당 개체수(indiv. m
−3) 로 환산하였다.
자료분석
동물플랑크톤 군집의 생태환경을 비교 분석하기 위한 군집지수로 Shannon and Weaver (1963)의 종 다양성 지 수 (H’)를 구하였다. 동물플랑크톤의 종조성과 출현 개체 수에 대한 조사월간의 유의한 차이를 검정하기 위하여 통 계 패키지 SPSS(version12.0)를 이용하여 독립표본 t 검정 (two independent samples t-test) 를 수행하였다. 동물플랑 크톤 출현양상의 유사도를 파악하기 위해 Bary-Curtis의 유사도지수를 근거로 비가중 산술평균(UPGMA)에 의한 dendrogram을 작성한 후, 계층적 군집방법(Hierarchical Fig. 1. Map of the study area showing locations of sampling station in the Korea Strait during February and July,
2009
Table 1. Summary of the sampling stations and water samples used for testing hydrographic conditions during February and July, 2009
Station Latitude (N)
Longitude (E)
Date Time Bottom
depth (m)
Sampling depth (m)
CTD depth Feb. 2009 Jul. 2009 Feb. 2009 Jul. 2009 (m)
1 34
o47.32' 129
o05.97' 10 16 15:00 17:15 105 0-100 103
2 34
o16.44' 128
o40.45' 10 16 22:20 23:30 94 0-90 92
3 33
o44.84' 128
o15.65' 11 17 2:00 6:30 127 0-122 125
4 33
o14.58' 127
o49.44' 11 17 7:15 11:50 118 0-103 116
5 33
o22.30' 127
o24.00' 11 17 11:50 15:10 130 0-125 126
6 33
o30.00' 127
o00.00' 11 17 15:35 17:30 121 0-116 119
7 33
o49.00' 127
o00.00' 11 17 19:30 21:30 92 0-87 88
8 34
o08.00' 127
o00.00' 11 18 22:50 1:00 42 0-37 38
9 34
o19.00' 127
o45.00' 12 18 3:20 5:30 39 0-34 34
10 34
o28.17' 128
o20.44' 12 18 7:00 9:05 56 0-51 53
clustering) 으로 집괴분석(cluster analysis)을 하였다. 집괴 분석은 Clarke and Gorley (2006)의 매뉴얼을 참고하여 PRIMER(version 6.1.6) 프로그램을 이용하여 수행하였 다. 정준대응분석(Canonical Correspondence Analysis:
CCA) 은 CANOCO(CANOnical Community Ordination) 프로그램(version 4.5; Ter Braak and Šmilauer 2002)을 이 용하여 환경요인(수온, 염분, 수심, 엽록소-a)과 calanoid 요각류와의 상호 연관성을 파악하였다. 정준대응분석은 대상종의 개체수 자료에 근거하여 유사성 거리를 계산하 여 배열함으로써 출현 양상을 표현함과 동시에 환경요인 과의 상관관계의 모형을 도면화함으로써 상호간의 관계를 이해하는데 적절하며, 도면에서 중심으로부터의 화살 길 이와 방향, 그리고 기울기에 따라 각 출현종과의 상관성 정도가 결정된다. 정준대응분석에 사용된 calanoid 요각류 는 소수출현종에 의한 분석의 과대평가를 낮추기 위해 점 유율 >0.1%, 출현빈도 >30%을 동시에 만족시키는 종들 만을 대상으로 분석하였다. 또한, 환경요인 중 수온과 염 분자료는 수층의 평균값을 사용하였으며, 엽록소-a는 각 수심에서 측정된 값을 적분하여 수층의 대표값으로 사용 하였다. 독립표본 t 검정, 집괴분석, 정준대응분석에 사용 된 모든 자료는 정규분포 가정을 만족시키기 위해 log
e(x+1) 로 변환하여 분석하였다.
3. 결 과
환경요인
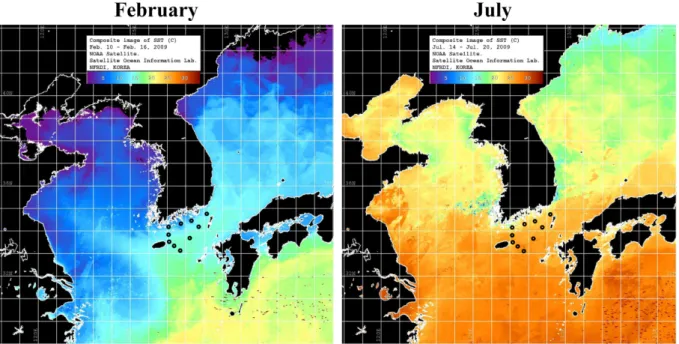
NOAA/AVHRR로부터 분석된 조사해역 표층 수온의
주간 평균 자료를 Fig. 2에 나타내었으며, 분석된 자료에 는 2월과 7월의 조사기간이 포함되어 있다. 2월의 표층 수 온은 남해 연안해역과 제주도 남동쪽 외해역에서 각각 10
oC와 17
oC 내외로 수온 구배가 뚜렷하게 관찰되었다.
조사해역의 7월 표층 수온은 약 21~28
oC 범위로 2월과 같이 외해역에서 연안해역으로 갈수록 낮아지는 양상을 보였다.
CTD 에 의해 관측된 수온과 염분의 변화에 대해 살펴보 면(Fig. 3), 2월의 표층 수온과 표층 염분은 각각 10.9~
16.4
oC와 34.1~34.5 PSU의 범위를 보였으며, 제주도 남동 쪽 외해역(St3-5)의 표층수온은 평균 16.3
oC 로 남해 연안 해역(St8-10)의 평균 12.2
oC와 4.1
oC 의 차이를 나타내었 다. 수온과 염분의 수직 변화 양상은 제주도 인근과 남동 쪽 외해역에 위치한 정점들(St3-7)의 경우 표, 저층간에 차 이를 보이지 않아 수직적 혼합이 활발한 것으로 나타났으 나, 대마도 인근해역(St1, 2)에서는 표, 저층간에 약 3
oC 내외의 차이를 보였다.
7 월의 표층 수온은 19.9~26.7
oC 의 범위로 정점간에 6.8
oC의 차이를 보였으며, 표층 염분 범위는 29.2~32.7 PSU 로 2월에 비해 낮은 염분을 나타내었다. 제주도 인근 과 동남쪽의 해역(St3-7)에서 25
oC 이상의 고수온을 보인 반면에 나머지 정점들에서는 22
oC 이하를 나타내었다.
2월과 다르게 모든 정점에서 수온 및 염분 약층이 형성되 었다. 특히, 대마도 인근의 정점 1의 경우 저층에서 10
oC 내외의 저수온이 관찰되어 다른 정점들과 특징적으로 구 분되었는데, 이는 동해 냉수가 여름철에 대한해협의 서수 도로 유입되어 저층에 수온이 낮은(<10
oC) 냉수가 존재한
Fig. 2. Weekly average sea surface temperature (SST) derived from NOAA/AVHRR during sampling periods in
February and July 2009. Sampling stations are marked as circles
다는 보고(Lim and Chang 1969; Cho and Kim 1998)와 관련성이 있는 것으로 여겨진다. 제주도 인근의 정점 6과 7 에서는 표층에서 20 m까지 30 PSU 이하의 낮은 염분이 관찰되어 저염수의 영향을 시사하였다.
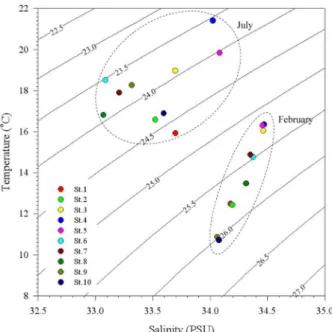
수괴의 특성을 파악하기 위하여 수온과 염분의 수층 평 균 자료를 이용하여 T-S diagram을 작성하였다(Fig. 4) 2월의 모든 조사정점은 저수온(<16.5
oC), 고염분(>34.0 PSU)의 특징으로 7월과 뚜렷이 구분되었다. 2월의 경우
제주도 남동쪽 외해역(St3-5)이 고수온(>16.0
oC), 고염분
(>34.4 PSU) 수괴를 나타내었으며, 남해 연안해역(St9,
10) 은 다른 정점들에 비해 저수온(<10.9
oC), 저염분(<34.1
PSU) 의 수괴 특징을 보였다. 7월에도 제주도 남동쪽 외해
역(St3-5)에서 고수온(>18.4
oC), 고염분(>33.6 PSU) 수괴
특성이 관찰되었고, 제주도 인근과 남해 연안해역(St6-9)
이 중간단계의 수온(>16.3
oC), 저염분(<33.3 PSU) 수괴를,
그리고 대마도 인근과 거제도 외해역(St1, 2, 10)이 저수온
Fig. 3. Vertical profiles of temperature and salinity at each sampling station during February and July, 2009
(<16.3
oC), 중간단계의 염분(>33.4 PSU) 수괴로 각각 구 분되었다.
2월과 7월의 각 정점별 엽록소-a 농도의 수직적 변화 를 Fig. 5에 나타냈다. 2월에는 표층과 저층에서 각각 0.46~1.80 µg l
−1와 0.54~1.79 µg l
−1의 농도 범위를 보였 으며, 수층간의 농도 차이는 0.5 µg l
−1이하로 크지 않았
다. 2월의 표층과 저층의 최대 농도는 각각 거제도 외해역 의 정점 1과 제주도 남동쪽 외해역의 정점 4에서 나타냈 다. 7월의 엽록소-a는 표층과 저층에서 각각 0.15~1.76 µg l
−1와 0.04~1.67 µg l
−1의 농도 범위를 보였으며, 대부분의 정점들에서 수층간에 1.0 µg l
−1이상의 비교적 큰 변화 폭 을 보여 2월과는 다른 양상을 나타냈다. 또한, 제주도 남
Fig. 4. A T-S diagram showing the relationships between temperature (T), salinity (S), and density using a data set of average vertical temperatures and salinity at each sampling station for February and July, 2009
Fig. 5. Vertical profiles of chlorophyll-a at each sampling station during February and July, 2009
Fig. 6. Variations in number of observed species and
Shannon-Wiener diversity index (H') at each
sampling station during February and July, 2009
동쪽에 위치한 정점 4와 5는 각각 수심 50 m와 30 m에서 최대 농도가 관찰되어 표층에서 20 m 사이에서 최대 농 도를 보인 나머지 정점들과 차이를 보였다.
동물플랑크톤
조사해역에서 출현한 동물플랑크톤의 출현종수는 두 조 사월에서 제주도 남동쪽 외해역(St3-5)에서 45종 이상으 로 높았으며, Shannon-Weaver의 다양도 지수 또한 이 해 역에서 2.5 이상으로 다른 정점들과 큰 차이를 보였다 (Fig. 6).
동물플랑크톤의 평균 개체수는 2월(1024.7±570.3 indiv. m
−3)이 7월(457.7±133.9 indiv. m
−3) 에 비해 2.2배 정도 많았다(Table 2). 동물플랑크톤 가운데 요각류 (copepods), 유공충(Foraminiferans), 패충류(ostracods), 미충류(appendicularins), 절지동물 유생(nauplii) 등이 평
균 개체수 10 indiv. m
−3이상으로 주요 분류군이였으며, 이 중 요각류만이 두 조사월간에 유의한 차이(two independent samples t-test, p<0.05)를 보인 것으로 분석 되었다.
요각류가 2월과 7월에 각각 78.6%와 73.1%의 평균 점 유율을 차지하여 동물플랑크톤 분류군 가운데 가장 우점 하였다. 고수온, 고염분의 수괴특성을 보였던 제주도 남동 쪽 외해역(St3-5)에서 요각류의 점유율은 나머지 정점들에 비해 상대적으로 낮았으며, 이러한 경향은 이 정점들(St3- 5) 에서 50% 이하의 점유율을 보였던 7월에 더욱 뚜렷하 였다(Fig. 7). 미충류는 2월과 7월에 각각 10.7%와 6.2%
의 평균 점유율로 요각류 다음으로 높았으며, 2월에는 대 마도 인근의 정점 1, 2와 남해 연안의 정점 9, 10에서 상 대적으로 높은 점유율을 보였지만 7월에는 제주도 남동쪽 외해역(St3-5)에서 높아 대조를 보였다. 그 외에 유공충과 total zooplankton abudance in February and July 2009. Differences between two months were tested by two independent samples t-test (***p<0.001, **p<0.01, *p<0.05, ns: not significant)
Taxa February July
t-test
Mean ± SD RC Mean ± SD RC
Total zooplankton 1024.7 ± 570.3 457.7 ± 133.9 **
Copepods 805.5 ± 431.3 78.6 334.4 ± 135.9 73.1 **
Foraminiferans 23.5 ± 33.4 2.3 22.0 ± 39.9 4.8 ns
Pteropods 0.0 ± 0.0 1.3 ± 2.6 0.3 ns
Ostracods 25.5 ± 22.0 2.5 15.0 ± 20.0 3.3 ns
Hydromedusae 0.6 ± 1.0 0.1 0.5 ± 0.6 0.1 ns
Siphonophorans 2.5 ± 3.9 0.2 0.3 ± 0.7 0.1 *
Chaetognaths 3.9 ± 3.1 0.4 7.7 ± 4.7 1.7 ns
Cladocerans 0.0 0.0 0.3 ± 0.6 0.1 ns
Amphipods 0.2 ± 0.4 <0.1 7.8 ± 10.7 1.7 ***
Isopods 0.0 0.0 <0.0 ± <0.0 <0.1 ns
Decapods 0.2 ± 0.2 <0.1 0.1 ± 0.3 <0.1 ns
Cumaceans <0.0 ± <0.0 <0.1 0.0 0.0 ns
Euphausiids 0.1 ± 0.2 <0.1 0.2 ± 0.3 <0.1 ns
Salpids 0.2 ± 0.3 <0.1 1.2 ± 2.3 0.3 ns
Appendicularians 110.1 ± 136.8 10.7 28.1 ± 32.1 6.2 ns
Doliolids 0.0 0.0 0.5 ± 1.3 0.1 ns
Nauplii 23.0 ± 22.1 2.2 10.0 ± 6.6 2.2 ns
Polychaete larva 2.0 ± 2.4 0.2 1.4 ± 2.0 0.3 ns
Decapod larva 4.5 ± 4.9 0.4 3.5 ± 3.8 0.8 ns
Echinodermate larva 3.3 ± 4.3 0.3 4.3 ± 8.1 0.9 ns
Cirriped larva 1.7 ± 3.5 0.2 2.5 ± 4.5 0.5 ns
Gastropod larva 8.5 ± 7.0 0.8 11.8 ± 18.9 2.6 ns
Bivalve larva 8.5 ± 7.4 0.8 3.3 ± 2.4 0.7 *
Others (larva) 0.8 ± 1.3 0.1 1.3 ± 2.1 0.3 ns
패충류는 두 조사월에서 2-5%의 평균 점유율을 나타내었 고, 다른 조사해역에 비해 제주도 남동쪽 외해역(St3-5)에 서 상대적으로 높은 점유율을 보여 요각류와 반대 양상을 나타냈다.
요각류
2월과 7월, 두 조사월 동안에 출현한 요각류는 calanoids 51 종, cyclopods 4종, harpacticoids 2종, poecilostomatoids 10 종 등을 포함하여 총 34속 67종으로 동정되었다(Table 3). 요각류의 출현종 가운데 Paracalanus parvus s.l.과 Oithona similis 는 2월과 7월에 모든 조사정점에서 출현하 였고(출현빈도 100%), Euchaeta plana는 7월에만 모든 조 사정점에서 출현하였다. 평균 점유율이 3% 이상인 요각 류의 우점종에 대해 살펴보면, 2월에는 P. parvus s.l.
(51.0%), O. similis(16.8%), Oithona plumifera(8.4%), Acartia omorii(5.7%), Paracalanus aculeatus(5.2%),
Corycaeus affinis(4.5%) 등이였다. 7월에는 P. parvus s.l.
(62.1%), Calanus sinicus(7.4%), O. plumifera(6.4%), O.
similis(5.9%), E. plana(3.7%), A. omorii(3.7%) 등이 요각 류의 우점종으로 출현하였다(Table 3).
요각류의 평균 출현 개체수는 2월과 7월에 각각 805.5 indiv. m
−3와 334.4 indiv. m
−3로 독립표본 t 검정에서 두 조사월간에 유의한 차이(p<0.01)를 보였다(Table 2). 요각 류 개체수는 2월에 제주도 남동쪽 외해역의 정점 5에서 247.3 indiv. m
−3로 최소치를 기록했으며, 최대치는 거제 도 외해역의 정점 10(1500.3 indiv. m
−3) 과 대마도 인근의 정점 1(1588.9 indiv. m
−3) 에서 나타냈다(Fig. 7). 7월의 요 각류 개체수는 506.7 indiv. m
−3로 최대치를 보인 제주도 동쪽 해역의 정점 6을 제외한 모든 정점들에서 500 indiv.
m
−3이하의 개체수를 보였다.
2 월과 7월에 출현한 요각류의 주요종들을 대상으로 두 조사월간의 개체수 차이에 대한 유의성 정도와 평균 개체 Fig. 7. Variations in abundance and relative contribution of zooplankton major taxa at each sampling station during
February and July, 2009
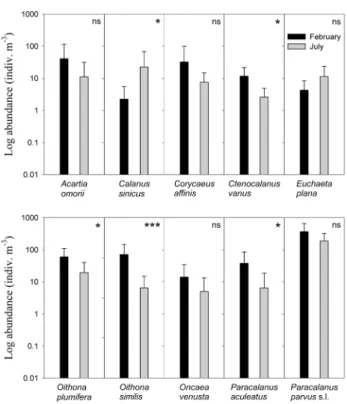
수를 Fig. 8에 나타냈다. C. sinicus의 개체수는 2월에 비 해 7월에 유의하게 높았으며, 반대로 Ctenocalanus vanus, O. plumifera, O. similis, P. aculeatus 등은 7월에 비해 2월에 유의하게 높은 것으로 나타났다. 그러나 A. omorii, C. affinis, E. plana, Oncaea venusta, 그리고 최우점종이 였던 P. parvus s.l., 등의 개체수는 두 조사월 간에 유의한 차이를 보이지 않았다.
군집분석
동물플랑크톤의 출현 개체수에 근거한 Bray-Curtis 유
사도 지수 자료를 이용하여 집괴분석을 실시하였다(Fig.
9). 그 결과, 유사도 지수 60% 수준에서 조사정점들은 2월 과 7월에 각각 3개 Group으로 구분되었다. 2월에는 제주 도 남동쪽 외해역의 정점들(St3-5)과 제주도 동쪽 및 북단 해역의 정점들(St6, 7)이 각각 Group A와 Group B로 구 분되었고, 대마도 인근 해역과 남해 연안해역의 정점들 (St1, 2, 8, 9, 10)이 Group C로 구분되었다. Group A에서 수층의 평균 수온과 염분은 각각 16.2
oC 와 34.5 PSU로 Group B(14.8
oC, 34.4 PSU) 와 Group C(12.0
oC, 34.2 PSU) 에 비해 고수온, 고염분의 환경을 나타냈고, 출현종수와 Korea Strait in February and July 2009. Bold characters represent the most abundant species(relative con- tribution: >3%); and numbers in brackets the occurrence of species in February and July, respectively
Taxa / species Taxa / species Taxa / species
CALANOIDS Clausocalanus sp. (0, 10) Pseudocalanus newmani* (60, 20) Acartiidae Ctenocalanus vanus* (80, 70) Scolecithricidae
Acartia omorii** (50, 50) Eucalanidae Scolecithrx bradyi* (0, 10) Acartia danae* (10, 30) Eucalanus attenuates* (20, 0) Scolecithrix danae* (40, 10)
Acartia sp. (0, 30) Eucalanus bungii var. bungii* (0, 20) Scollecithricella longispinosa* (20, 20) Aetideidae Eucalanus subcrassus* (20, 20) Scollecithricella sp. (50, 20)
Aetideus giesbrechti* (10, 0) Eucalanus mucronatus* (20, 50) CYCLOPOIDS Augaptilidae Eucalanus subtenuis* (80, 10) Oithonidae
Haloptilus longicornis* (0, 10) Rhincalanus cornatus* (50, 10) Oithona plumifera* (60, 80)
Calanidae Euchaetidae Oithona similis** (100, 100)
Calanus sinicus** (90, 90) Euchaeta concinna* (40, 80) Oithona fallax* (0, 20) Canthocalanus pauper* (0, 20) Euchaeta plana* (90, 100) Oithona spp. (30, 50) Cosmocalanus darwini* (20, 20) Euchaeta rimana* (50, 60) HARPACTICOIDS Mesocalanus tenuicornis* (20, 20) Lucicutiidae Clytemnestridae
Nannocalanus minor* (20, 30) Lucicutia flavicornis* (40, 10) Clytemnestra rostrata* (60, 80) Neocalanus gracilis* (40, 30) Lucicutia ovalis* (0, 20) Miraciidae
Undinula vulgalis* (0, 30) Metridinidae Microsetella gracilis* (40, 10) Calocalanidae Pleuromamma gracilis* (50, 10) POECILOSTOMATOIDS Calocalanus pavo* (20, 20) Pleuromamma abdominalis* (20, 0) Corycaeidae
Calocalanus pavoninus *(20, 0) Paracalanidae Corycaeus affinis** (80, 80) Calocalanus plumulosus* (0, 20) Acrocalanus gibber* (50, 70) Corycaeus dahlia* (10, 50) Candaciidae Acrocalanus gracilis* (10, 0) Corycaeus speciosus* (10, 0) Candacia catula* (20, 10) Acrocalanus monachus* (40, 0) Corycaeus pacificus* (0, 20) Centropagidae Delius nudus* (20, 40) Corycaeus spp. (30, 50) Centropages abdominalis** (40, 0) Paracalanus aculeatus* (80, 40) Oncaeidae
Centropages furcatus* (10, 0) Paracalanus parvus s.1. **(100, 100) Oncaea conifer* (40, 70) Centropages tenuiremis** (0, 20) Pontellidae Oncaea mediterranea* (30, 50) Clausocalanidae Labidocera rotunda** (0, 10) Oncaea venusta* (80, 70) Clausocalanus arcuicornis* (70, 20) Labidoecera sp. (0, 10) Oncaea spp. (10, 30) Clausocalanus furcatus* (50, 30) Pontellina plumata* (0, 10) Sapphirinidae
Clausocalanus pergens* (20, 20) Pseudocalanidae Sapphirina stellata* (0, 20)
Fig. 8. Variations in the most abundant copepod species (mean±SD) during February and July, 2009.
Differences between 2 months were tested by the t-test used for 2 independent samples (***p<
0.001, **p<0.01, *p<0.05, ns: not significant)
Fig. 9. Dendrogram of station associations derived from a cluster analysis based on the abundance of zooplankton during February and July, 2009
Table 4. Dominant species contributing > 60% to the relative abundance and mean of species number, diversity index, average temperature and salinity of each cluster during the 2 sampling seasons, calculated using Bray-Curtis cluster analysis (see Fig. 9)
Dominant taxa
Cluster level
February July
Group A Group B Group C Group A Group B Group C
Foraminiferans 10 17
Ostracods 9 8
Appendicularians 16 15
Gastropod larvae 7
Acartia omorii 6 11
Calanus sinicus 10
Corycaeus affinis 5
Oithona plumifera 23 8 5 7
Oithona similis 10 11 13 8
Paracalanus aculeatus 6 5
Paracalanus parvus s.1. 10 43 46 54 68 12
Cumulative contribution (%) 68 62 86 69 87 71
Species number 47 33 31 30 23 46
Diversity index 2.5 1.8 1.9 1.9 1.4 2.7
Average temperature of water column (
oC) 16.2 14.8 12.0 16.6 17.0 19.5
Average salinity of water column (PSU) 34.5 34.4 34.2 33.2 33.4 33.8
서 상대적으로 높았다(Table 4). Group A의 경우 외양해 역에서 주로 출현하는 O. plumifera(23%)가 가장 우점한 반면에 Group B와 Group C에서 연안해역에서 주로 출현 하는 P. parvus s.l.이 40% 이상으로 우점하였다. 또한, Group A에서는 외양해역에서 주로 관찰되는 유공충 (10%), 패충류(9%), P. aculeatus(6%) 등도 비교적 높은 점유율을 차지하여 우점종의 출현양상에서 Group B 및 Group C와 큰 차이를 보였다.
7월의 경우에는 대마도 인근과 제주도 동쪽 및 북단의 정점들(St1, 2, 6, 7, 8)은 Group A로 구분되었고, 남해 연 안해역의 정점들(St9, 10)과 제주도 남동쪽 외해역의 정점 들(St3-5)은 각각 Group B와 Group C로 구분되었다(Fig.
9). Group C에서 수층의 평균 수온과 염분은 각각 19.5
oC 와 33.8 PSU로 나머지 두 Group에 비해 고수온, 고염분 의 특징을 나타냈고, 출현종수와 다양도 지수도 각각 46 종과 2.7로 나머지 두 Group과 큰 차이를 보였다(Table 4). Group A 와 Group B에서는 연안종 P. parvus s.l.가 각 각 54%와 68%의 점유율로 가장 우점하였고, 이 외에도 연안종 C. sinicus(Group A)와 A. omorii(Group B)가 10% 이상의 점유율을 차지하였다. 반면에 Group C의 경 우 17%의 점유율을 보인 유공충이 가장 우점하였고, Group A 와 Group B에서는 출현하지 않거나 낮은 점유율 을 보였던 미충류(15%), 패충류(8%), P. aculeatus(5%) 등 도 높은 점유율을 보여 두 Group과는 다른 양상을 보였다.
Calanoid 요각류의 정준대응분석
Calanoid 요각류 가운데 점유율 0.1% 이상과 출현빈도 30% 이상을 동시에 만족시키는 주요종을 대상으로 정준 대응분석을 실시하여 환경요인과의 상관성과 분포양상을 파악하였다. 2월과 7월의 분석 결과를 살펴보면(Fig. 10), 제 1, 2축의 전체 자료 분산에 대한 누적 기여율은 2월과 7월에 각각 75.3%와 76.5%를 나타냈고 또한, 제 1, 2축에 서 4개의 환경요인(수온, 염분, 수심, 엽록소-a)이 2월과 7 월에 출현한 calanoid 요각류 변화의 92.4%를 설명하였 다(Table 5). 일부 정점(St 6, 7)에서 출현종과의 분포 양상 에서 2월과 7월간에 다소간의 차이를 보이지만 전반적으 로 유사한 분석결과를 나타냈다. 2월과 7월의 분석결과에 서 P. parvus s.l., C. sinicus, A. omorii, Centropages abdominalis 등은 제 1축을 기준으로 오른쪽에 위치하여 왼쪽에 위치한 나머지 종들과 분포 양상에서 대조를 보였 다. 이 4종은 외양역보다는 연안 해역에서 상대적으로 높 은 개체수를 보이는 연안종으로(문 등 2010), 2월과 7월에 공통적으로 대마도 인근 해역(St1, 2)와 남해 연안해역(St 8-10)에 주로 출현하였다. 반면에 제주도 남동쪽 외해역 (St3-5) 에서는 Clausocalanus arcuicornis, Clausocalanus Fig. 10. Ordination plot of the canonical correspondence
analysis (CCA) showing the major calanoid copepods in relation to the hydrographic factors during February and July, 2009. Species labels are: A. dan; Acartia danae, A. omo; Acartia omorii, Aca sp.; Acartia sp., C.abd; Centropages abdominalis, C. arc; Calausocalanus arcuicornis, C. fur; Clausocalanus furcatus, C. sin; Calanus sinicus, C. van; Ctenocalanus vanus, E. con;
Euchaeta concinna, E. pla; Euchaeta plana, E.
sub; Eucalanus subtenuis, N. gra; Neocalanus gracilis, N. min; Nannocalanus minor, P. acu;
Paracalanus aculeatus, P. gac; Pleuromamma gracilis, P. new; Pseudocalanus newmani, P. par;
Paracalanus parvus s.l., S. dan; Scolecithrix
danae, Sco sp.; Scollecithricella sp.
furcatus, Eucalanus subtenuis, Nannocalanus minor, Neocalanus gracilis, Scolecithrix danae, P. aculeatus, Pleuromamma gracilis 등의 외양성 난류종(강 1992;
Kang and Hong 1995)들이 상대적으로 높은 개체수를 보인 것으로 분석되었다. 이 외양성 난류종들은 2월과 7 월의 분석결과에서 수온 및 염분과 양(+)의 상관성을 보 인 반면에 음(−)의 상관성을 보인 연안종들과 대조를 보 였다.
4. 고 찰
수온과 염분의 측정 결과에서 2월(겨울)과 7월(여름)에 조사해역 가운데 제주도 남동쪽 외해역(St3-5)에서 다른 해역에 비해 고수온, 고염분의 수괴가 관찰되어 대마 난류 의 유입을 시사하였다. 대한해협으로 유입되는 대마 난류 의 세기는 계절적으로 가을철에 가장 강하고 겨울철에 가 장 약한 것으로 보고되어 있다(Teague et al. 2002; Guo et al. 2006). Guo et al. (2006) 에 의하면, 대한해협에 영향을 미치는 대마 난류는 타이완해협(Taiwan Strait), 타이완 북 동쪽, 일본 큐슈(Kyushu) 남서쪽의 쿠로시오 지류 등 크 게 3개 경로를 통해 유입되며, 이 3개의 해류들은 계절에 따라 세기를 달리하며 영향을 미치고 있다고 한다. 대한해 협을 통과하는 대마 난류는 가을철에는 남해 연안해역까 지 영향을 미치지만 다른 계절에는 주로 제주도 남단의 외해역을 거쳐 배타적 경제수역의 경계선 하단을 따라 이 동하며 이후 쓰시마섬을 통과하여 동해로 유입되는데
(Kang and Hong 1995), 이러한 요인에 의해 2월과 7월에 제주도 남동쪽 외해역의 수괴가 다른 해역과 특징적으로 구분된 것으로 판단된다.
동물플랑크톤의 종조성과 개체수 자료에 근거한 집괴분 석 결과에서도 2월과 7월에 공통적으로 제주도 남동쪽 외 해역이 하나의 Group으로 구분되어 물리적인 관측 결과 와 일치하였다. 특히, 제주도 남동쪽 외해역에서는 유공 충, 패충류, 요각류 O. plumfera와 P. aculeatus 등이 상대 적으로 높은 점유율을 차지하여 다른 해역과 특징적으로 구분되는 주요한 요인으로 작용하였다. O. plumfera와 P.
aculeatus 은 다양한 수괴가 만나 전선을 형성하는 타이완 인근 해역에서 연구된 바에 의하면 고수온, 고염분의 수괴 특성을 가진 쿠로시오 해류가 유입되는 해역에서 높은 개 체수를 보이는 종들로 알려져 있다(Hsieh et al. 2004;
Liao et al. 2006; Lan et al. 2009). 또한, 대부분의 유공충 은 난류를 선호하며(Kuroyanagi and Kawahata 2004), 패 충류의 출현 양상을 통해 난류의 유입여부를 판단할 수 있는 것으로 보고되어 있다(Tanaka 2008). 따라서 이 분류 군들의 출현 여부와 분포 양상은 대마 난류의 유입과 밀 접한 관련성을 가지는 것으로 판단된다. 제주도 남동쪽 외 해역의 종다양도 지수는 2월과 7월에 2.5 이상으로 다른 해역에 비해 월등히 높았는데, 이러한 결과는 이 해역에 대마 난류가 영향을 미침에 따라 다양한 외양성 난류종들 이 유입되어 동물플랑크톤 군집의 안정성 향상에 기여하 였기 때문이다. 실제로 다양한 수괴가 형성되는 해역에서 종다양도 지수를 비교하면 난류의 영향을 받는 해역에서 Table 5. Summary of canonical correspondence analysis (CCA) for major species of calanoid copepods and hydro-
graphic factors in February and July 2009
Axis-1 Axis-2 Axis-3 Axis-4
February
Eigen values 0.395 0.107 0.029 0.012
Species-environment correlations 0.974 0.943 0.743 0.667 Cumulative percentage variance
of species data 59.3 75.3 79.6 81.5
of species-environment relation 72.7 92.4 97.7 100.0
Sum of all eigenvalues 0.667
Sum of all canonical eigenvalues 0.543
July
Eigen values 0.424 0.170 0.035 0.014
Species-environment correlations 0.974 0.982 0.877 0.772 Cumulative percentage variance
of species data 54.6 76.5 81.0 82.8
of species-environment relation 66.0 92.4 97.8 100.0
Sum of all eigenvalues 0.776
Sum of all canonical eigenvalues 0.643
(Tseng et al. 2008). 특히, 난류를 따라 유입되는 요각류의 외양성 난류성 종에는 Eucalanus, Euchaeta, Undinula 속 (genus) 의 다양한 종들이 포함되는데, 이 속들은 대표적인 대형 요각류 종으로서 먹이 자원량의 측면에서 긍정적인 영향을 미칠 수 있다.
7월 제주도 동쪽 및 북단 해역(St6, 7)에서 수심 20 m까 지 30 PSU 이하의 저염수가 관찰되었고, 이 저염수의 영 향을 받은 것으로 예측되는 남해 동쪽 연안해역(St 8)과 대마도 인근 해역(St1, 2)에서도 표층(0-10 m)에서 31.5 PSU 이하의 저염분으로 다른 해역과 특징적으로 구별되 었다. 저염수가 관측된 해역들은 동물플랑크톤의 집괴분 석에서도 하나의 Group으로 구분되어 다른 해역과 대별 되었고 특히, 요각류 C. sinicus의 평균 개체수와 점유율이 각각 42 indiv. m
−3와 10%로 다른 해역에 비해 매우 높았 다. 남동풍이 우세해지는 여름철에 장강 희석수가 동중국 해 해수와 혼합되어 제주도 방향으로 이동한다는 사실 (Lie et al. 2003) 과 본 조사와 유사한 2009년 여름철에 제 주도 인근 해역까지 장강 희석수가 영향을 미쳤다는 보고 ( 국토해양부 2009)를 고려하면, 7월에 조사해역에서 관찰 된 저염수는 장강 희석수의 유입에 의한 것으로 판단된 다. 따라서 저염수 해역에 집중된 C. sinicus는 장강 희석 수의 주 이동경로인 동중국해 및 황해 남단에서 유입된 것으로 추정해 볼 수 있다. 그러나 이 종은 황해와 동중국 해에서 년 중 출현하지만 여름철에는 연안 해역에서 거의 자취를 감추며 고수온 환경을 극복하기 위해 황해 중앙 및 남단의 저층에 존재하는 황해 저층냉수(Yellow Sea Bottom Cold Water) 에 집중적으로 분포하는 것으로 알려 져 있다(Wang et al. 2003; Pu et al. 2004). 이와 더불어, 장강 희석수는 주로 표층 부분의 염분을 저하시키며 확 산·이동한다는 측면에서, 본 조사해역에서 출현한 C.
sinicus 가 장강 희석수을 따라 유입되었다고 쉽게 단정하 기는 어렵다. 다만, 장강 인근에 용승 해역이 존재하고 이 용승 해역에서는 C. sinicus가 집적되는 현상이 발생함과 더불어 표층부까지 높은 개체수를 유지한다는 점(Wang et al. 2003)에서, 여름철 대한해협 및 남해에 장강 희석수에 의한 C. sinicus의 유입 가능성을 전혀 배제할 수 없다. 결 국, 위에서 언급한 해역(Group A; St1, 2, 6, 7, 8)을 중심 으로 저염분의 특성을 가진 장강 희석수가 여름철에 영향 을 미친 것은 분명하지만, 장강 희석수에 의한 C. sinicus 의 유입 여부는 본 조사의 제한된 자료만으로는 판단하기 어려우며 동중국해, 황해 남단, 제주도 인근해역에 대한 해수 흐름의 특성과 C. sinicus의 분포 양상을 종합적으로 파악하여야만 가능할 것으로 사료된다.
요각류 P. parvus s.l.와 A. omorii는 남해 연안해역에서 우점하는 대표적인 연안종으로 알려져 있으며(심과 노
하는 해역은 남해 연안수의 영향이 강하게 미치는 곳으로 해석할 수 있다. P. parvus s.l.와 A. omorii는 2월과 7월에 공통적으로 높은 점유율로 집괴분석에서 각각 Group C[2 월: 대마도 인근 해역(St1, 2)와 남해 연안해역(St8-10)]와 Group B[7 월: 남해 연안의 중앙 및 동쪽 해역(St9, 10)]로 구분되는데 결정적인 역할을 하였다. 이러한 P. parvus s.l.
와 A. omorii의 분포 양상으로 미루어 보아 남해 연안수의 세력이 7월의 경우 일부 해역에 국한되지만, 2월에는 대마 도 인근 해역(St1, 2)까지 광범위하게 영향을 미친 것으로 추정된다. 이러한 결과는 남해 연안수가 겨울철에는 북서 계절풍과 수표면 열 이동 등에 의해 연안역에서 외해역으 로 확장된다는 보고(나 등 1990; 양 1994)와 일치한다.
요각류 군집 중 calanoid 요각류는 수괴 변화에 매우 민 감하게 반응하며, 외양성 난류종의 경우 고수온, 고염분의 수괴를 선호하여 출현하는 것으로 알려져 있다(강 1992;
Kang and Hong 1995). 2월과 7월, calanoid 요각류의 주 요종에 대한 정준대응분석에서 외양성 난류종은 수온 및 염분은 양(+)의 상관성을 보였으며, 다른 해역에 보다는 제주도 남동쪽 외해역(St 3-5)에서 집중적으로 출현하였 다. 또한, 본 조사에서 출현한 외양성 난류종들 가운데 C.
arcuicornis, C. furcatus, S. danae, P. aculeatus 는 대마 난 류 및 쿠로시오 난류의 유입을 간접적으로 지시하는 지표 종으로서의 가치가 입증된 바 있다(Hsieh et al. 2004;
Hwang et al. 2006). 따라서 calanoid 요각류에 대한 정준 대응분석의 결과, 위에서 언급한 유공충 및 패충류의 출현 양상, 관측된 수온 및 염분의 분포 양상, 그리고 선행 연구 를 종합적으로 고찰해 보면, 2월과 7월의 조사해역 가운 데 제주도 남동쪽 외해역에 대마 난류가 상대적으로 강하 게 영향을 미친 것으로 여겨진다.
결론적으로 2월과 7월의 대한해협에는 대마 난류, 장강 희석수, 남해 연안수 등을 지시하는 수괴가 관찰되었으 며, 동물플랑크톤의 군집 구조와 분포 특성은 수괴 특성의 영향을 강하게 받은 것으로 판단되었다.
사 사
본 연구는 한국해양연구원의 연구사업인 “북서태평양 의 해양환경 변화가 한반도 주변해(대한해협)에 미치는 영 향 연구(PE98731)”와 국토해양부 R&D 과제인 “기후변화 가 남해해양생태계에 미치는 영향 평가 시범연구 (PM56600)” 의 일환으로 수행되었습니다.
참고문헌