Escherichia coli의 지방산 대사과정 및 배양조건 조절을 통한 FAEE 생산
문관우1, 임승락1, 박혜민1, 윤현식1, 김세경2*, 백종윤1*
FAEE Production in Escherichia coli by Control of Fatty Acid Metabolism and Cultivation Condition
Kwan Woo Moon1, Seung Rak Lim1, Hye Min Park1, Hyun Shik Yun1, Se Kyung Kim2*, and Jong Youn Baik1*
Received: 25 June 2020 / Revised: 9 August 2020 / Accepted: 10 August 2020
© 2020 The Korean Society for Biotechnology and Bioengineering
Abstract: Global warming is a major issue of modern cli- mate change, and caused by the greenhouse effect mainly due to carbon dioxide emitted by burning fossil fuels. To reduce the emission of carbon dioxide, scientists have focused on replacing fossil fuels with renewable energy. Biofuel, one form of renewable energy derived from the carbon sources of natural biomass, can contribute to reduced carbon dioxide emission. In this study, Escherichia coli was engineered for the production of fatty acid ethyl ester (FAEE), biodiesel derived from fatty acids. For production of FAEE, pyruvate decarboxylase (pdc), alcohol dehydrogenase II (adhB), acyl- Coenzyme A : diacylglycerol acyltransferase (atfA), and fatty acid synthesis (FAS) pathway genes were selected and cloned on to various plasmids. We designed and introduced various combinations of expression constructs in E. coli to enhance FAEE production. As a result, a 3.88-fold increase in FAEE production was achieved by modifying the expression levels of the genes involved in the initiation step and the β-oxida- tion step. The production of FAEE at different growth phases as well as the effect of the culture conditions such as tempera- ture and carbon sources on FAEE productivity were studied.
As a future study, scale-up studies through fermenter cultiva- tion would contribute to mass production of biodiesel.
Key words: biodiesel, fatty acid ethyl ester, Escherichia coli, initiation step, β-oxidation step
1. INTRODUCTION
온실효과는 현대 기후 변화 중에서 가장 큰 문제점으로 인식 되는 지구 온난화의 원인 중 하나로, 지구로 들어온 태양의 복사열이 지구밖으로 다시 나가지 못하고 순환하는 현상을 의미한다 [1]. 온실효과를 유발하는 기체를 온실가스라고 하 는데, 그 예로 화석연료가 연소하며 생기는 이산화탄소, 메 탄, 아산화질소 등이 있다 [2]. 이 중 가장 큰 원인이 되는 이 산화탄소는 전 세계적으로 화석연료 사용량에 비례하여 배 출되는 양이 증가하고 있고 [3], 이로 인해 지구 온난화를 유 발하여, 해수온도와 육지온도의 증가로 극지방의 빙하가 녹 아 해수면이 상승하는 등, 생태계의 파괴가 일어나고 있다 [4]. 따라서 이산화탄소 배출량을 줄이기 위해 화석연료를 대체할 수 있는 재생가능한 에너지의 개발 및 연구가 진행되 어왔다 [5,6]. 바이오 디젤은 동·식물 유래의 긴 사슬 지방산 에스테르로 [7], 화석연료 유래의 디젤유를 대체하거나, 디 젤유에 첨가하여 사용이 가능하기 때문에 현존하는 디젤 기 관을 변형없이 바로 사용할 수 있다는 장점이 있다 [8]. 현재 상업적으로 가장 많이 사용되는 바이오 디젤은 지방산 메틸 에스테르 (FAME)로, 동·식물 유래의 트리글리세라이드와 화석연료 유래 메탄올의 에스테르화 반응에 의해 생성된다.
하지만 이 과정에서 화학반응을 위한 고열이 필요하며 부산 물로 글리세롤이 다량 생산된다는 단점이 있다 [9]. 이 문제 를 해결하기 위해 미생물 대사과정을 조절하여 바이오 디젤 중 하나인 지방산 에틸 에스테르 (FAEE)를 생합성하기 위한 연구들이 다양하게 진행되었다 [10-12]. 미생물에서 생합성 되는 FAEE는 에탄올과 acyl-CoA를 전구체로 하여 효소에
1인하대학교 생명공학과
1Department of Biological Engineering, Inha University, Incheon 22212, Korea
Tel: +82-32-860-7513; Fax: +82-32-872-4046, E-mail: [email protected]
2이화여자대학교 화학신소재공학과
2Division of Chemical Engineering and Materials Science, Ewha Womans University, Seoul 03760, Korea
Tel: +82-2-3277-4756, Fax: +82-2-3277-3535 E-mail: [email protected]
Research Paper
의해 생합성 되므로 부산물이 생기지 않아 친 환경적이라는 장점이 있다. 미생물 유래 FAEE 생산량을 늘리기 위한 연구 들은 대사과정 조절을 통해 FAEE 의 전구체인 에탄올의 생산 량과 [13], 지방산의 생산량을 증가시킨 연구들이 있다 [14].
FAEE 생합성은 2006년 Rainer. K. 연구진이 에탄올 생산량 증 가를 위해 Zymomonas mobilis에서 pyruvate를 acetaldehyde로 전환하는 pyruvate decarboxylase (pdc)와 전환된 acetaldehyde 를 ethanol로 전환하는 alcohol dehydrogenase II (adhB)를 대 장균에 도입하고, 에탄올과 지방산을 반응하여 FAEE를 생합 성 하기 위해 Acinetobacter baylyi에서 유래한 acyltransferase ( atfA) 유전자를 도입한 연구를 진행하였다[15]. 그 후, Steen, E.J. 연구진에 따르면 thioesterase I (tesA), acyl-CoA ligase ( fadD) 유전자 과발현과 acyl-CoA dehy-drogenase (fadE) 유 전자 제거를 통한 FAEE 생산량 증가 등 대장균의 지방산 대 사회로 조절을 통해 FAEE 생산량을 향상시킨 연구들이 진 행 되었다 [16].
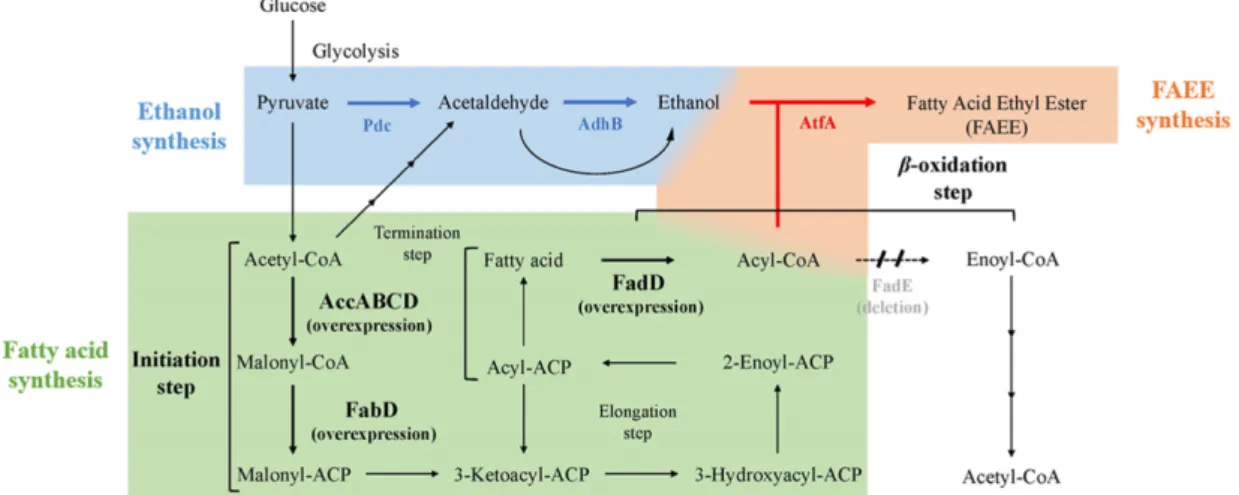
본 연구에서는 Escherichia coli에 pdc, adhB 두 유전자를 도입하여 FAEE의 전구체인 에탄올 생산량을 향상시킨 균주 를 제작하였고, atfA 유전자를 추가로 도입하여 제작한 FAEE 생산 균주에 배양온도와 글루코스의 첨가량 등을 조 절해 적합한 배양조건을 확인하였다. 그리고 지방산 대사회 로의 개시단계 유전자 acetyl-CoA carboxylases (accABCD), malonyl-CoA:ACP transacylase ( fabD)의 과발현과 베타옥시 데이션 단계 유전자 fadD의 과발현, fadE 제거를 통해 지방 산 생산량을 향상시켜 FAEE 생산량 증가를 목적으로 하였 다 (Fig. 1).
2. MATERIALS AND METHODS
2.1. 사용 균주 및 플라스미드
재조합 플라스미드의 제조와 유지에는 E. coli XL1-Blue를 사용하였고, FAEE 생산을 위한 균주는 외래 단백질을 효과 적으로 발현할 수 있는 E. coli BL21(DE3) (Stratagene, San
Diego, California, USA)을 사용하였다. 지방산 대사회로의 개 시단계 (accA, accB, accC, accD, fabD)와 베타 옥시데이션 단 계 (fadD)의 유전자의 genomic DNA는 E. coli K-12 MG1655로 부터 도입하였다 [16-18]. 에탄올의 생산량 향상을 위해 도입 한 pdc, adhB 유전자는 Zymomonas mobilis ZM4 (ATCC 31821) 균주에서, 그리고 에탄올과 지방산을 FAEE로 합성하게 하는 acyltransferase들 중 하나인 atfA 유전자는 Acinetobacter baylyi ADP1 (ATCC 33305) 균주에서 획득하였다. 유전자의 발현 을 위한 클로닝 벡터는 pCDFDuet
TM-1 (Merck, Damstadt, Germany)과 pCOLADuet
TM-1 (Merck, Damstadt, Germany)를 사용하였다. 본 연구에서 사용된 모든 FAEE 생산 균주는 지 방산 생산량의 향상을 위하여 pKOV 벡터를 이용한 방법으 로 베타 옥시데이션 단계의 fadE 유전자를 제거하였다 [19].
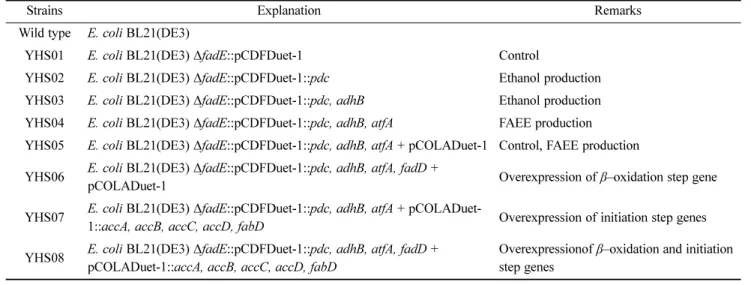
모든 DNA의 클로닝은 표준 과정을 따라 진행되었으며 사용 된 프라이머는 코스모진텍(Cosmo Genetech, Inc., Seoul, Korea)에서 제작하였다 [20]. Table 1은 본 연구에서 사용된 균주들의 목록이다.
2.2. 배양조건
플라스미드를 포함하는 E. coli BL21 (DE3)는 LB 배지 (Tryptone 10 g/L, NaCl 5 g/L, Yeast extract 5 g/L)에 10 g/L의 글루코스를 첨가하여 배양하였다. Selection marker의 경우 streptomycin은 50 mg/L, kanamycin은 60 mg/L 농도로 첨가 하였다. 500 mL flask에 200 mL의 배지를 넣어서 실험하였으 며 37
oC, 180 rpm에서 배양을 시작하여 OD(
600 nm)가 0.8~1.0 일 때 Isopropyl- 1-thio-β-D-galactopyranoside (IPTG)를 1 mM의 농 도가 되도록 첨가하여 단백질 발현을 유도하였다. 배양온도 실험의 경우 발현 유도와 동시에 온도를 변화시켰다.
2.3. HPLC 분석
글루코스의 소모와 에탄올의 생산을 확인하기위해 Liquid chromatography (YoungLin 9100 HPLC system, Young Lin instrument Co., Anyang, Korea) 장비를 사용하여 분석하였다.
Column은 BioRad Aminex HPX-87H (Bio-Rad, Hercules
Fig. 1. Strategy of FAEE production by using ethanol and fatty acid synthesis pathway in E. coli.
California, USA)를 사용하였고, 이동상으로는 5 mM의 H
2SO
4를 사용하였다. 이동상은 0.6 mL/min의 속도로 흘려주었고 column 온도는 50
oC, RI detector 의 온도는 35
oC 로 하였다.
2.4. 지방산과 FAEE 추출 및 분석
지방산의 분석은 배양액 90 mL을 원심분리 (7,000 rpm, 10 min) 하여 동결건조 한 dry cell에 methanol:sulfuric acid (100:5, v/
v) 용액을 3 mL넣고 질소 충전 후 90
oC oven 에서 30분동안 methylation시켰다 [21]. 반응 후 hexane 700 μL와 물을 첨가 하여 orbital shaker에서 30분동안 교반하였다. 원심분리 (7,000 rpm, 5 min) 하여 hexane층인 위층을 분리하여 필터링 한 후 Gas chromatography (Agilent HP 6890 series, Agilent, Santa Clara, California, USA) 의 FID (flame ionized detector) 를 사용하여 분석하였다. Column은 Agilent J&W HP-5ms capillary column (30 × 0.25 × 0.25, length(m) × I.D.(mm) × film(μm)) (Agilent, Santa Clara, California, USA) 를 사용하였다.
FAEE의 분석을 위해 배양액 5 mL 당 chloroform:methanol (2 : 1, v/v) 용액을 6 mL 첨가하여 bead-beater (FastPrep-24
TM5G, MP Biomedicals, Irvine, CA, USA)를 사용하여 cell을 파 쇄하였다. 그 후 원심분리 (7,000 rpm, 5 min)하여 아래층 (chloroform) 을 분리한 후 질소를 사용하여 chloroform을 증 발시켰다. 여기에 500 μL의 hexane을 첨가하여 필터링 한 후 지방산과 같은 방법으로 분석하였다 [22,23].
3. RESULTS AND DISCUSSION
3.1. 이종 유전자 도입을 통한 대장균에서의 에탄올 생산량 증가
Control 균주는 벡터만 포함된 균주 YHS01을 사용하였고, pdc 유전자와 pdc, adhB 유전자를 실험군으로 선정하였다.
adhB 유전자만을 도입한 균주는 에탄올 생산량의 변화가 거 의 없었던 선행 연구 결과를 참고하여 포함하지 않았다 [24].
실험 균주 별 에탄올 생산량 (Fig. 2)을 비교해 보면 균주 YHS01에선 배양시작 후 23시간까지 에탄올 농도가 증가하 였고 최대 에탄올 농도는 0.34 g/L이었다. pdc 유전자만을 도 입한 균주 YHS02에선 배양시작 후 28시간에 최대 에탄올 농 도 2.66 g/L를 생산하였다. pdc, adhB 두 가지유전자를 도입 한 균주 YHS03에선 배양시작 후 에탄올 농도가 증가하여 13 시간에 최대 농도 (4.33 g/L)의 에탄올을 생산한 후 감소하였 다. 시간에 따라 에탄올 농도의 변화를 보면 실험에 사용한 3 가지 균주에서 에탄올 농도가 최대 농도 도달 후 점차 감소 하는 모습을 보였는데, 이것은 배지 내 글루코스가 다 소모 되었기 때문에 에탄올 생성 대사과정이 더 이상 진행되지 않 았기 때문이라고 생각된다 (data not shown). 에탄올 생산량 증대를 목적으로 E. coli에 pdc와 adhB를 도입한 다른 연구들 과 비교해 보면 pfl 유전자 위치에 pdc와 adhB를 염색체 통합 Table 1. List of recombinant plasmids and strains
Strains Explanation Remarks
Wild type E. coli BL21(DE3)
YHS01 E. coli BL21(DE3) ΔfadE::pCDFDuet-1 Control
YHS02 E. coli BL21(DE3) ΔfadE::pCDFDuet-1::pdc Ethanol production
YHS03 E. coli BL21(DE3) ΔfadE::pCDFDuet-1::pdc, adhB Ethanol production YHS04 E. coli BL21(DE3) ΔfadE::pCDFDuet-1::pdc, adhB, atfA FAEE production YHS05 E. coli BL21(DE3) ΔfadE::pCDFDuet-1::pdc, adhB, atfA + pCOLADuet-1 Control, FAEE production YHS06 E. coli BL21(DE3) ΔfadE::pCDFDuet-1::pdc, adhB, atfA, fadD +
pCOLADuet-1 Overexpression of β–oxidation step gene
YHS07 E. coli BL21(DE3) ΔfadE::pCDFDuet-1::pdc, adhB, atfA + pCOLADuet-
1::accA, accB, accC, accD, fabD Overexpression of initiation step genes
YHS08 E. coli BL21(DE3) ΔfadE::pCDFDuet-1::pdc, adhB, atfA, fadD + pCOLADuet-1::accA, accB, accC, accD, fabD
Overexpressionof β–oxidation and initiation step genes
Fig. 2. Ethanol production in recombinant strains. The black bar
is the amount of ethanol was produced by strain YHS01, the light
gray bar is the amount of ethanol was produced by strain YHS02,
the gray bar is the amount of ethanol was produced by strain
YHS03.
(chromosome integration)의 방법으로 삽입하여 4.0 g/L의 에 탄올을 생산하였다 [25]. 최근 코돈 최적화를 통한 퓨전 단백 질을 이용한 연구 [26]에서는 코돈 최적화 과정을 거친 균주 에서 24시간 동안 약 5.62 g/L의 에탄올을 생산하였으나 최 적화를 하지 않은 컨트롤 균주는 약 1.88 g/L의 생산량을 나 타낸 것으로 보아 부산물 대사회로 제거나 단백질 개발 등을 하지 않고 pdc, adhB 도입만으로 수행한 에탄올 생산량은 적 당한 수준이라고 생각한다. 균주 YHS02는 균주 YHS01에 비해 에탄올 생산량이 약 1.8배 증가하였고, 균주 YHS03의 경우 control 균주 YHS01 대비 약 10.8배 증가하였다. 이는 pyruvate에서 acetaldehyde로 전환되는 과정에서 기존 YHS01 균주는 acetyl-CoA, acetate, acetaldehyde의 여러 대사 단계를 거치는 반면 pdc 유전자를 포함한 YHS01과 YHS03 균주는 pyruvate에서 바로 acetaldehyde로 대사과정이 진행되기 때 문이라고 생각한다. 이후 진행한 실험은 글루코스를 포함하 며 pdc, adhB 두 가지 유전자를 포함시킨 균주를 사용하였다.
3.2. FAEE 생산에 관한 배양조건 최적화
에탄올 생산량이 증가된 것을 확인한 균주 YHS03에 FAEE 생산에 필수적인 atfA유전자를 도입하여 FAEE 생산 균주인 균주 YHS04를 제작하였다. LB 배지에 글루코스 농도를 10 g/L 와 배양온도를 37
oC 로 하여 균주 YHS04를 배양한 결 과 약 0.068 g/L의 FAEE가 생산되었고, FAEE의 구성은
saturated FAEE (sFAEE)인 ethyl laurate (C12:0) 0.11%, ethyl myristate (C14:0) 8.85%, ethyl palmitate (C16:0) 44.10%, ethyl stearate (C18:0) 5.3%와 unsaturated FAEE (usFAEE) ethyl palmitoleate (C16:1) 18.32%, ethyl oleate (C18:1) 23.32% 로 나타났다. 배양온도와 공급된 글루코스 농도에 따른 FAEE 생산량을 알아보기 위해 200 ml의 배지를 포함하는 500 ml flask에서 글루코스 농도를 10 g/L와 20 g/L, 그리고 배양온 도 30
oC와 37
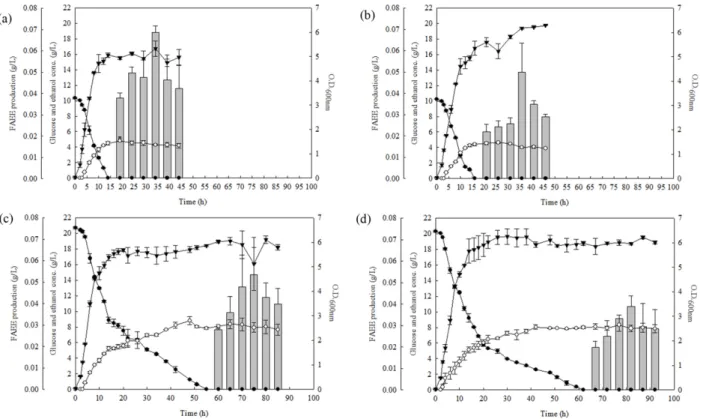
oC로 각각 다르게 하여 배양하였다. 실험은 같 은 조건에서 2회 반복하여 진행하였고, 배양시작 후 2시간마 다 샘플링하여 OD, 글루코스 잔량과 에탄올 생산량을 측정 하였고 FAEE는 글루코스가 모두 소모된 후부터 5 시간마다 6 회 측정하였다. 실험 결과는 반복실험의 평균과 데이터간 의 표준편차인 error bar로 나타내었다 (Fig 3). 실험에 사용 된 균주 YHS04는 같은 글루코스 농도를 공급해 주었을 때 30
oC에서 배양시작 후 Fig. 3(a), Fig. 3(b)는 14시간, Fig. 3(c), Fig. 3(d)는 12시간부터 더 높은 균주 생장을 보였으며, 글루 코스는 37
oC 에서 배양한 Fig. 3(a), Fig. 3(c)에서 30
oC 에서 배 양한 Fig. 3(b), Fig. 3(d)보다 각각 2시간, 7시간 더 빨리 소모 되었다. 에탄올은 배지 내 글루코스 농도가 같은 배양 조건 에서는 비슷한 수준으로 증가 및 유지되었지만, 10 g/L의 글 루코스에서 생산된 에탄올 농도는 최대 4.77 g/L이고, 20 g/L 글루코스에서 생산된 에탄올 농도가 최대 8.33 g/L로 높은 농도의 글루코스가 공급되면 에탄올 생산량이 증가되는 것
Fig. 3. Effect of glucose concentration and culture temperature on the production of FAEE. White circle is ethanol concentration, black
circle is glucose concentration, triangle is O.D. 600nm , gray bar is the amount of FAEE was produced by strain YHS04. Ethanol, glucose
concentration, and O.D. 600nm value were measured at two-hour intervals until glucose concentration dropped to 0. Then all indicators
were measured six times at five-hour intervals. (a): Cultivation at 10 g/L glucose and 37 o C, (b): 10 g/L glucose and 30 o C, (c): 20 g/L
glucose and 37 o C, (d): 20 g/L glucose and 30 o C.
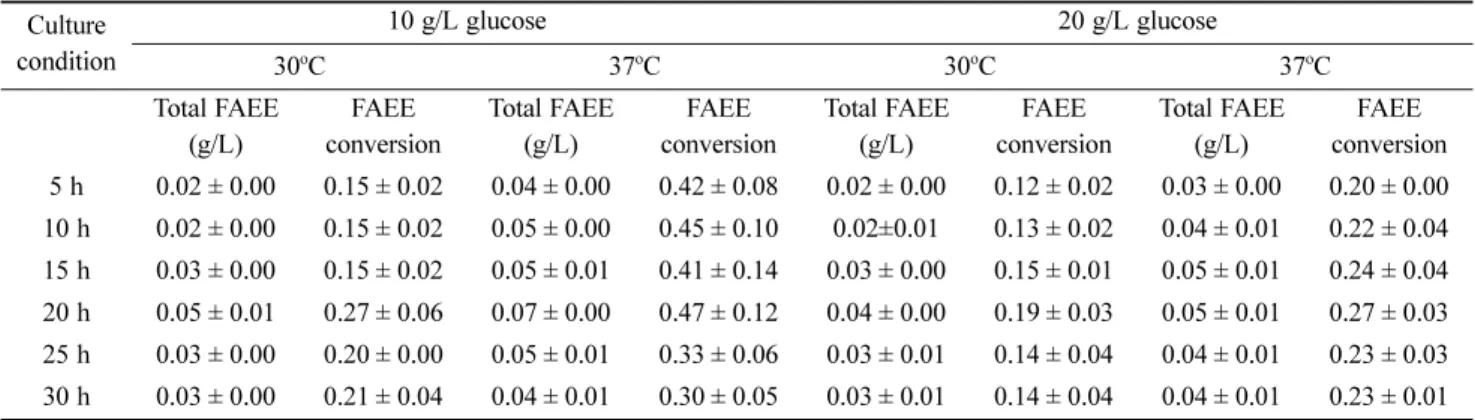
을 확인하였다. FAEE 생산량 검증을 위해 전체 지방산에 대 한 FAEE 전환율을 계산하여 Table 2로 나타내었다. 그 결과 10 g/L 또는 20 g/L의 글루코스 농도에서 배양을 하였을 때, FAEE 생산량은 30
oC 보다 37
oC에서 각각 1.40배 또는 1.74 배 높은 것을 확인하였다. 37
oC에서 배양하였을 때 20시간 이후 배지 내 에탄올과 지방산이 10 g/L 글루코스에서는 4.31 g/L ± 0.11, 0.09 g/L ± 0.04 이고, 20 g/L 글루코스에서는 7.92 g/L ± 0.61, 0.14 g/L ± 0.01로 충분한 에탄올과 지방산이 존재하였음에도 불구하고 두 가지 경우의 FAEE 생산량의 차이보다 FAEE 전환율의 차이가 큰 것을 보인다. 이것은 atfA유전자가 30
oC 보다는 37
oC에서 더 높은 활성을 나타내
기에 FAEE 전환율의 차이를 보인 것이라 생각한다 [27]. 따 라서 FAEE 생산에 있어 37
oC 배양온도에서 10 g/L 농도의 글루코스 공급의 조건이 최적의 조건이라 생각한다.
3.3. 지방산 개시단계 및 베타 옥시데이션 단계 관련 유전 자들의 과발현 영향확인
앞선 실험을 통해 에탄올의 양이 FAEE 생산에 있어 제한 요 인이 아닐 것이라 생각하고 지방산 대사회로의 개시단계와 베타 옥시데이션 단계의 유전자들을 조절한 균주들을 사용 하여 FAEE 생산의 향상 여부를 확인하였다. 배양에 사용된 균주는 control 균주 YHS05 (ΔfadE, pdc, adhB, atfA)에 지방 Table 2. Total FAEE production and FAEE conversion of total fatty acid
Culture condition
10 g/L glucose 20 g/L glucose
30oC 37oC 30oC 37oC
Total FAEE (g/L)
FAEE conversion
Total FAEE (g/L)
FAEE conversion
Total FAEE (g/L)
FAEE conversion
Total FAEE (g/L)
FAEE conversion 5 h 0.02 ± 0.00 0.15 ± 0.02 0.04 ± 0.00 0.42 ± 0.08 0.02 ± 0.00 0.12 ± 0.02 0.03 ± 0.00 0.20 ± 0.00 10 h 0.02 ± 0.00 0.15 ± 0.02 0.05 ± 0.00 0.45 ± 0.10 0.02±0.01 0.13 ± 0.02 0.04 ± 0.01 0.22 ± 0.04 15 h 0.03 ± 0.00 0.15 ± 0.02 0.05 ± 0.01 0.41 ± 0.14 0.03 ± 0.00 0.15 ± 0.01 0.05 ± 0.01 0.24 ± 0.04 20 h 0.05 ± 0.01 0.27 ± 0.06 0.07 ± 0.00 0.47 ± 0.12 0.04 ± 0.00 0.19 ± 0.03 0.05 ± 0.01 0.27 ± 0.03 25 h 0.03 ± 0.00 0.20 ± 0.00 0.05 ± 0.01 0.33 ± 0.06 0.03 ± 0.01 0.14 ± 0.04 0.04 ± 0.01 0.23 ± 0.03 30 h 0.03 ± 0.00 0.21 ± 0.04 0.04 ± 0.01 0.30 ± 0.05 0.03 ± 0.01 0.14 ± 0.04 0.04 ± 0.01 0.23 ± 0.01
Fig. 4. production of FAEE by recombinant strains of E. coli. White circle is ethanol concentration, black circle is glucose concentration, triangle is O.D. 600nm , gray bar is the amount of FAEE was produced by recombinant strains. Ethanol, glucose concentration and O.D. 600nm value were measured from 2 hours after the start of cultivation at one hour intervals until ethanol was produced, and then were measured at two-hour intervals until glucose concentration dropped to 0. After that all indicators were measured six times at five-hour intervals. (a):
strain YHS05, (b): strain YHS06 (c): strain YHS07, (d): strain YHS08.
산 대사회로의 베타옥시데이션 단계 유전자 과발현한 균주 YHS06 (ΔfadE, pdc, adhB, atfA, fadD), 지방산 대사회로의 개 시단계의 유전자를 과발현 균주 YHS07 (ΔfadE, pdc, adhB, atfA, accABCD, fabD), 두 단계 유전자들을 모두 과발현한 균 주 YHS08 (ΔfadE, pdc, adhB, atfA, accABCD, fabD, fadD)를 사용하였다. 실험결과는 3.2와 같이 같은 조건에서 2회 반복 진행하였고, 배양시작 후 2시간마다 샘플링하여 OD, 글루코 스 잔량과 에탄올 생산량을 측정하였고 FAEE는 글루코스가 모두 소모된 후부터 5시간마다 6회 측정하였다. 실험 결과는 반복된 실험의 평균으로 나타내었고, error bar는 데이터간의 표준편차로 하였다. 각 균주에 대한 결과를 Fig. 4에 나타냈 다. 균주 YHS08 배양을 통해 약 0.198 g/L의 FAEE를 생산하 였고, FAEE의 구성은 sFAEE인 ethyl laurate (C12:0) 0.02%, ethyl myristate (C14:0) 4.5%, ethyl palmitate (C16:0) 56.47%
ethyl stearate (C18:0) 3.1%와 usFAEE인 ethyl palmitoleate (C16:1) 4.81%, ethyl oleate (C18:1) 31.1%로 나타났다. YHS04 균주 배양을 통한 FAEE 구성과 비교해보면 sFAEE의 비율 이 증가한 것을 확인할 수 있는데, sFAEE는 디젤 연료의 품 질을 나타내는 세탄값이 높으므로 [28], 균주 YHS08을 통해 생산한 FAEE의 포화지방산 FAEE 비율이 균주 YHS04를 통 해 생산한 FAEE의 포화지방산 FAEE 비율보다 높아 더 좋은 품질의 연료일 것 이라 예상한다. 균주 YHS06은 균주 YHS05 에 비해 FAEE 생산량이 약 1.73배 증가하였는데, fatty acyl- CoA ligase를 암호화하는 fadD 유전자가 지방산의 acyl-CoA 로의 전환을 촉매하는 역할을 하기 때문에 FAEE의 전구체 인 acyl-CoA의 양이 늘어 FAEE 생산량이 증가하였을 것이 라 생각한다 [29]. 균주 YHS07은 균주 YHS05에 비해 FAEE 생산량이 약 2.97배 증가하였는데, 이는 지방산 대사회로의 개시단계 유전자의 과발현으로 인해 전구체인 지방산이 더 많이 생산되어 FAEE 생산량이 증가하였을 것이라 생각한다 [30,31]. 균주 YHS08은 균주 YHS05에 비해 FAEE 생산량이 약 3.88배 증가하였으며, YHS06균주 보다는 FAEE 생산량 이 약 2.25배, 균주 YHS07 보다는 FAEE 생산량이 약 1.31배 증가한 것을 확인하였다. 종결단계 유전자 (tesA)를 과발현 하여 0.037 g/L의 FAEE를 얻은 다른 연구결과에 비해 본 연 구에서는 지방산 대사과정에 관여하는 다양한 유전자를 조 절하여 약 5.32배의 FAEE를 생산하였다. 그리고 지방산 대 사회로의 개시 및 베타 옥시데이션 단계의 조절을 통해 이전 에 시도되지 않은 조합을 사용하여 FAEE의 생산량을 증가시 켰다.
4. CONCLUSION
바이오 디젤로 사용되는 FAME 합성의 단점을 해결하고자 대장균에서 FAME의 대체제로 사용되는 FAEE를 생합성하 고 그 양을 증가시키기 위해 외래 유전자들을 도입하였다.
FAEE 생산의 재료가 되는 에탄올이 약 10.8배 증가한 것을 확인하였고, 글루코스 농도와 배양온도를 조절하여 FAEE
생산에 최적의 배양조건을 결정하였다. 그리고 지방산 대사 회로의 개시단계와 베타 옥시데이션 단계를 동시에 조절한 균주는 control 균주 대비 FAEE 생산량이 최대 3.88배 증가 함을 확인하였다. 또한 지방산 대사회로의 개시단계 유전자 를 과발현한 균주에 비해 FAEE 생산량이 최대 2.25배 증가 하였고, 베타 옥시데이션 단계 유전자를 과발현한 균주에 비 해 FAEE 생산량이 최대 1.31배 증가하였다. 따라서 대장균 의 지방산 대사회로의 개시단계 또는 베타 옥시데이션 단계 를 각각 조절하는 것 보다 동시에 조절하였을 때 FAEE 생산 량 향상에 효과적임을 확인하였다.
Acknowledgements
이 논문은 2019학년도 인하대학교의 지원 및 정부(과학기술정 보통신부)의 재원으로 한국연구재단의 지원을 받아 수행된 연 구임 (No. 2015R1C1A2A01052672).
REFERENCES
1. Nema, P., S. Nema, and P. Roy (2012) An overview of global cli- mate changing in current scenario and mitigation action. Renew- able Sustainable Energy Rev. 16: 2329-2336.
2. Ramanathan, V. (1988) The greenhouse theory of climate change:
a test by an inadvertent global experiment. Science 240: 293-299.
3. Nejat, P., F. Jomehzadeh, M. M. Taheri, M. Gohari, and M. Z. A.
Majid (2015) A global review of energy consumption, CO2 emis- sions and policy in the residential sector (with an overview of the top ten CO2 emitting countries). Renewable Sustainable Energy Rev. 43: 843-862.
4. Cazenave, A. and G. L. Cozannet (2014) Sea level rise and its coastal impacts. Earth's Future 2: 15-34.
5. Merkx-Jacques, A., H. Rasmussen, D. M. Muise, J. J. Benjamin, H. Kottwitz, K. Tanner, M. T. Milway, L. M. Purdue, M. A. Scaife, and R. E. Armenta (2018) Engineering xylose metabolism in thraustochytrid T18. Biotechnol. Biofuels. 11: 248.
6. Qiao, K., T. M. Wasylenko, K. Zhou, P. Xu, and G. Stephanopou- los (2017) Lipid production in Yarrowia lipolytica is maximized by engineering cytosolic redox metabolism. Nat. Biotechnol. 35:
173-177.
7. Sarve, A., S. S. Sonawane, and M. N. Varma (2015) Ultrasound assisted biodiesel production from sesame (Sesamum indicum L.) oil using barium hydroxide as a heterogeneous catalyst: compara- tive assessment of prediction abilities between response surface methodology (RSM) and artificial neural network (ANN). Ultra- son. Sonochem. 26: 218-228.
8. Patel, R. L. and C. Sankhavara (2017) Biodiesel production from Karanja oil and its use in diesel engine: A review. Renewable Sus- tainable Energy Rev. 71: 464-474.
9. Morrison, W. R. and L. M. Smith (1964) Preparation of fatty acid methyl esters and dimethylacetals from lipids with boron fluoride–
methanol. J. Lipid Res. 5: 600-608.
10. Kim, H. M., T. U. Chae, S. Y. Choi, W. J. Kim, and S. Y. Lee (2019) Engineering of an oleaginous bacterium for the production of fatty acids and fuels. Nat. Chem. Biol. 15: 721-729.
11. de Jong, B. W., S. Shi, J. O. Valle-Rodríguez, V. Siewers, and J.
Nielsen (2015) Metabolic pathway engineering for fatty acid ethyl ester production in Saccharomyces cerevisiae using stable chromo- somal integration. J. Ind. Microbiol. Biotechnol. 42: 477-486.
12. Gao, Q., X. Cao, Y.-Y. Huang, J.-L. Yang, J. Chen, L.-J. Wei, and Q. Hua (2018) Overproduction of fatty acid ethyl esters by the ole- aginous yeast Yarrowia lipolytica through metabolic engineering and process optimization. ACS Synth. Biol. 7: 1371-1380.
13. Yu, A., Y. Zhao, J. Li, S. Li, Y. Pang, Y. Zhao, C. Zhang, and D.
Xiao (2020) Sustainable production of FAEE biodiesel using the oleaginous yeast Yarrowia lipolytica. MicrobiologyOpen e1051.
14. Rahman, Z., B. H. Sung, J. Nawab, M. F. Siddiqui, A. Ali, A. Ger- aldi, and S. C. Kim (2019) Enhanced Production of Fatty Acid Ethyl Ester with Engineered fabHDG Operon in Escherichia coli.
Microorganisms 7: 552.
15. Kalscheuer, R., T. Stölting, and A. Steinbüchel (2006) Micro- diesel: Escherichia coli engineered for fuel production. Microbiol- ogy 152: 2529-2536.
16. Steen, E.J., Y. Kang, G. Bokinsky, Z. Hu, A. Schirmer, A.
McClure, S. B. Del Cardayre, and J. D. Keasling (2010) Microbial production of fatty-acid-derived fuels and chemicals from plant biomass. Nature 463: 559-562.
17. Magnuson, K., S. Jackowski, C. O. Rock, and J. E. Cronan (1993) Regulation of fatty acid biosynthesis in Escherichia coli. Micro- biol. Mol. Biol. Rev. 57: 522-542.
18. Cronan Jr, J. E. and G. L. Waldrop (2002) Multi-subunit acetyl- CoA carboxylases. Prog. Lipid Res. 41: 407-435.
19. Link, A. J., D. Phillips, and G. M. Church (1997) Methods for gen- erating precise deletions and insertions in the genome of wild-type Escherichia coli: application to open reading frame characteriza- tion. J. Bacteriol. 179: 6228-6237.
20. Jopseph Sambrook, G., Michael R., (2012) Molecular cloning : a laboratory manual / Michael R. Green, Joseph Sambrook. 4nd ed., Cold Spring Harbor, NY, USA.
21. Marchetti, J. and A. Errazu (2008) Esterification of free fatty acids using sulfuric acid as catalyst in the presence of triglycerides. Bio-
mass Bioenergy 32: 892-895.
22. Lee, C. M., B. Trevino, and M. Chaiyawat (1996) A simple and rapid solvent extraction method for determining total lipids in fish tissue. J. AOAC Int. 79: 487-492.
23. Ma, F. and M. A. Hanna (1999) Biodiesel production: a review.
Bioresour. Technol. 70: 1-15.
24. Gyu Beom, J. (2017) Metabolic Engineering of Escherichia coli for the production of FAEE: Engineering of FAEE production pathway for the improvement of FAEE production by FAS gene introduction into Escherichia coli. M. D. Thesis. University of Inha, Incheon, Korea.
25. Ohta, K., D. Beall, J. Mejia, K. Shanmugam, and L. Ingram (1991) Genetic improvement of Escherichia coli for ethanol production:
chromosomal integration of Zymomonas mobilis genes encoding pyruvate decarboxylase and alcohol dehydrogenase II. Appl. Envi- ron. Microbiol. 57: 893-900.
26. Lewicka, A., J. Lyczakowski, G. Blackhurst, C. Pashkuleva, K.
Rothschild-Mancinelli, D. Tautvaišas, H. Thornton, H. Villan- ueva, W. Xiao, and J. Slikas (2014) Fusion of pyruvate decarboxy- lase and alcohol dehydrogenase increases ethanol production in Escherichia coli. ACS Synth. Biol. 3: 976-978.
27. Stöveken, T., R. Kalscheuer, U. Malkus, R. Reichelt, and A. Stein- büchel (2005) The wax ester synthase/acyl coenzyme A: diacyl- glycerol acyltransferase from Acinetobacter sp. strain ADP1:
characterization of a novel type of acyltransferase. J. Bacteriol.
187: 1369-1376.
28. Knothe, G. (2009) Improving biodiesel fuel properties by modify- ing fatty ester composition. Energy Environ. Sci. 2: 759-766.
29. Duan, Y., Z. Zhu, K. Cai, X. Tan, and X. Lu (2011) De novo bio- synthesis of biodiesel by Escherichia coli in optimized fed-batch cultivation. PLoS One 6: e20265.
30. Davis, M. S., J. Solbiati, and J. E. Cronan (2000) Overproduction of acetyl-CoA carboxylase activity increases the rate of fatty acid biosynthesis in Escherichia coli. J. Biol. Chem. 275: 28593-28598.
31. Zhang, X., A. Agrawal, and K.Y. San (2012) Improving fatty acid production in Escherichia coli through the overexpression of mal- onyl coA Acyl carrier protein transacylase. Biotechnol. Prog. 28:
60-65.