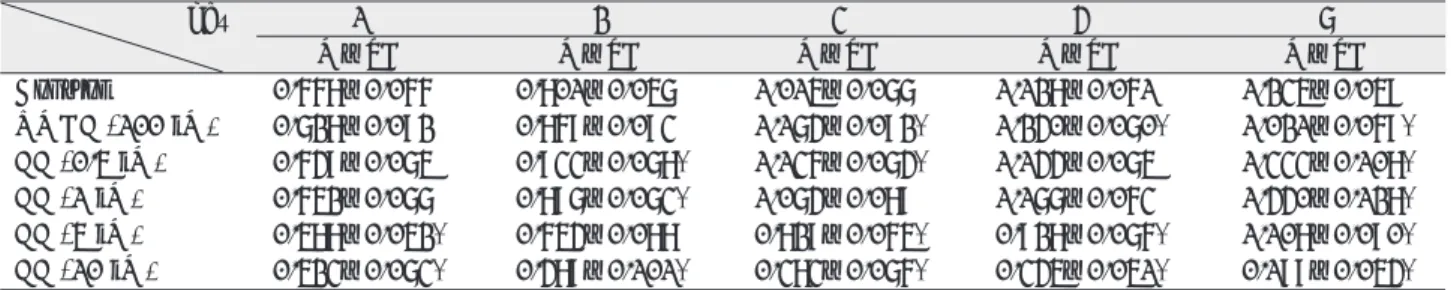
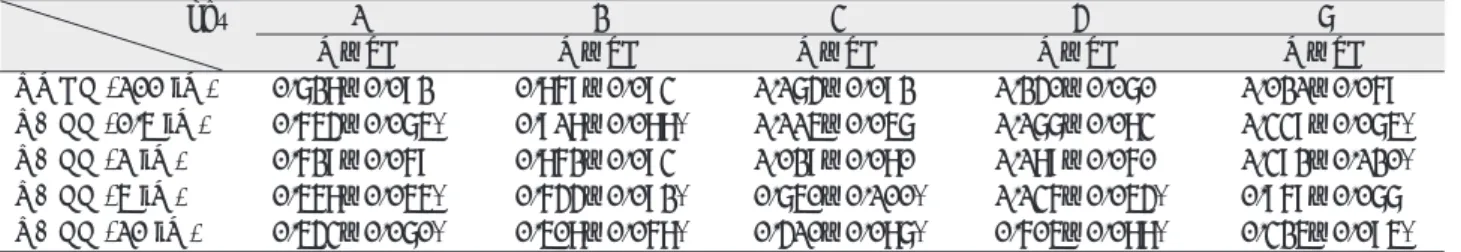
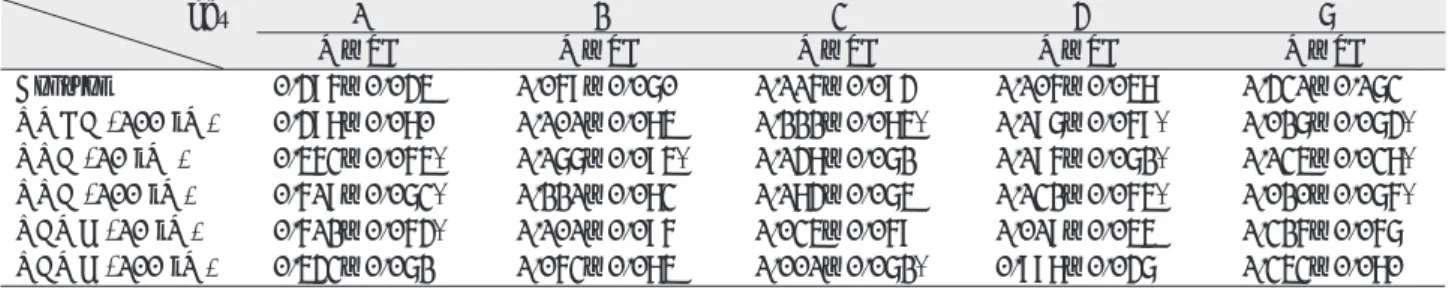
Ⅰ. INTRODUCTION
Normal human oral keratinocyte (NHOK) is keratinized stratified squamous epithelium, and its proliferation is primarily accomplished by divi-
sion of basal cell which exists in the lowest layer like the epithelium of the general skin. The oral epithelium has different characteristics from epithelium of general skin such as hairlessness, no sweat glands, and always being wet. It shows 김인수∙이 원∙김성훈*∙최 봄**
가톨릭대학교 의정부성모병원 치과 구강악안면외과, *교정과, **보철과
N-methyl-D-aspartate 수용기의 다양한 조절이 일차 배양된 정상사람구강각화세포의 생존에 미치는 영향