대장균을 이용한 Akt/PKB Protein Kinase의 발현 및 활성화
서일대학 식품영양과이재학*
Expression and Activation of Akt/PKB Protein Kinase using Escherichia coli. Lee,Jaehag*. Department of Food and Nutrition, Seoil University, Seoul 131-702, Korea − Among signal transduction systems by protein phosphorylation Akt/PKB protein kinase which is one of serine/threonine kinases, is known to regulate the survival and death of the cell and glucose metabolism. Thus, Akt/PKB protein kinase has been used as one of the target proteins to find anti-cancer agents from natural products. In this study, human Akt/PKB protein kinase was expressed in Escherichia coli expression system for the mass production. Human Akt/PKB protein kinase expressed in E. coli formed inclusion body under the general condition. However, most of the expressed protein was solubilized under the culture temperature at 27oC and 0.01-0.09 mM of IPTG for induc- tion of the protein expression. The expressed protein was purified using Ni2+-NTA agarose column and con- firmed by using anti-Akt antibody. Subsequently, the purified human Akt/PKB protein kinase was activated by in vitro phosphorylation using cellular extract containing kinases. The activated protein was confirmed to phosphorylate the specific fluorescent peptide specially designed as the artificial substrate for Akt/PKB pro- tein kinase.
Key words: Akt/PKB, Escherichia coli expression system, purification, Akt/PKB specific fluorescent pep- tide, anti-cancer agent
서 론
포유동물세포는 외부환경, 세포의 변화, 외부 신호를 인 식하고 이에 대응하여 반응하기 위해 진화학적으로 보존된 여러 신호전달기전들을 가지고 있다. Hemmings 그룹에 의 해 세포생존을 위한 신호전달과정에 Akt/PKB protein
kinase가 중요한 역할을 한다는 사실이 밝혀진 이후, 이의
활성화 조절 기작에 대한 연구가 활발히 진행되고 있다[3].
단순히 하나의 발암유전자로 발견된 Akt/PKB kinase는 해 당 유전자의 염기서열과 단백질 서열의 분석으로, 새로운 phosphatidylinositol 3 kinase(PI3K)의 major effector protein이라는 사실이 밝혀졌다. 이 효소에는 N-terminal에 pleckstrin homolgy(PH) domain이 존재하며, PH 도메인의 존재로 이 효소가 PI3K lipid products에 의해 세포막에 운 반되는 단백질 중의 하나로 밝혀지게 된 것이다[6, 10].
세포내의 신호전달 체계는 생체를 유지하는데 가장 핵심 적인 세포의 생사 및 항상성 등에 필요한 조절을 담당하고 있으며, 이를 조절할 수 있는 조절물질의 개발은 학문적으 로나 산업적으로 중요하다[7, 14, 16]. 이의 중요성을 인식하 여 신호전달 조절물질을 선별하기 위한 연구가 진행된 바 있
으나 큰 성과를 얻지 못하였다. 그 주요 원인은 신호전달의 주요 messenger인 인산화 분석 방법이 너무 어렵고, 고 비 용을 필요로 하며, 시간이 많이 걸리는 어려움을 지적할 수 있다. Akt/PKB kinase의 경우 cell signaling의 상부에 위치 하고 있는 kinase로서 XXRXRXXS/TXX(X: hydrophobic
amino acid) 서열에 대한 특이적인 인산화를 통하여 세포의
생존과 관련된 signal을 관장하고 있다. 이러한 인산화의 표 적 단백질 중에는 세포의 암화에 필수적인 telomerase를 포 함하여, 당대사에 관련된 조절 유전자, 세포사를 유도하는 조절유전자들이 포함되어 있다[6, 10, 13, 15]. 따라서 Akt/
PKB kinase의 활성을 억제하는 저해제를 개발할 경우 세포
내 신호전달체계에 대한 연구용으로 매우 유용하게 작용할 것이며, 항암제 등 다양한 의약품 개발이 가능할 것이다[8, 9, 11, 17]. 이외에도, Akt/PKB kinase가 당뇨병, 노인성 치매 등과도 연관이 있다는 보고들을 염두에 둔다면, Akt/PKB- kinase inhibitor의 의약품으로서의 개발 가능성이 많음을 시 사해 주고 있다. 지금까지 Akt kinase assay에 사용하는 Akt
kinase 단백질은 모두 동물세포에서 생산하였으며, 결과적으
로 상당한 비용을 요구하고 있다. 최근에는 이러한 Akt/PKB kinase를 대체할 수 있는 방선균 유래의 kinase를 이용하여 인산화를 실시하고, 이러한 시스템을 이용하여 Akt/PKB kinase 저해제를 성공적으로 발굴한 예도 있다[4, 17]. 이러 한 Akt/PKB kinase의 중요성을 감안하여, 본 연구에서는 Akt kinase assay에 직접 사용할 수 있는 Akt/PKB kinase를
*Corresponding author
Tel: 82-2-490-7509, Fax: 82-2-490-7507 E-mail: [email protected]
확보하기 위해, Akt/PKB kinase를 동물세포가 아닌 대장균 에서 직접 발현시키고 활성화시키는 연구를 실시하였다.
재료 및 방법
DNA 추출 및 조작
야생형 Akt/PKB kinase 유전자는 pUSEamp-Akt1 cDNA (catalog # 21-153, Upstate Biotechnology사, USA)를 구입 하여 사용하였다. 대장균에서의 형질전환 및 plasmid DNA 재조합 조작은 Maniatis 등의 방법에 따라 추출하였다[12].
각종 제한효소 및 T4 DNA ligase(Takara, Korea) 등은 구 입하여 사용하였으며, 반응조건은 제조 회사의 권장 방법에 따라 실시하였다.
발현벡터의 제작
구입한 pUSEamp-Akt1 cDNA 재조합 벡터를 EcoRI/
BamHI으로 자르고 얻은 1.75-kb의 단편을 같은 제한효소를
이용하여 자른 pET28a(+)(Novagene, USA) 벡터에 삽입하 여 길이 7.1-kb의 pET28-Akt1 재조합 벡터를 제작하였다.
대장균에서 유전자의 대량 발현
pET28-Akt1을 형질전환 시킨 Escherichia coli BL21을 LB 액체 배지에 ampicillin(50 µg/mL)과 함께 접종하여 12 시간 배양한 후 3,000 × g에서 10분간 원심분리하여 균체만 을 모은 후 동량의 LB 액체배지를 이용하여 현탁하였다.
3,000× g에서 10분간 다시 원심분리한 후 LB 배지에 재현
탁시켜 ampicillin(50 µg/mL) 100 µL와 함께 1 mL을 100 mL의 LB 액체 배지에 접종하여 2시간 배양한 후, IPTG를 0.01-0.09 mM을 첨가하여 20-37oC에서 3시간 동안 발현시 킨 다음 3,000 × g에서 20분간 원심분리 후 분석하였다.
단백질의 정량
각각의 배양 조건에서 성장한 균체의 단백질의 양을 측정 하기 위해서 Bradford 분석법을 이용하였다[2]. 배양한 균체 에 각각 300 µL의 완충액을 넣어 현탁시키고, 초음파분쇄기 로 분쇄한 후 10,000 × g에서 30분간 원심분리하여 얻어진 침전물과 supernatant를 이용하여 각각의 protein의 양을 측 정하였는데, 이때 BSA(bovine serum albumin)를 표준품으 로 이용하여 표준검량곡선을 작성한 후 측정하였다.
Akt/PKB kinase의 정제
Akt kinase를 정제하기 위하여, IPTG 0.5 mM로 발현을 유도시킨 대장균을 27°C에서 24시간 배양 후 원심분리하고, 세포만을 disruption buffer [25 mM K2HPO4/KH2PO4(pH 7.5) + 300 mM NaCl, 20 mM imidazole, 0.2 mM CoCl2,
AEBSF]에 현탁하여 초음파로 분쇄하였다. 원심분리로 세포
침전물을 제거한 cell lysate를 nickel-nitrotriacetic acid
(Ni2+-NTA) resin(Qiagen, Germany)과 섞어 1시간 방치하였 다. Ni2+-NTA agarose 컬럼을 동일한 완충액으로 3회 세척 하고, 250 mM imidazole 함유 elution buffer로 His6- tagged protein을 용출하였다. 용출한 단백질은 dialysis buffer [25 mM K2HPO4/KH2PO4, pH 7.5, 0.2 mM CoCl2] 로 충분히 투석하여 사용하였다.
Western blot analysis
분리한 단백질이 목적하는 Akt kinase 임을 명확히 밝히 기 위해서 SDS-PAGE 후 젤에 전개된 단백질을 Pharmacia 사의 multiphore electrophoresis kit를 이용하여 electrotransfer 방법에 의해 nitrocellulose membrane에 옮겼다. Ponceau S staining으로 transfer된 protein band를 확인한 후 1x TBS (Tris-buffered saline: 20 mM Tris, 500 mM NaCl, pH 7.5)으로 washing한 후 blocking buffer(5% dried milk in PBS, 0.05% Tween-20) 100 mL를 넣어 실온에서 3시간 동 안 반응시켰다. 이후, anti-Akt antibody와 1차 반응을 시키 고, Supersignal West Femto Chemiluminescence western blotting detection system(Pierce Biotech, USA)을 이용하여 band의 형성을 확인한 후 건조시켜 보관하였다.
Akt/PKB kinase 활성화
정제한 Akt 단백질은 293 cell lysate를 이용하여 활성화 시켰다. 간단하게, Akt protein kinase(10 unit/µL) 20 µL, 10× kinase buffer(200 mM HEPES [pH 7.4], 143 mM MgCl2, 10 mM EDTA) 3µL, 20 mM DTT 3 µL, cell lysate (1 mg/mL) 1µL, 2 mM ATP 3 µL를 섞어 37oC에서 30분 간 반응 시켰다. 활성화된 Akt/PKB kinase는 필요 시 Ni2+- NTA column으로 정제하여 사용하였다[12].
비방사성 protein kinase assay
Protein kinase assay는 형광 물질인 FITC 가 부착된 기질 (FITC-TRRSRTESIT)을 이용하여 실험하였다. 반응액은 FITC-peptide 5µg, 2 × reaction buffer(40 mM HEPES [pH 7.2], 20 mM MgCl2, 20 mM MnCl2, 2 mM DTT, 40 mM ATP, 2µg phosphatidyl serine) 10 µL에 증류수를 혼합하여 총 부피를 20 µL로 조정하고, 30oC에서 30분 반 응한 후 95oC에서 10분간 열처리로 반응을 정지시킨 sample 을 0.8% agarose gel에서 전개하여 활성을 확인하였다[4].
결과 및 고찰
세포는 세포 밖의 신호(signal)를 세포내로 전달해주는 다 양한 신호전달기구에 의존하여 생명현상을 유지한다[1, 5].
그 중 세포내의 작은 2차 신호전달 물질(second-messenger molecules)은 중요한 역할을 하는데, 특히 inosites는 kinase 와 반응하는 것으로 알려져 있다. Growth-factor regulated
serine/threonine kinase인 protein kinase B(Akt/PKB)는 phosphoinositide 3-OH kinase(PI-3K) product와 결합하여 plasma membrane으로 translocation되며 phosphoinoside- dependent kinase 1(PDK1) 등에 의하여 473번째 위치의 serine과 308번째 위치의 threonine이 인산화 됨으로써 활성 화(activation)된다[3]. 이렇게 활성화 된 Akt는 다양한 스트 레스에 의해서 야기되는 apoptosis로부터 세포를 보호하는 survival signal을 전달해주며 insulin과 관련된 대사과정에 참여하여 중요한 역할을 하는 것으로 밝혀졌다. 따라서 이 와 같은 Akt kinase 활성을 조절 할 수 있는 조절물질은
Akt kinase가 관여하는 다양한 조절 기능으로부터 유추할 때
암, 당뇨, 치매 등을 치료할 수 있는 신약 개발에 많은 기여 를 할 수 있을 것으로 판단된다. 따라서 세포내 신호전달 기 구 중 Akt등을 포함하는 인산화를 통한 신호전달기구를 분 석할 수 있는 assay kit개발을 목적으로 human Akt kinase 를 활성이 상실되지 않은 형태로 대장균에서 대량으로 발현 시키는 연구를 실시 하였다.
Human Akt/PKB의 대장균에서의 발현
대장균 발현 vector인 pET28a(+)에 human Akt/PKB의
cDNA 유전자를 삽입한 pET28-Akt1 재조합 벡터를 제조한
후, 이를 E. coli BL21에 형질전환하였다. 형질전환체 접종 시 IPTG를 최종농도 0.5 mM이 되도록 첨가하여 37oC에서 1시간 배양한 후 cell lysate를 SDS-PAGE로 확인해 본 결과
Akt/PKB로 추정되는 단백질이 발현됨을 알 수 있었다(Fig.
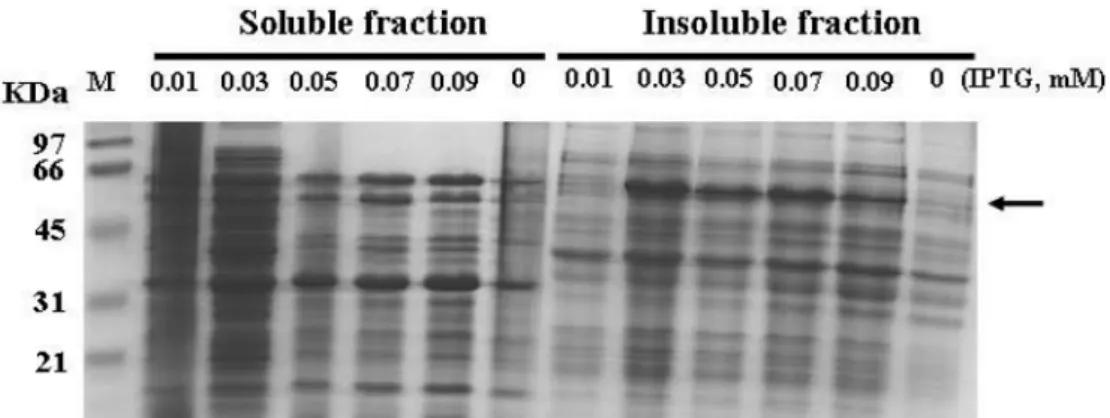
1). 예상되는 발현 단백질은 60 kDa 정도이지만 발현된 단 백질의 분자량은 이 보다 약간 큰 것으로 분석되었다. 하지 만 상기 조건에서 발현된 단백질은 soluble fraction이 아닌 insoluble fraction으로 발현되어 보다 구체적인 발현 조건 개 선이 요구되었다(data not shown).
Akt/PKB 발현조건 최적화
Akt/PKB를 soluble form으로 대량 발현시키기 위해서
IPTG 농도를 바꾸어주거나 배양온도를 낮추어 주는 등 다
양한 조건에서 형질전환 된 E. coli를 배양하는 실험을 실 시하였다. 전반적으로는 27oC에서 3시간 배양 하였을 때 가 용성 발현 단백질 양이 많았다. 따라서 다양한 IPTG 농도 하에서 27oC에서 3시간 발현시킨 Akt kinase의 발현 양상 을 Fig. 2에 나타냈다. 결과적으로, IPTG가 0.01-0.09 mM 까지 Akt 단백질이 soluble form으로 발현되었으나, in- soluble form도 상당량 존재하였다. 최종적으로 0.03-0.07 mM의 IPTG 농도에서 soluble form으로의 발현율이 가장 높은 것으로 판단되었으며, 이후의 모든 발현 실험은 0.05 mM IPTG 농도로 27oC에서 3시간 발현을 유도하여 진행 하였다.
Fig. 1. Detection of the expressed protein in E.coli BL21 trans- formed with pET28-Akt1. The total cellular protein of E. coli was analyzed by SDS-polyacrylamide gel elctrophoresis. lane M, molecular weight standards; lane 1, sample from E. coli carry- ing pET28a(+); lane 2, sample from E. coli carrying pET28-Akt.
The overexpressed protein with a molecular weight of 60 kDa is indicated by the arrow.
Fig. 2. SDS-PAGE analysis of the soluble (left) and insoluble (right) cellular protein prepared from E. coli BL21(pET28-Akt1). Lane 1 represents the low molecular weight size marker. E. coli cultures were incubated at 27oC. The final concentration of IPTG used for induc- tion was indicated in each lane and the band for Akt kinase is marked by the arrow.
발현된 Akt/PKB의 정제
발현된 Akt kinase는 Ni2+-NTA agarose column을 사용 하여 정제하였다(Fig. 3). 예상대로 insoluble form의 Akt 단 백질은 제시된 컬럼 조작 조건에서 분리되지 않았으나, soluble Akt만이 성공적으로 분리되었다. 분리된 단백질은 예 상 하는 분자량 보다 약간 큰 것으로 생각되었지만, anti-Akt antibody를 이용한 Western blot 결과 human Akt/PKB임을 확인할 수 있었다.
Akt/PKB의 in vitro 활성화 및 nonradioactive kinase assay
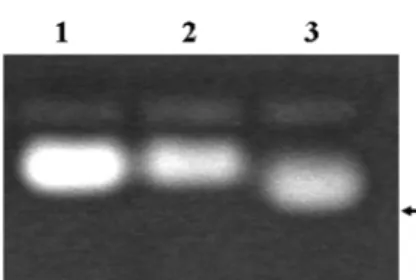
E. coli에서 발현 되어 정제된 Akt/PKB는 473번째 위치 의 serine과 308번째 위치의 threonine이 인산화 되지 않았 으며, 따라서 kinase 활성을 나타내지 않는다. 활성형 Akt/
PKB를 얻기 위하여 정제된 Akt/PKB를 293 cell lysates와 반응시켜 인산화 시킨 후 다시 정제하여 활성형 Akt/PKB를 얻을 수 있었다. 이렇게 얻어진 활성형 Akt kinase가 이전에 보고된 형광 peptide를 이용한 nonradioactive kinase assay system에 이용될 수 있는 지 확인하였다(Fig. 4). 그 결과, 활 성화되지 않은 Akt 단백질에서는 peptide의 인산화가 관찰 되지 않았지만, 활성화 된 Akt 단백질에서는 peptide를 인산 화 시킬 수 있음이 관찰되었다. 이러한 사실은 대장균에서 발현시켜 얻은 Akt kinase로도 nonradioactive kinase assay 의 kinase source로 사용할 수 있음을 명백하게 보여 주었다.
일반적으로 kinase의 활성 측정을 위해 단백질 기질이나 펩 타이드에 방사성 인산이 전달되는 정도를 측정하거나 인산화 잔기에 특이적인 항체를 이용하지만, 이러한 방법들은 위험
성이 높고, 시간이 오래 걸리며, 고가여서 대량 assay에 적용 하기는 불가능하다. 그러나, 상기에서 설명한 nonradioactive Akt protein kinase의 활성 측정법은 방사성 동위원소를 사 용하지 않고도 감도가 뛰어나고 신속하게 2시간 이내에 Akt protein kinase의 활성을 측정할 수 있다[4, 6]. Akt protein
kinase가 기질로 이용하는 형광 펩타이드를 인산화시키면 펩
타이드의 전하가 −2가 만큼 환원되며, 이러한 전하량의 변 화에 따라 인산화된 펩타이드와 인산화되지 않은 펩타이드 가 중성 pH상태의 agarose gel 전기 이동 양상이 달라지게 된다. 즉, 인산화된 펩타이드는 양극으로 빠른 속도로 이동 하지만 인산화되지 않은 펩타이드는 중성을 띄어 이동하지 않거나 천천히 이동하게 된다. 이러한 방법을 이용하여 Akt protein kinase에 특이적인 저해제를 식물로부터 분리하여 구 조 및 효능을 보고한 논문이 있으나, 모든 assay에는 반드시 kinase source가 필요하며, 현재 시판되는 Akt kinase를 사 용하기에는 너무 고가이다[17]. 본 연구 결과에서 제시하고 있는 것처럼 Akt kinase를 대장균에서 발현시키고 이를 assay에 사용한다면, 학문적으로는 물론 다양한 Akt kinase inhibitor 탐색에 많은 도움이 될 것으로 사료된다.
요 약
단백질 인산화를 통한 세포내 신호전달기구 중 serine/
threonine kinase에 속하는 Akt/PKB는 세포 생존과 사멸, 당 대사 등을 조절하는 것으로 알려져 있다. 이러한 이유로,
Akt/PKB 단백질은 천연물질들로부터 항암제를 탐색하기 위
한 한 가지 target으로 사용되어 왔다. 본 연구에서는 Akt/
PKB 단백질을 대량으로 생산하기 위하여 대장균의 단백질 발현 시스템을 이용하여 human Akt/PKB 단백질을 발현시 켰다. 대장균에서 대량 발현된 Akt는 일반적인 조건에서는 inclusion body를 형성하였다. 배양온도 27oC에서 0.01-0.09 mM IPTG로 발현 유도 시 발현된 human Akt/PKB 단백질 Fig. 4. Phosphorylation of the FITC-labeled synthetic peptide by the purified Akt kinase. The Akt kinase was activated and used for a nonradioactive protein kinase assay as described in materials and methods. Lane 1 represents the substrate in dis- tilled water used in this study, and lane 2 and 3 represent the reac- tions with the intact Akt kinase (lane 2) and the activated Akt kinase (lane 3), respectively. The arrow indicates the phosphory- lated peptide that migrates faster than that of nonphosphotrylated form on agarose gel electrophoresis.
Fig. 3. Purification of Akt kinase by Ni2+-NTA affinity column chromatography. SDS-PAGE of the proteins during purifica- tion step (left) and Western blot analysis with anti-Akt mono- clonal antibodies (right) were performed. lane M, molecular weight standards; lane 1, insoluble fraction of cell-free lysate from E. coli BL21(pET28-Akt1); lane 2, passed proteins of insoluble fraction from Ni2+-NTA column; 3, effluent of insoluble fraction from Ni2+-NTA column; lane 4. soluble fraction of cell-free lysate from E. coli BL21(pET28-Akt1); lane 5, passed proteins of solu- ble fraction from Ni2+-NTA column; lane 6-8, washed solution from Ni2+-NTA column; 9, effluent of soluble fraction from Ni2+- NTA column; W, Western blot analysis of the purified protein (lane 9) using anti-Akt antibody. The band for Akt is indicated by the arrow.
상당 부분이 가용화 되었다. 발현된 Akt kinase를 Ni2+-NTA agarose column으로 정제하고, anti-Akt antibody를 이용하 여 정제된 단백질이 Akt kinase 임을 확인하였다. 정제된
human Akt/PKB 단백질은 세포추출물에 존재하는 인산화 단
백질을 이용하여 in vitro에서 인산화 되었으며, 인산화된 활 성형 human Akt/PKB protein kinase는 human Akt/PKB protein kinase 특이 형광 peptide를 특이적으로 인산화하였다.
감사의 글
본 논문은 2008년도 서일대학 학술연구비에 의해 연구되 었음.
REFERENCES
1. Bourret, R. B., K. A. Borkovich, and M. I. Simon. 1991.
Signal transduction pathways involving protein phosphorylation in prokaryotes. Annu. Rev. Biochem. 60: 401-441.
2. Bradford, M. M. 1976. A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Anal. Biochem. 72: 248- 254.
3. Fayard, E., L. A. Tintignac, A. Baudry, and B. A.
Hemmings. 2005. Protein kinase B/Akt at a glance. J. Cell Sci. 118: 5675-5678.
4. Her, J.-H., Y. H. Cheong, J.-H. Kim, S.-K. Sin, C. G. Hyun, J. Chun, S. S. Kang, D.-K. Kang, and S.-K. Hong. 2002.
Identification of a protein kinase using a FITC-labelled synthetic peptide in Streptomyces griseus IFO13350. Kor. J.
Microbiol Biotechnol. 30(3): 235-240.
5. Hong, S.-K., A. Matsumoto, S. Horinouchi, and T. Beppu.
1993. Effects of protein kinase inhibitors on in vitro protein phosphorylation and cellular differentiation of Streptomyces griseus. Mol. Gen. Genet. 236: 347-354.
6. Kang, S. S., T. Kwon, D. Y. Kwon, and S. I. Do. 1999. Akt protein kinase enhances human telomerase activity through phosphorylation of telomerase reverse transcriptase V subunit. J. Biol. Chem. 274: 13085-13090.
7. Kase, H., K. Iwahashi, Y. Matsuda, M. Takahashi, C.
Murakata, A. Sato, and M. Kaneko. 1987. K-252a com- pounds, novel and potent inhibitors of protein kinase C and cyclic nucleotide-dependent protein kinases. Biochem.
Biophys. Res. Commun. 142: 436-440.
8. Ko, J. H., S. W. Yeon, H. S. Lee, T. Y. Kim, D. Y. Noh, K.
S. Shim, S.-K. Hong, and S. S. Kang. 2004. Inhibitory effects of dithiolo-thione derivative SWU-20009 on Akt activity. J. pharmaceutical society of Korea 48(2): 105-110.
9. Kondapaka, S. B., M. J. Zarnowski, D. R. Yver, E. A.
Sausville, and S. W. Cushman. 2004. 7-Hydroxystaurosporine (UCN-01) inhibition of Akt thr308 but not ser473 phos- phorylation: a basis for decreased insulin-stimulated glucose transport. Clinical Cancer Res. 10: 7192-7198.
10. Kwon, T., D. Y. Kwon, J. Chun, J. H. Kim, and S. S. Kang.
2000. Akt protein kinase inhibits Rac1-GTP binding through phosphorylation at serine 71 of Rac1. J. Biol. Chem. 275:
423-428.
11. Lee, H. Y., S. H. Oh, J. K. Woo, W. Y. Kim, C. S. Van Pelt, R. E. Price, D. Cody, H. Tran, J. M. Pezzuto, R. M. Moriaty, and W. K. Hong. 2005. Chemopreventive effects of deguelin, a novel Akt inhibitor, on tobacco-induced lung tumorigenesis. J. Natl. Cancer Inst. 97: 1695-1699.
12. Maniatis, T., E. F. Fritsch, and J. Sambrook. 1982. Molecular Cloning: A Laboratory Manual. Cold Spring Harbor Laboratory Press, Cold Spring Harbor, New York, U. S. A.
13. Paz, K., Y.-F. Liu, H. Shorer, R. Hemi, D. LeRoith, M.
Quan, H. Kanety, R. Seger, and Y. Zick. 1999. Phos- phorylation of insulin receptor substrate-1 (IRS-1) by protein kinase B positively regulates IRS-1 function. J. Biol. Chem.
274: 28816-28822.
14. Sharma, S. V., T. Agatsuma, and H. Nakano. 1998. Targeting of the protein chaperone, HSP90, by the transformation suppressing agent, radicicol. Oncogene 16: 2639-2645.
15. Takeuchi, H., Y. Kondo, K. Fujiwara, T. Kanzawa, H. Aoki, G. B. Mills, and S. Kondo. 2005. Synergistic augmentation of rapamycin-induced autophagy in malignant glioma cells by phosphatidylinositol 3-kinase/protein kinase B inhibitors.
Cancer Res. 65: 3336-46.
16. Uehara, Y. and H. Ogawara. 1991. Use and selectivity of herbimycin A as inhibitor of protein tyrosine kinases.
Methods Enzymol. 201: 370-379.
17. Yang, H. Y., H.-S. Lee, J.-H. Ko, S.-W. Yeon, T.-Y. Kim, B.
Y. Hwang, S. S. Kang, J. Chun, and S.-K. Hong. 2006.
Identification of 3'-hydroxymelanetin and liquiritigenin as Akt protein kinase inhibitors. J. Microbiol. Biotechnol. 16:
1384-1391.
(Received March 13, 2009/Accepted April 16, 2009)