사람 치은섬유모세포에서 NF- κ B와 JNK 활성 억제를 통한 돌김 에탄올 추출물의 항염증 효과
박충무1, 윤현서2*
1동의대학교 임상병리학과 조교수, 2동의대학교 치위생학과 부교수
Anti-inflammatory effect of Porphyra yezoensis ethanol extract through the inhibited NF-κB and JNK activation in LPS-PG stimulated HGF-1
cells
Chung-Mu Park
1, Hyun-Seo Yoon
2*1Dept. of Clinical Laboratory Science, Dong-Eui University
2Dept. of Dental Hygiene, Dong-Eui University
요 약 사람치은섬유모세포(human gingival fibroblast, HGF)는 치은조직에 존재하는 주요한 세포의 형태 중 하나로 외부 자극에 반응하여 다양한 염증대사물질을 생산한다. 본 연구에서는 돌김에탄올추출물(PYEE)이 Porphyromonas gingivalis 로부터 분리한 lipopolysaccharide로 염증이 유도된 HGF-1 cell에서 항염 효과를 보이는지 분석하고자 하였다. LPS-PG 에 의해 과발현된 iNOS와 COX-2는 PYEE의 처리에 의해 농도 의존적으로 발현이 감소되었고, nuclear factor (NF)-κB 또한 동일한 양상으로 활성이 억제되었다. 신호전달물질 중 c-Jun NH2-terminal kinase (JNK)의 인산화만이 PYEE에 의해 억제되었다. 그리고 항염 작용에 관여하는 것으로 알려진 2상 효소 중 하나인 NAD(P)H:quinone dehydrogenase (NQO)-1 도 분석하였고, 이 효소는 PYEE의 처리에 의해 강하게 발현이 유도가 되었다. 결론적으로 돌김에탄올추출물은 치주질환 에방과 치료를 위한 후보물질로 활용 가능할 것으로 사료된다.
주제어 : 돌김추출물, 염증, Mitogen-activated protein kinase, Nuclear factor kappa B
Abstract Human gingival fibroblast (HGF) is the main cell type existed in periodontium and produces a variety of inflammatory mediators by external stimuli. In this study, the anti-inflammatory activity of Porphyra yezoensis ethanol extract (PYEE) on LPS-PG lipopolysaccharide from Porphyromonas gingivalis activated HGF-1 cell. Up-regulated iNOS and COX-2 expressions by LPS-PG were significantly attenuated by PYEE treatment in a dose-dependent manner. In addition, activated nuclear factor (NF)-κB was also dose-dependently inhibited by PYEE treatment. Among upstream signaling molecules, PYEE treatment inhibited phosphorylation of c-Jun NH2-terminal kinase (JNK) but did not give any effect on other molecules. On the other hand, one of phase II enzymes, NAD(P)H:quinone dehydrogenase (NQO)-1, was analyzed due to its anti-inflammatory activity, which was upregulated by PYEE treatment. Consequently, PYEE could be candidates for the prevention and treatment of periodontal diseases.
Key Words :Inflammation, Mitogen-activated protein kinase, Nuclear factor kappa B, Porphyra yezoensis
*This work was supported by Dong-eui University Grant.(201802740001)
*This work was supported by the National Research Foundation of Korea (NRF) grant funded by the Korea government (MSIT) (NRF-2017R1C1B5017311)
*Corresponding Author : Hyun-Seo Yoon ([email protected]) Received September 11, 2018
Accepted December 20, 2018
Revised October 16, 2018 Published December 28, 2018
1. 서론
염증반응은 물리적 자극이나 미생물의 감염 등으로 인해 발생하는 손상을 복구하기 위한 조직의 수복반응을 말한다[1]. 발생하는 염증의 대부분은 급성에서 치유되나 그 중 일부는 만성으로 이환되고, 이러한 만성염증은 조 직손상을 촉진하여 암으로 진행되기도 한다. 따라서 염 증반응이 발생하면 초기에 완화 및 제거하는 것이 중요 하고 이를 위해 많은 항염증제가 사용이 되고 있으나 부 작용이 많이 발생하고 있다[2]. 따라서 부작용이 발생하 지 않는 천연물 유래 물질들이 항염증 후보물질로 많이 분석되고 있다[3].
치주 질환은 치은조직에 국한된 가벼운 손상인 치은 염(gingivitis)과 치아지지조직까지의 손상을 포함하는 치주염(periodontitis)으로 구분할 수 있고, 구강내에 존 재하는 여러 종류의 세균 군에 의해 발생하며 특히 Porphyromonas gingivalis가 가장 주요한 원인균인 것 으로 알려져 있다[4,5]. P. gingivalis는 그람 음성 혐기성 간균으로 치주질환이 발생한 병소에서 증가하고 열구상 피에 침투하며 이 과정에서 P. gingivalis가 생산하는 대 사산물이나 독소, 특히 염증성 cytokine에 의해 치주조직 이 상해를 입게 된다[6,7]. P. gingivalis의 감염에 의해 발생한 염증은 치주조직의 파괴와 치조골의 재흡수를 유 발하고 결국 치아의 상실로 이어지게 된다.
세균 세포벽의 구성 성분인 lipopolysaccharide (LPS) 는 숙주 방어 체계를 자극하여 염증 반응을 시작하는 인 자 중 하나이다[8-10]. 염증 반응은 주로 면역세포에 의 해 진행되지만 치은 섬유모세포(human gingival fibroblast, HGF)와 같이 LPS의 자극에 대항하여 숙주 방어 체계에 관여하는 세포도 있다[11]. 특히, 치은 섬유 모세포는 P. gingivalis로부터 분리한 LPS (LPS-PG)에 대하여 extracellular signal-regulated kinase (ERK), c-Jun NH2-terminal protein kinase (JNK)와 p38 mitogen activated protein kinase (MAPK)과 같은 세포내 신호전 달물질을 조절하여 염증 매개 인자인 interleukins (ILs), tumor necrosis factor (TNF)-α, inducible nitric oxide synthase (iNOS)와 cyclooxygenase (COX)-2의 발현에 관여하는 것으로 알려져 있다[12]. 그러므로 치주질환의 유발과 진행에 밀접히 관련된 이들 염증 매개 인자들의 활성 조절이 치주조직의 염증 반응을 억제하는데 중요한 과정이라고 할 수 있다.
NAD(P)H:quinone dehydrogenase (NQO)-1은 세포
질 내 존재하며 항산화작용을 통해 세포보호작용을 하는 효소 중 하나이다. 이 효소는 퀴논의 대사과정에서 반응 성이 높은 세미퀴논이 아니라 독성이 적은 하이드로퀴논 으로 변환을 함으로써 세포보호 능력을 보이는 것으로 알려져 있다[13].
섬유질, 단백질, 미네랄과 비타민이 풍부한 반면 지방 함량은 낮은 해조류는 열량이 낮고 풍부한 영양성분을 가져 중요한 대안식품 중 하나로 생각되고 있다[14]. 해 조류는 항산화물질이 풍부하고, 다양한 생리활성 성분으 로 인해 소장·대장암, 피부암, 유방암 동물 실험계를 통해 항암활성 또한 있는 것으로 알려졌다[15]. 그 중 미역, 다 시마와 함께 식용으로 널리 이용되고 있는 김(Porphyra) 은 우리나라의 서해, 남해에서 광범위하게 양식되고 있는 보라털과의 해조류로 심혈관계에 좋은 eicosapentaenoic acid (EPA)와 천연 산화방지 성분이자 항노화, 항암 효 과를 보이는 클로로필, 카로티노이드 및 피코빌린 등의 색소 성분도 포함하고 있다[14,16]. 영양이 풍부하고 활용 가치가 높은 식품인 김은 여러 방면의 연구를 통해 그 생 리활성이 확인되었고, 특히 항염증과 항산화 분야에 있 어서도 그 활성이 분석되었으나 대부분의 연구는 생쥐 대식세포를 이용한 실험에 국한되어 있다[17,18]. 최근 치 은 섬유모세포(HGF-1)을 이용하여 Curcuma xanthorrhiza supercritical 추출물 (CXS)의 항염 효과가 검증되었으나 이도 HGF-1 cell 단독이 아닌 대식세포를 함께 진행하였 다[19]. 본 연구에서는 돌김 추출물이 사람 치은 섬유모 세포의 염증 활성에 대한 억제 효과를 분석하여 치주염 완화능에 대한 기초자료를 제공하고자 하였다.
2. 연구방법
2.1 연구대상
2.1.1 세포배양 및 시료
실험에 사용한 사람 치은섬유모세포(human gingival fibroblast, HGF-1 cell line)는 American Type Culture Collection (ATCC, CRL-2014; Rockville, MD, USA)에 서 분양받았고, 10%의 fetal bovine serum (FBS, Hyclone, South Logan, UT, USA), 100 Unit Penicillin/Streptomycin (Hyclone)이 포함된 Dulbecco’s modified eagle’s medium (DMEM) 배지를 사용하여 3 7℃, 5% CO2가습된 조건에서 배양하였다. 돌김 에탄올
추출물(Porphyra yezoensis ethanol extract, PYEE)은 제주생물다양성연구소(Jeju, Korea)에서 분양받았다. P.
gingivalis에서 분리한 LPS (LPS-PG)는 Invivogen (San Diego, CA, USA)으로부터, dimethyl sulfoxide (DMSO), sodium dodecyl sulfate (SDS)는 Sigma-Aldrich (St.
Louis, MO, USA)로부터 구입하였다. 1차 항체인 iNOS, COX-2, phospho-p65, NQO-1, phospho-Akt, phospho-ERK, phospho-JNK, phospho-p38과 2차 항체인 horseradish peroxidase (HRP)-conjugated anti-rabbit IgG 항체는 Santa Cruz Biotechnology (Dallas, TX, USA)와 Cell Signaling Technology (Boston, MA, USA)에서 구입하 여 분석에 사용하였다.
2.2 연구방법
2.2.1 세포독성 평가
세포생존율은 EZ-Cytox cell viability assay kit (Daeil Lab. service, Seoul, Korea)을 사용하여 측정하였 다. HGF-1 cell을 24 well plate에 1×105cell/well의 농도 로 파종한 후 24시간동안 배양하고 PYEE 시료를 농도별 로 처리하였다. 그리고 2시간 후 LPS-PG를 1 μg/ml의 농도로 처리하고 다시 20시간 배양하였다. 그 후 EZ-Cytox 시약 10 μl를 첨가하여 1시간 동안 배양한 후 microplate reader (Benchmark Plus, Bio-Rad, Hercules, CA, USA)로 450 nm에서 흡광도를 측정하였다.
2.2.2 Western blot 분석
HGF-1 cell을 100-mm dish에 5×106cells/dish의 농도 로 파종하고 24시간동안 부착한 후 PYEE를 농도별로 처 리하였다. 2시간 후 1 μg/ml의 농도로 LPS-PG를 처리하 고 다시 24시간동안 배양하였다. 상기의 방법으로 처리 된 세포는 염증매개물질(iNOS와 COX-2)의 분석에 이용 하였다. 그리고 파종된 세포에 PYEE시료와 LPS-PG (1 μg/ml)를 동시에 처리하고 4시간동안 배양된 세포는 전 사인자와 신호전달물질의 분석에 사용하였다. 시료 처리 가 끝난 세포는 PBS로 2회 세척하고 0.5 ml의 단백질 추 출 용액(PRO-PREP, Intron Biotechnology, Seongnam, Korea)으로 수확한 후 얼음에 10분 동안 정치하여 세포 를 용해하였다. 용해된 세포가 포함된 단백질 추출 용액 은 13,000 ×g에서 5분동안 원심분리한 후 분리된 상층을 새 튜브로 옮긴 후 분석에 사용하였다. 단백질 농도는 Bradford법으로 정량하였다. 분리한 단백질 25 μg이 포
함된 시료를 준비한 후 10% SDS-polyacrylamide gel에 전기영동 후 polyvinylidene fluoride membrane (PVDF, Bio-Rad)으로 이동시켰다. 단백질이 이동된 PVDF membrane은 5% non-fat dry milk를 TBST에 녹인 용 액으로 1시간동안 실온에서 블로킹을 진행하였다. 그 후 1:100-1,000의 비율로 희석한 1차 항체와 섞은 후 4℃에 서 24시간동안 보합반응을 하였다. 1차 항체와의 보합반 응이 끝난 PVDF membrane은 TBST로 3회 세척 후 다 시 1:1,000으로 희석한 2차 항체와 실온에서 2시간동안 보합반응을 하였다. 2차 항체까지 반응이 끝난 후 enhanced chemiluminescence solution (ECL, Santa Cruz Biotechnology)을 이용하여 반응을 유도한 후 ECL sensitive film에 감광시켜 단백질의 발현 변화를 측정하 였고 Gel Doc EQ system (Bio-Rad)으로 정량 분석하였다.
2.3 분석방법
3회 반복 시행한 모든 실험 결과는 SPSS 통계 프로그 램(version 25.0, SPSS Inc., Chicago, IL, USA)을 이용하 여 평균 ± 표준편차(mean ±SD)로 나타내었다. 실험군 간의 유의성 검증에는 일원배치 분산분석(one-way ANOVA)을 이용하였고, Duncan's multiple range test 방법으로 사후 검증을 시행하였다.
3. 연구결과
3.1 LPS-PG로 유도된 HGF-1 cell의 염증에서 PYEE의 iNOS와 COX-2 억제 효과
PYEE가 HGF-1 cell에 독성을 미치는지 여부를 먼저 확인한 결과, 본 실험에서 사용한 50, 100, 200, 400 μg/ml 의 농도 범위에서는 세포생존율의 변화가 일어나지 않는 것을 확인할 수 있었다 (data not shown). 그리고 PYEE 가 LPS-PG에 의해 유도된 HGF-1 cell의 염증 억제 효 과를 가지는지의 여부를 확인하기 위하여 iNOS와 COX-2의 유전자 발현량을 분석한 결과 Fig. 1에서 보는 바와 같이 LPS-PG에 의해 발현량이 증가하였고, PYEE 를 50, 100, 200, 400 μg/ml의 농도로 처리했을 때 농도 의존적으로 감소하는 것을 확인할 수 있었다. 이 결과로 미루어보아 PYEE는 LPS-PG에 의해 유도된 HGF-1 cell의 염증 반응을 억제할 수 있는 능력이 있음을 알 수 있었다.
Fig. 1. PYEE inhibited protein expression levels of iNOS and COX-2 in LPS-PG stimulated HGF-1 cells. Panel A shows protein expression levels of iNOS and COX-2 by PYEE treatment. All signals were normalized to protein levels of actin, an internal control, and expressed as a ratio (Panel B).
Data represent the mean±SD of triplicate experiments. Values sharing the same superscript are not significantly different at p<0.05 by Duncan’s multiple range test.
3.2 LPS-PG로 유도된 HGF-1 cell의 염증에서 PYEE에 의한 NF-κB와 JNK의 활성 억제 효과 LPS-PG에 의해 유도된 iNOS와 COX-2의 과발현은 염증 전사인자인 NF-κB에 의해 매개된다. 이 염증 전사 인자는 세포질 내에서 불활성의 형태로 존재하다가 NF- κB의 subunit 중 p65가 인산화되어 활성화되면 핵 내로 이동하여 염증매개인자들의 합성을 위한 유전자를 전사 하게 된다. 본 실험에서는 PYEE가 LPS-PG로 유도된 NF-κB의 구성단백질인 p65의 인산화와 그 상위신호전 달체계인 MAPK와 phosphoinositide 3-kinase (PI3K)/Akt를 western blot으로 분석하였다. 그 결과 Fig. 2와 Fig. 3에서 보는 바와 같이 LPS-PG에 의해 유 도된 p65의 인산화가 PYEE에 의해 농도 의존적으로 감 소하는 것을 확인할 수 있었고, 상위신호전달물질 중에 서는 JNK가 PYEE에 의해 인산화가 억제되는 것을 볼 수 있었다.
Fig. 2. PYEE inhibited phosphorylation of NF-κB in LPS-PG stimulated HGF-1 cells. Panel A shows phosphorylated status of p65 by PYEE. All signals were normalized to protein levels of actin, an internal control, and expressed as a ratio (Panel B). Data represent the mean±SD of triplicate experiments. Values sharing the same superscript are not significantly different at p<0.05 by Duncan’s multiple range test.
Fig. 3. PYEE inhibited phosphorylation of JNK in LPS-PG stimulated HGF-1 cells. Panel A shows phosphorylation levels of Akt, ERK, JNK and p38 by PYEE. All signals were normalized to protein levels of actin, an internal control, and expressed as a ratio (Panel B). Data represent the mean±SD of triplicate experiments. Values sharing the same superscript are not significantly different at p<0.05 by Duncan’s multiple range test.
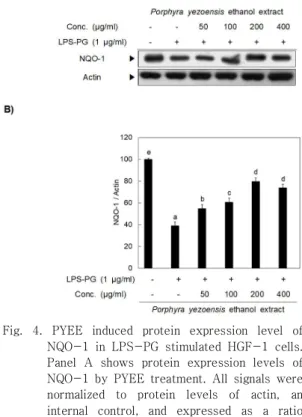
3. LPS-PG로 유도된 HGF-1 cell의 염증 에서NQO-1 발현 유도를 통한 PYEE 의 항염 효과
제독효소인 2상 효소(phase II enzyme)의 발현 유도에 의해 염증이 억제된다는 사실이 보고됨에 따라 본 실험 에서도 2상 효소 중 하나인 NQO-1의 발현을 분석하였다 [20]. Fig. 4에서 보는 것과 같이 LPS-PG에 의해 일시적 으로 감소했던 NQO-1은 PYEE의 처리에 의하여 농도 의존적으로 다시 증가하는 것을 확인할 수 있었다. 본 연 구의 결과 PYEE는 LPS-PG에 의해 유도된 염증을 MAPK 중 하나인 JNK와 전사인자인 NF-κB의 인산화 억제를 통해 조절하고, 2상 효소중 하나인 NQO-1의 발 현 유도를 통해 조절하는 것으로 밝혀졌다.
Fig. 4. PYEE induced protein expression level of NQO-1 in LPS-PG stimulated HGF-1 cells.
Panel A shows protein expression levels of NQO-1 by PYEE treatment. All signals were normalized to protein levels of actin, an internal control, and expressed as a ratio (Panel B). Data represent the mean±SD of triplicate experiments. Values sharing the same superscript are not significantly different at p<0.05 by Duncan’s multiple range test.
4. 논의
LPS와 같은 염증 유발물질로부터의 자극에 의해 염증 이 발생하면 대식세포에서는 NO, PGE2, 염증성 cytokine 인 TNF-α, IL-6, IL-1β 등이 분비된다. 염증반응에서 지 표물질로 흔히 사용되는 NO는 NOS에 의해 L-arginine 으로부터 합성되고, NOS에는 endothelial NOS (eNOS), neuronal NOS (nNOS), iNOS의 세가지가 존재한다. 이 중 iNOS의 과발현에 의한 NO 과다 생성과 방출로 인하 여 염증반응을 촉진하는 역할을 하며, 조직손상과 신경 손상은 물론 유전자변이를 일으키기도 한다[21,22]. 또 다 른 염증의 지표인 COX-2는 arachidonic acid로부터 prostaglandin의 생성을 촉진하는 효소로 그 중 PGE2는 만성 염증 조직과 악성 종양 조직에서 과 발현이 관찰되 기도 한다[23]. Daghigh F. 등의 연구 결과에 따르면 TNF-α, IL-1β, interferon (IFN)-γ와 같은 염증성 사이 토카인의 처리에 의해 HGF cell에서 NO의 발현이 증가 되었고, 치주염 유발 동물모델에서도 유의한 NO의 증가 가 관찰되었다[24]. COX-2 또한 염증이 발생된 동물의 치주조직에서 과 발현되었고 이의 조절을 통해 치주염을 억제할 수 있다는 사실이 보고되었다[25]. 따라서 치주를 구성하는 주요한 세포중 하나인 HGF cell에 LPS-PG로 유도된 iNOS와 COX-2의 발현을 유의적으로 억제할 수 있는 물질은 치주염을 완화할 수 있는 활성을 가진 것으 로 생각할 수 있고 본 연구에서 사용한 PYEE는 이 두 가 지 염증 지표물질의 유전자 발현을 농도가 높아질수록 화과가 커 농도 의존적 효과를 보였다.
NF-κB는 세포분화, 염증반응, 세포 부착, 면역 반응 등과 관련된 다양한 유전자의 발현에 관여하는 전사인자 의 역할을 하는 것으로 알려져 있으며, 종양형성, 자가 면 역질환, 염증 질환 유발에도 관여를 하고 있다. NF-κB는 세포질 내에서 p50과 p65가 결합된 heterodimer에 IκB가 부착된 형태의 불활성형으로 존재하다가 reactive oxygen species (ROS), LPS, 염증성 cytokine과 같은 자극으로 I κB kinase에 의해 IκB가 인산화되면서 proteasome으로 이동 후 phospho-IκB가 분해된다. IκB가 유리되면서 p50 과 p65의 heterodimer 또한 활성화되고 이 때 p65가 인산 화 된다. 그리고 활성화된 NF-κB는 핵내로 translocation한 후 염증매개물질인 iNOS, COX-2, 염증성 cytokine의 전 사를 위한 promoter region에 결합하여 유전자 발현을 촉진 시킨다[3]. 따라서 NF-κB 활성 조절을 통해 염증
매개 물질을 억제할 수 있게 되면 치주염을 포함해 다양 한 염증 관련 질환 치료에 효과적일 수 있을 것으로 생각 한다. 본 실험에서는 LPS-PG로 활성화된 p65의 인산화 가 PYEE의 처리에 의해서 농도 의존적으로 억제된 것으 로 보아 PYEE는 NF-κB 활성을 조절함으로써 HGF-1 cell의 염증을 억제하는 것으로 생각된다. 이는 Kim 등 [19]의 연구에서 C. xanthorrhiza 추출물의 HGF-1 cell의 염증 억제효과가 있는 것으로 나타난 것과 유사한 결과 를 보였으나 염증 유발에 있어 본 연구는 LPS-PG를 이 용하였고, Kim 등은 LPS를 이용하여 차이를 보였다.
MAPKs는 세포 증식, 분화, 생존과 사멸과 같은 다양 한 세포 반응의 조절과 관련된 serine/threonine kinase로 써 염증 매개 인자들의 발현을 조절하는 NF-κB, activator protein (AP)-1등과 같은 전사인자의 활성화에 영향을 주는 가장 대표적인 신호전달물질이다. 현재까지 포유류의 세포에서는 ERK, JNK, p38 등 최소 세 종류의 MAPKs 신호전달경로가 보고되어 있다[3]. 여러 선행 연 구를 통해 ERK는 세포 외부의 다양한 자극 인자에 대해 반응하고 전사인자를 활성화할 수 있는 반면, JNK와 p38 은 LPS나 TNF-α와 같은 염증성 사이토카인이나 세포 스트레스 유발 물질에 의해 유도된 스트레스에 의해 활 성화되는 것으로 알려져 있다[26-28]. Akt 또한 세포 증 식과 성장, 그리고 세포주기 조절 등 다양한 세포 반응의 조절과 관련되어 있는 것으로 알려져 있고 PI3K에 의해 활성화되면서 외부 신호를 생체내로 전달한다[3]. 본 연 구에서는 PYEE가 HGF-1 cell에서 LPS-PG에 의해 유 도되는 염증 반응에서 MAPKs나 PI3K/Akt 활성도에 미 치는 영향을 분석하기 위하여 Akt, ERK, JNK, p38의 인 산화 정도를 Western blot analysis를 이용하여 분석하였 다. 그 결과 PYEE는 Akt, ERK, p38의 인산화에는 영향 을 미치지 못하였으나 JNK의 활성은 효과적으로 억제하 는 것을 확인할 수 있었다. 이 결과로 PYEE는 JNK의 활 성을 억제함으로써 항염증 작용을 나타내는 것으로 생각 된다.
한편, 세포 내 호흡 과정 중 산소의 불완전한 환원으로 인하여 발생하는 ROS는 세포에 손상을 주는 여러 인자 중 하나이고, 세포는 superoxide (SOD), heme oxygenase (HO)-1, NQO-1과 같은 항산화 효소를 활성화시킴으로 써 이들로부터 자신을 방어한다. 그러나 LPS에 노출되면 과량의 ROS가 생산되고, 이에 따라 세포내 산화적 스트 레스가 축적되면 여러 종류의 질환이 유발되기도 한다
[29]. 세포질에 존재하는 NQO-1은 quinone 화합물을 환 원시키고 이 과정에서 활성산소에 의한 세포 독성을 감 소시키기도 하는 것으로 알려져 있다. Yang 등[30]의 연 구에서는 Nrf2에 의한 NQO-1과 HO-1의 활성이 염증 인자인 iNOS와 COX-2의 발현 억제와 관련 있음을 보여 주었다. 본 실험에서도 2상 효소인 NQO-1이 LPS-PG가 처리된 HGF-1 cell에서 어떻게 발현되는지를 알아보았 고, 그 결과 LPS-PG의 처리에 의해 억제된 NQO-1의 발 현이 PYEE의 처리에 의해 다시 증가되는 것을 확인할 수 있었으며 이는 Yang등의 연구에서 작용한 것과 같은 기전이 HGF-1 cell에서도 일어날 수 있음을 보여준 것으 로 생각할 수 있다[30].
결론적으로 PYEE는 HGF-1 cell에서 LPS-PG에 의 해 유도된 염증에서 NO와 PGE2의 활성을 매개하는 유 전자인 iNOS와 COX-2를 농도의존적으로 억제하였고, 이는 전사인자인 NF-κB와 JNK의 활성 억제를 통한 것 임을 알 수 있었을 뿐만 아니라 2상 효소인 NQO-1의 활 성을 유도함으로써 더 강한 항염 활성을 보였다. 이를 통 해 PYEE는 치주질환의 발생을 억제하며, 예방을 위한 기능성소재로서의 가치를 발견할 수 있었다.
5. 결론
인간 치은섬유모세포는 치주조직에 존재하는 주요한 세포의 한 형태로써 염증성 자극에 반응하여 여러 염증 매개 물질을 분비한다. 본 연구에서는 치주염을 일으키 는 주요한 원인균 중 하나인 P. gingivalis로부터 분리한 LPS를 이용하여 인간 치은섬유모세포인 HGF-1 cell에 염증을 유도한 후 PYEE의 항염효과를 분석하였다. 실험 결과 LPS-PG에 의해 유도된 iNOS와 COX-2의 단백질 발현은 PYEE의 처리에 의해 농도 의존적으로 억제되었 고, 전사인자인 NF-κB 또한 동일하게 발현이 줄어들었 다. 그리고 이들의 발현을 조절하는 상위신호전달물질인 MAPKs와 PI3K/Akt의 인산화를 분석한 결과 PYEE의 처리에 의해 JNK의 활성은 억제되었으나 Akt, ERK, p38 에는 영향을 미치지 않았음을 알 수 있었다. 또한 과발현 으로 항염증효과를 나타내는 것으로 알려진 제2상 효소 중 하나인 NQO-1을 분석한 결과 LPS-PG에 의해 줄어 들고 PYEE의 처리로 다시 발현이 증가되었다. 결론적으 로 PYEE는 NF-κB와 JNK의 활성 억제와 NQO-1 활성
유도를 통해 HGF-1 cell에서 LPS-PG에 의해 유도된 염 증을 조절하는 것으로 사료되고, 본 논문을 통해 PYEE 는 치주질환 억제에 효과적인 항염 후보물질로 이용될 수 있을 것으로 생각한다.
REFERENCES
[1] T. Lawrence, D.A. Willoughby & D.W. Gilroy. (2002).
Anti-inflammatory lipid mediators and insights into the resolution of inflammation. Nat Rev Immunol, 2(10), 787-795.
DOI : 10.1038/nri915
[2] J.K. Kundu & Y.J. Surh. (2008). Inflammation: gearing the journey to cancer. Mutat Res, 659(1-2), 15-30.
DOI : 10.1016/j.mrrev.2008.03.002
[3] Y.J. Surh. (2003). Cancer chemoprevention with dietary phytochemicals. Nat Rev Cancer, 3(10), 768-780.
DOI : 10.1038/nrc1189
[4] G.C. Armitage. (1999). Development of a classification system for periodontal diseases and conditions. Ann Periodontol, 4(1), 1-6.
DOI : 10.1902/annals.1999.4.1.1
[5] G. Hajishengallis & R.J. Lamont. (2014). Breaking bad:
manipulation of the host response by porphyromonas gingivalis. Eur J Immunol, 44(2), 328–338.
DOI : 10.1002/eji.201344202
[6] P.K. Sreenivasan & A. Gaffar. (2008). Antibacterials as anti-inflammatory agents. Dual action agents for oral health. Antonie Van Leeuwenhoek, 93(3), 227-239.
DOI : 10.1007/s10482-007-9197-8
[7] D. Noda, T. Hamachi, K. Inoue & K. Maeda. (2007).
Relationship between the presence of periodontopathic bacteria and the expressing of chemokine receptor mRNA in inflamed gingival tissue. J Periodontal Res, 42(6), 566-571.
DOI : 10.1111/j.1600-0765.2007.00984.x
[8] D.T. Graves, M. Oskoui & S. Volejnikova. (2001).
Tumor necrosis factor modulates fibroblast apoptosis, PMN recruitment, and osteoclast formation in response to P. gingivalis infection. J Dent Res, 80(10), 1875-1879.
DOI : doi.org/10.1177/00220345010800100301
[9] L. Kocgozlu, R. Elkaim, H. Tenenbaum & S. Werner.
(2009). Variable cell responses to P. gingivalis lipopolysaccharide. J Dent Res, 88(8), 741-745.
DOI : 10.1177/0022034509341166
[10] Y.K. Han, I.S. Lee & S.I. Lee. (2017). JAK/STAT
pathway modulates on Porphyromonas gingivalis lipopolysaccharide- and nicotine-induced inflammation in osteoblasts. J Dent Hyg Sci, 17(1), 81-86.
DOI : 10.17135/jdhs.2017.17.1.81
[11] G.J. Seymour & E. Gemmell. (2001). Cytokines in periodontal disease: where to from here?. Acta Odontologica, 59(3), 167-173.
DOI : 10.1080/000163501750266765
[12] L. Li, W. Sun, T. Wu, R. Lu & B. Shi. (2017). Caffeic acid phenethyl ester attenuates lipopolysaccharide- stimulated proinflammatory responses in human gingival fibroblasts via NF-κB and PI3K/Akt signaling pathway.
Eur J Pharmacol, 794(5), 61-68.
DOI : 10.1016/j.ejphar.2016.11.003.
[13] D. Ross, J.K. Kepa, S.L. Winski, H.D. Beall, A. Anwar
& D. Siegel. (2000). NAD(P)H: quinone oxidoreductase 1 (NQO1): chemoprotection, bioactivation, gene regulation and genetic polymorphisms. Chem Biol Interact, 129(1-2), 77-97.
DOI : 10.1016/S0009-2797(00)00199-X
[14] M.N. García-Casal, J. Ramírez, I. Leets, A.C. Pereira &
M.F. Quiroga. (2009). Antioxidant capacity, polyphenol content and iron bioavailability from algae (Ulva sp., Sargassum sp. and Porphyra sp.) in human subjects.
Brit J Nut, 101(1), 79-85.
DOI : 10.1017/s0007114508994757
[15] H. Funahashi, T. Imai, T. Mase, M. Sekiya, K. Yokoi, H.
Hayashi, A. Shibata, T. Hayashi, M. Nishikawa, N. Suda, T. Hibi, Y. Mizuno, K. Tsukamura, A. Hayakawa & S.
Tanuma. (2001). Seaweed prevents breast cancer?. Jpn J Cancer Res, 92(5), 483–487.
DOI : 10.1111/j.1349-7006.2001.tb01119.x
[16] S.J. Oh, J.I. Kim, H.S. Kim, S,J. Son & E.O. Choe. (2013).
Composition and antioxidant activity of dried laver.
dolgim. Kor J Food Sci Technol, 45(4), 403-408, 2013.
DOI : 10.9721/kjfst.2013.45.4.403
[17] E.S. Shin, H.J. Hwang, I.H. Kim & T.J. Nam. (2011) A glycoprotein from Porphyra yezoensis produces anti-inflammatory effects in liposaccharide-stimulated macrophages via the TLR4 signaling pathway. Int J Mol Med. 28(5), 809-815.
DOI : 10.3892/ijmm.2011.729
[18] H.A. Lee, I.H. Kim & T.J. Nam. (2015) Bioactive peptide from Pyropia yezoensis and its anti-inflammatory activities. Int J Mol Med. 36(6), 1701-1706.
DOI : 10.3892/ijmm.2015.2386
[19] S.K. Kim, K.E. Kook, C.H. Kim, J.K. Hwang. (2018).
Inhibitory Effects of Curcuma xanthorrhiza Supercritical
Extract and Xanthorrhizol on LPS-Induced Inflammation
in HGF-1 Cells and RANKL-Induced
Osteoclastogenesis in RAW264.7 Cells. J Microbiol.
Biotechnol, 28(8) 1270-1281.
DOI : 10.4014/jmb.1803.03045
[20] H.L. Yang, S.W. Lin, C.C. Lee, K.Y. Lin, C.H. Liao, T.Y.
Yang, H.M. Wang, H.C. Huang, C.R. Wu & Y.C. Hseu.
(2015). Induction of Nrf2-mediated genes by Antrodia salmonea inhibits ROS generation and inflammatory effects in lipopolysaccharide-stimulated RAW264.7 macrophages. Food Funct, 6(1) 230-241.
DOI : 10.1039/c4fo00869c
[21] C. Nathan. (1992). Nitric oxide as a secretory product of mammalian cells. FASEB J, 6(12), 3051-3064.
DOI : 10.1096/fasebj.6.12.1381691
[22] H.N. Lee, D.Y. Lim, S.S. Lim, J.D. Kim & J.H. Yoon.
(2011). Anti-inflammatory effect of ethanol extract from Eupatorium japonicum. Kor Soc Food Sci Technol, 43(1), 65-71.
DOI : 10.9721/kjfst.2011.43.1.065
[23] H.K. Na & Y.J. Surh. (2006). Intracellular signaling network as a prime chemopreventive target of (-)-epigallocatechin gallate. Mol Nutr Food Res, 50(2), 152-159.
DOI : 10.1002/mnfr.200500154
[24] F. Daghigh, R.C. Borghaei, R.D. Thornton & J.H. Bee.
(2002). Human gingival fibroblasts produce nitric oxide in response to proinflammatory cytokines. J Periodontol, 73(4), 392-400.
DOI : 10.1902/jop.2002.73.4.392
[25] M. Holzhausen, C.J. Rossa, E.J. Marcantonio, P.O.
Nassar, D.M. Spolidorio & L.C. Spolidorio. (2002). Effect of selective cyclooxygenase-2 inhibition on the development of ligature-induced periodontitis in rats. J Periodontol, 73(9), 1030-1036.
DOI : 10.1902/jop.2002.73.9.1030
[26] M.J. Robinson & M.H. Cobb. (1997). Mitogen-activated protein kinase pathways. Curr Opin Cell Biol, 9(2), 180-186.
DOI : 10.1016/S0955-0674(97)80061-0
[27] U. Hidding, K. Mielke, V. Waetzig, S. Brecht, U.
Hanisch, A. Behrens, E. Wagner & T. Herdegen. (2002).
The c-Jun N-terminal kinases in cerebral microglia:
immunological functions in the brain. Biochem Pharmacol, 64(5-6), 781-788.
DOI : 10.1016/s0006-2952(02)01139-5
[28] V. Waetzig, K. Czeloth, U. Hidding, K. Mielke, M.
Kanzow, S. Brecht, M. Goetz, R. Lucius, T. Herdegen &
U.K. Hanisch. (2005). c-Jun N-terminal kinases (JNKs) mediate pro-inflammatory actions of microglia. Glia, 50(3), 235-246.
DOI : 10.1002/glia.20173
[29] K.G. Lee, HI. Lee & H.S. Jeong. (2015).
Cheogjogupye-tang has anti-oxidant potential through the activation of Nrf2. J Physiol & Pathol Korean Med, 29(2), 174-179.
DOI : 10.15188/kjopp.2015.04.29.2.174
[30] C. Yang, C. Zhang, Z. Wang, Z. Tang, H. Kuang & A.N.
Kong. (2016). Corynoline isolated from corydalis bungeana turcz. exhibits anti-inflammatory effects via modulation of Nfr2 and MAPKs. Molecules, 27(8), 1-15.
DOI : 10.3390/molecules21080975
박 충 무(Chung Mu Park) [정회원]
․2005년 2월 : 인제대학교 임상병리 학 전공 (이학석사)
․2011년 2월 : 인제대학교 식의약생 명공학 전공 (이학박사)
․인제대학교 의과대학 병리학교실 연구강사
․2013년 3월 ∼ 현재 : 동의대학교 임상병리학과 조교수
․관심분야 : 천연물의 항산화·항염증 기전
․E-Mail : [email protected]
윤 현 서(Hyun-Seo Yoon) [정회원]
․2008년 8월 : 인제대학교 보건대 학원 (보건학석사/병원경영)
․2011년 2월 : 인제대학교 대학원 (보건학박사)
․2013년 3월 ∼ 2018년 2월 : 동의 대학교 치위생학과 조교수
․2018년 3월 ∼ 현재 : 동의대학교 치위생학과 부교수
․관심분야 : 임상치과학, 보건학, 예방의학
․E-Mail : [email protected]