Kor. J. Appl. Microbiol. Bioeng.
Vol. 13, No. 1, 19-25! 1985)
효모 HIS5 유전자에 관한 연구
- Saccharomyces cerevi si ae HIS 5 유전자의
5' 상류영역의 염기배열 -
鄭東孝 · 西臨淸二*. 大*훌泰治*
중앙대 학교 농과대학 식품가공학과
*오사카대학 공학부 발효공학과 (1984 년 12 월 29 일 수리)
S tudies on the HIS 5 Gene of Yeast
- The nucleotide sequence of 5' upstream region of the HIS 5 Gene of Saccharomyces cerevisiae
Dong Hyo Chung, Kyoni Nishiwaki *and Yasuji Oshima *
Department of Food Science and Technology.
College of Agriculture. Chung-ang University
* Department of Fermentation Technology.
Facu\ty of Engineering. Osaka University (Received December 29, 1984)
The H!S5 gene 01 Saccharomyces cereuisiae host was encoded histidinol phosphate aminotranslerase (E.C“ 2.6. I ‘ 9). The H!S5 gene 01 Saccharomyces cereuisiae was c10ned on plasmid pSH 530. This gene mighted be transcripted lrom a promoter 01 yeast gene both in E. coli and yeast hosts‘
We have determined the nucleotide sequence 01 the yeast H!S5 gene and its 5 ’ and 3 ’ f1anking sequences. There are no large dillerences between the relative levels 01 H!S5 mRNA molecules with different 5 ’ termini in represent and derepressed cell. In the DNA sequence upstream lrom the 5' termini 01 H!S5 mRNA we have lound live c10sely related copies 01 a 9 base pair sequence. The sequence is also repeated in the 5 ’ noncoding regions 01 H!SI, H!S3, HIS4, H!S5 and TRP5. Closely related sequence are not lound f1anking repeat sequence plays a role in the regulation 01 amino acid biosynthetic genes subject to the general amino acid control
19
유전물칠의 본체가 DNA 이다 따라서 이 DNA 의 화학분해법(1, 2)과 DNA polymerase 를 이용하여 3 ’ 염기배열을 결정하여 유전정보를 직접 해석하지 않 방향에의 DNA 사슬의 신장을 특이적인 엽기로 정 으면 안된다 DNA 엽기배열의 결정은 생영 과학의 지시키는 효소볍" ‘”이 있 a 나 어느 것이나 polyac- 기초분야에 있어서 뿐만아니라 biotechnology 둥의 rylamide gel 전기영동엽우로 각 엽기를 분리한다.
응용분야에서도 필수의 연구과제이다. 최근 효모의 HISl-5 유전자중 HISl ' 이 • HI 유전자 DNA 엽기배열 결정에는 대별하여 방사션 S3"'. HIS 4"'" 유전자의 엽기배열이 결정되었고 동위원소 "p 로 표지한 DNA를 어느 염 기에 특이 이외에도 TRP5 '인• LEU 2,00'. PGK ''''. cytochro- 적£로 작용하는 시약으로 DNA 사슬을 절단하는 ome'U" SUC 2'‘’1 등의 유전자의 염기배열이 차츰
톨톨..., Yeast
c::::그 E. coli
¢따TIl1l 2-umDNA pBR 322
pSH530 17.4Kb
3 3
걱 Pst 1 9 'Zl Bgll1
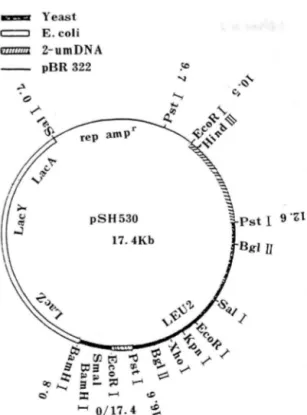
Fig. L Structure and restriction endonuc\ease map of plasmid pSH 530.
The double line represnts lac' ZYA gene of E. coli. The heavy line and the solp lines represant yeast LEU2 gene and 2 umDNA. The thin line shows DNA derived from pBR322. ampr; Ampicillin resistance.
밝혀지고 있으나 HIS5 유전자에 관한 연구는 Os- hima(U) 연구외에는 찾아 볼 수 없다.
여기에서는 HIS 5 유전자 DNA 의 염기배열을 염 기 특이적인 화학수사를 행하여 수사염기위치에서 DNA 사슬을 절단하는 Maxam-Gilbert 방법이)에 따 라 결정하여 그 결과의 일부를 보고하며 아울러 다 음에 곧 이어 DNA 염기배열의 어느 영역에서 mR-
NA 와 단백질이 합성되는가 즉 전사개시점과 번역 개시점을 결정하려고 한다.
실험재료 및 방법
Plasmid pSH 530 과 제한효소
HIS 5 유전 자함유 plasmid는 효모 2 μm plasmid 의 복제 기 점 과 대 장율 lac’ Z 유전 자에 promoter 를 부여시킨 잡종효크 plasmid (pSH 530, marker LEU 2 )로서 BamHI 절단부위에 HIS5 gene 을 삽입한
17.4 kb의 크기 의 것 이 다 (Fig. 1 ). 이는 Casadaban 이 구축한 것으로 lac ’Z의 제 8 번째 의 codon 에서 lac operon 의 구조유전자가 있고 이 부분에서 단백 질의 N 말단을 만드는 promoter fragment 가 삽입된 reading f rame 이 있으면 lac Z 유전자가 발현되는 unique 한 vector 이 다.
제 한효소 Ava II, S au 3 A는 New England Bîo- labs제 품, HinC, Hind’ m 는 Takara Shuzo Co.
Ltd 제품을 사용하였다.
Polyacrylamide gel 에서 DNA 의 회수
영기배열을 결정하려는 DNA 단편을 제한효소로 plasmid 에서 끊어 내어 polyaCI'ylamide gel 전기영동에 서 분리한 다음 polyacryamide gel을 자외선 확인으 로 DNA 밴드를 절단하여 siliconized glass wool 을 채우고 끝을 벌봉한 pipetteman 에 넣고 siliconized 유리막대기(흑은 spatule) 로 1 mm' 정도로 깨뜨린 다. 여 기 에 0.5 M NH‘-acetate-lO mM Mg-acetate -0.1%SDS-0.1 mM EDTA 용액으로 37"C -420C 에 서 하룻 밤 방치 추출시 킨다. 다음에 chip 끝을 가위 로 끊어 내고 Eppendorf tube에 넣고 저속원섬 분리 (2,000-4,000 rpm) 한다 용출된 DNA 용액을 Epp- endorf tube 에 옮겨 ethanol (-20'C ) 침 전 을 행 하고 50mM Tris-HCl(pH 8.0-1 mM EDTA 로 포화된 phenol : chloroform (1 : 1) 으로 처 리 하여 다시 et - hanol 칭전을 2 - 3 회 행하고, Maxam-Gilbert 방법 이}에 따라 DNA 의 영기배열을 결정하였다.
Bacterial alka
’
ine phosphatase ~~ 2.1얻어 진 DNA 단편 (1 μg) 을 200 μl 의 50 mM Tris -HCl (pH 8.0) 의 완충용액에 녹여 대장균 alkaline phosphatase ( 5 μg, 0.2 unit) 용액 을 가하여 37'C, 30분간 반응시켜 DNA 단편의 5 '말단의 인산을 제 거하였다. 다음에 50 mM Tris-HCl (pH 8. 이 -1 mM EDTA 흐 포화시 킨 phenol chlorform ( 1 1 ) 으 로 처리하여 반응이 끝난 효소를 실활시키고 ether 로 phenol 을 제거한 다음 감압하에서 건조시키고 다 시 ethanol (-20"C ) 칭전으로 정제하였다 칭천으로 얻어 진 DNA 단편 (5' 말단으로 1-50 p moI) 을 40 μl 의 무균수에 녹였다.
T 4 polynucleotide kinase 에 의한 5 ' 말단의 32p 의 표지
위에서 얻은 40 μl 의 dephosphorylated DNA ( 1 -10 p moI) 에 5 μl 의 완충용액 (500 mM Tris-HCl
(pH 7.6)-100 mM MgCl, -50mM DTT(dithiothreitoI) -1 mM spermidine(Sigma)-1 mM EDTA) 5 μl 의 (r
"P) ATP (3, 000 Ci/m mol, New England Nuclear,
Vol.13, No.l,
。 r Amersham, Internation Ltd. Bucks, England) 용 액 맞 1 ml 의 T 4 polynucleotide kinase (3 unit, Ta- kara Shuzo Co.) 용액 을 첨 가혼합하여 37"C 에서 60 분간 반응하였다.
DNA 단편의 5' 말단。 1 f1ush의 경우에는 마찬가지 로 대 장균 alkaline phosphatase 로 처 리 하여 침 전을 얻고 이 침전 DNA 단편에 무율수 5 μl 와 20 mM Tris-HCI (pH 9.5) -1 mM spermidine-O. 1 mM ED- TA 용액 35μl 플 가하여 용해시키고 이것을 90 "C에 서 2 분간 실활하고 나서 급냉시켰다. 곧 5μl 의 ,500 mM Tris-HCI (pH 9.5) -100 mM MgCI,-50 mM
DTT -50% glycer이용액. 5 μl 의 (r- "P)ATP (50 p. mole에 만족치 옷한 경우에는 carrier 를 청가하 였다) 맺 1 μl 의 T 4 polynucleotide kinase를 가하 여 37 "C에서 30분간 반응하였다. 그리고 200μl 의
?.5 M NHracetate 용액을 가하고 반응을 정지시
키고 다음 1μg의 tRNA ( 1 mg/ml, Sígma) 를 가하 여 ethanol 침천을 하였다. 이를 0.3 M Na-acetate 용액에 녹여 750 μl 의 ethanol 로 2 회 원심하였다.
양 5 ' 말단이 표지된 DNA 단편의 제한효소에 의 한 secondary cleavage 의 분리와 회수
’위에서 침전 건조회수된 양 5' 말단이 표지된 .<5' end labeled) DNA 단편을 200 μl 의 증류수에 녹 여 1/ 10 량의 10 배 농도의 TA 완충용액과 1/10 용 (容)의 1 mg/ml BSA (혈청알부민)용액 및 예상 제 한효소(20-30 unit)를 가하여 반응을 하였다. 반응 후 ethanol 침전을 행하고 이것을 30μl 의 Tris-HCI
(pH 7.5)-1 mM EDTA 용액에 녹이고 5 μl 의 색소 용액 (10 mM EDTA-O. 05% (w/v) bromophenol blue -60% (w/v) sucrose) 을 가하여 6 % polyacrylamide disc gel에 서 150-200 V, 2 - 3 시 간 전 기 영 동 하 였다. 천기영옹이 끝난다음 gel 을 꺼내어 EtBr 용
액 (0.5 μg/m\) 에서 30분간 염색 후 자외션을 조사
하면서 말단이 label 된 필요한 DNA 단편을 함유한 부분을 gel 에서 끊어 내었다. 이것을 DNA 회수의 경우와 같이 300 μl 의 용출액 ω.5M NH‘-acetate- 10 mM Mg-acetate-1 mM EDTA-O. 1 % SDS- 20 μg/ml tRNA) 를 가하여 37"C에서 하룻 밤 땅치추출 하고 이를 Eppendorf tube 에 회수하였다. 이와같이 회수된 DNA 용액을 ethanol 칭전과 마찬가지로 강 압 건조시키고 방사활성을 Cerenkov 계수에 의하여 scintillation counter (Beckmarr L-250) 에서 측정 하 였다. 측정 후 칭전을 35μl 의 우균수에 녹였다.
염기 특이적인 화학수사
CD Guanine 의 특이반응 (G 시료)
Guanine 잔기에서의 한정 (限定) 분해는 Eppendorf
21
tube 에 1 μl 의 TE 완충용액에 녹여 초음파 처리한 calf thymus DNA (65 mg/ml, Sigma 이 i5} sonicat ed ctDNA) 용액 200 μl 의 dimethyl sulfate (DMS) 완충 용액 (50 M sodium cacodylate (pH 8.0)-1 mM ED- TA 용액〕 및 5μl 의 5' 말단이 label 된 DNA 용액을 가하였다. 그리고 다시 lμl 의 DMS 용액을 가하여 23 "C에서 4 분간 반응하였다. 반응 후 50μl 의 D-
MS 반응정 지 용액 (1. 5 M Na-acetate-(pH 7.0) -1 M
β-mercapto ethanoI) 을 가하여 반응을 정 지 시 키 고 곧 750 μl 의 뱅냉 ( - 10"C) ethanol 을 가하여 침천 건조시키고 다음의 piperidine 조작을 행하였다.
(g) Guanine 과 adenine 으| 특이반응 (A+G 시료) Guanine과 adenine 잔기 (C+T) 의 한정분해는 Ep- péndorf tube 에 34 μl 의 무균수와 l μl 의 ctDNA
(sonicated calf thymus DNA) 용액 빛 10 μl 의 5' 말단 label 된 DNA 용액을 가하고 1 μl 의 1 M ace- tic acid 용액을 첨가한 다음 45"C에서 15분간 반응 하였다. 반응 후 200 μl 의 hydrazine 정지용액 (0.3 M Na-acetate-0.1 mM EDTA 용액)을 가하여 잘 혼합 하여 반응을 정지시키고 @과 같이 ethanol 침전을 한 다음 다음의 piperidine 조작을 행하였다.
@ Cytosine 과 thymidine 의 특이반응(C+T 와 c 의 반응)
Cytosine 과 thymidine 의 잔기의 한정분해는 Epp- endorf tube 에 1 μl 의 sonicated ctDNA 용액과 9 μl 의 우균수 및 10 μl 의 5' 말단 label 한 DNA 용액 을 가하고 30μl 의 hydrazine (Piere Chemical) 을가 한 후 23 "C에서 7 분간 반웅하였다. 반응 후 200 μ l 의 hydrazine 정 지 액 (0.3 M sodium acetate-0.1 mM EDTA) 을 가하여 반응을 정지시키고@와 같이 75μl 의 ethanol 로 칭전, 진공건조하였다.
@ Cytosine 의 한정분해 (C 시료)
Cytosin 의 잔기의 한정분해는 역시 Eppendorf tube 에 1 μl 의 ctDNA용액과 5 μl 의 5 M NaCI 용 액 그리고 5 μl 의 5' 말단 label 된 DNA 용액을 가 하고 30 μl 의 hydrazine 을 첨가한 다음 23 "c에서
7 분간 반응하였다.@와 같이 hydrazine 반응정지 ethanol;침 천 을 행 하고 이 하 piperidine 반응의 조작을 행하였다.
@ Piperidine 반 응
Adenine 과 guamne 잔기 cytosine 과 thymidine 잔 기 빛 cytosine 잔기의 특정한 분해는 어느 것이나 200μl 의 0.3 M Na-acetate 용액을 첨가하여 반응 을 정지시키고 곧 750μl 의 맹냉 ( - 10"C) ethanol을 가하여 칭 천, 건조시 켰다. 강압건조한 DNA 에 100 ul 의 1 M piperidine 용액올 가하여 DNA 흘 녹인다
음 90"C 에서 30분간 가열한 다음 각각을 동결건조 (Yamato RC-ll. Hitachi 160vp-S 진공펌 프) 를 60 분간 행하고 20μl 의 무균수에 녹였다. 동결건조된 DNA 표풍의 방사능을 Cerenkov 계수로 부터 액체
scintillation counter 로 측정 하였다. 다음에 적 땅한 량의 엽기배열결정용 염색액 (8% deionid formamide 10 mM NaOH-l mM EDTA 0.1% xylenecyanol F-
FCxC 로 약함) -0.1 % BPB (bromophenol blue) J 1
μl 당의 DNA 시료의 방사활성이 10,000-20,000 c-
pm 가 되게 가하여 녹였다. 이 각각의 용액을 90"C 에서 1 분간 가열하고 곧 맹수중에서 냉각하여 저 속원심하였다.
염기배월 컬정용 gel 의 작제
잘 씻은 유리판(가로 40 cm, 세로 30 cm, 두께 4 mm) 을 두장을 준비하고 한 장은 미리 내측연을 sili - conize 하여 두고 다흔 한 장은 acetone 으로 내측을 잘 닦아 둔다. 이 두장의 유리판의 사이에 두께
0.3 mm 의 spacer (양측 저부)를 끼우고 cnp 으로 고 정시컨다. 영기배열결정용의 gel 은 "P 으로 표지 된 DNA 의 말단에서 읽을 수 있는 벙위에 따라 20
%, 8 %, 6 % gel 을 상법에 따라 작제하였다.
G 허전기영동
하룻 밤 방치하여 둔 gel 에서 comb 를 제거하고 lane 에 냥아 있는 acrylamide를 천기영동 완충용액 (TB 완충용액)으로 잘 씻어낸 다음 새로이 만든 천기영 동 완충용액을 가하여 약 1500 V 에서 2 - 3 시간 전(前)영동을 하였다. 천영동 후 DNA시료의 일 부(보통 1 - 2 μ J)를 미세한 모세관으로 apply
하였다. 본 영동은 1500-2000 V 에서 행 하고 6%
gel 은 1 % xylene cyan이 이 gel 의 상단에 서 30 cm 이동한 시청에 8 % gel 은 bromo phenol blue 가 38 m 이동한 시점에 (끝 부분) , 그리고 20% gel 은 bromophenol blue 가 15 cm 이동한 시정에서 전기영 동을 정지하였다 "P 로 표지된 DNA 의 말단에서 읽을 수 있는 범위는 20 % gel 에서 1-40 엽기쌍,
8 % gel 에서 25-130 염기쌍, 6 % gel 에서 100- 200 염기짱이다.
Autoradiography와 현상
천기영동종료후유리판의 한쪽을 spatule 으로 주 의 갚게떼어 내고 gel 은 saran w-ap 으로 주륨이 생 기 지 않게 잘 싼다. 노출용 film holder (Eastman Kodak Com p. ) 에 유리판을 넣고 그 위에 X선 필 릎 (K얘ak XS- 1) 을 얹고 필요에 따라 중강치를 넣어 holder 를 crip 으로 고청시킨 다음 - 80"C 에서 방사능의 양에 따라 수얼 간 노출하였다. 방사능이
20,000 cpm 인 경우에는 1 일 노출로 가능하였다.
2,000 cpm 의 경 우에 는 intensifier (Kadak) 를 사용할 때는 하루의 노출로도 가능하였다. 현상은 X선 필 릎용의 현상액에 5-10분간 당가두었다가 청지액 (3 % 식초산)에서 현상을 정지하였다. 다음에 x
션 필릎용의 산성 경악 정착제에 핀릎을 맘근 다음 펄릉을 냉수중에서 장시간 씻어 잘 건주시킨 다음 펄름에 현상된 염기배열을 읽었다.
결과및 고찰
HIS6 gene 의 5' 상류의 염기배멸 HIS6
Gene 의 5' 상류의 염기배얼온 Fig.2 와 같다.
Fig.2 와 같이 전체로서 736 엽기쌍을 이루고 있 었 다. 여 기 에 서 directed repeated sequence (•) 가 12곳, inverted repeated seφence ( ... ) 가 세곳 이 었다. 특히 HIS 5 promoter 에 제얼 큰 부분을 차지 하는 open reading frame (= ) 이 존 재 하고 이 와같은 frame 은 TRP 5, HIS 4 gene 에도 몇 개씩 존재하고 실제로 RNA 가 이곳에서 전사하는 것 ξL 로 생각되는 곳이다. 이 엽기배열 중 ATG 배열은 meitnonine 을 code 하고 여기에서 mRNA 에서 poly - peptide 사슬로 번역이 개시되는 번역개시점이다.5' 상류에는 전사개시첨 (cap) 으로 생각되는 점 (-69)과 그리고 RNA polymerase 의 인식배열로 생각되 는 pribnow box (-5'-A -A-T-A-A-T-3' box, 진 핵 생 물의 경우, 원시핵세포의 경우는Hogness box) 가 두 곳 ( -102와 -139) 이 있다. 특히 Fink 둥이 지 적한 general amino acid control 의 consensus seq-
uence(d 부분)가 세 곳이나존재하는 것을 알 수 있
다. 그리고 HIS 3, HIS 4 그리고 HIS5 의 ATG 주 변의 염기배열주변은 Fig. 3 과 같다. - 3 은 다같 이 A 이고 HIS4, HIS 5 의 +14 에서 +8 까지의 엽 기배열은 -G-T-T-T-T-로 C block 은 없었다.
HIS6 gene 의 5' 상류의 공륭배얼
5' 상류의 염기배열중에는세포의 생육환경에 따 른 신호 즉 배지 중의 탄소 에너지원, 혹은 질소화합 물의 종류둥에서 유래되는 신호를 받는 엽기배열이 있고 여기에서 작동하는 기구가 있기 마련이다. 효 모나 Neuroøpora 둥의 진핵미생물에서도 최근에 이 르러 정 (正) . 부(員) . 양(陽) 조절인자에 관계하는 신호천달계가 해영되고 있다. 이 최종단계에서 구 조유천자의 5' 상류부에서 수신에 작용하는 것으로 생각된다.
대개의 아미노산합성에 작용하는 효소구조유천자
Vol.13, No.l, 23
-616 RindIII
AGCτTIAGCTG~AAττcττσ~ACTGTCTTTCGCCGAAACCτsττTGATGACτACτ
-560
TCAτCAATττττττTτττTCτGCCGCA'τTCCAAA~τCAτAACτTTGCAAAAATAAAGGGTAAATGGT7A
a b
-490
AAAATτGττAτCAτAAAτ ... :\A.GG7GACCGG'ττAτAτTGAGACCττTCCTGGACGGτAACτAAτACAGAAGC
..
b a
-420 Sau3A
CAττGG7AATGCAATAAτττATττGATCATGτGACCACGAτCCGGG'τGAGACτATτCAAAAAAGGAGτCA
c
-350
AGCAττGAAAτAAττAAτGAC':AATCCGAAG':-:AATT G7TAGGAGTCAATτG7:'TτTτCCAATGAAT~A
. . • - - + ‘ . . c===
o e • e
-280 c
ATC:GAGATGAC’:AAACτAC:AATTττCAAτAGτ'I'CATGG~AτAGτ GACC7AττAG7GC:-:-:~C=
..
Sau3A d.
GG~TC:G7: GACτ CAC:7 CAAττGATG7:7CττACC:TGACAτ~~CAτACττGAτATIττAτcτcτCACG
---+
*--「-140 묘 d ‘ x ---;:,-야 • RB f .- càP
TτATATAACTTGAAAA~ATGCACACAGτTCτGττCAAτATACCCτCCAAτsτGTAAAAACAGτττTτCC
----
---~ y i … z -z-70
ATTGAτTAC7CτTAAτ’:T Gτ-:TCC'I' GCτAAACCAGCAGτACGτGτGτG':CG":ATAτAττ.AAAATTACACτ -~
l 70
ATGGτττττGAτTTGAAAAGAATτ G7τAGACCAAAAAτTτATAACττGGA.ACC7τATCGCτcτGCAAGAG MetVaLPh~~pLeuLysArgI1eVa lArgP~oLγs Il eτyrAsnL euG1 uP ~oTyrArgCysA1 aArg
AvaII 140
AτGAτ'IT CACCGAGGGTATAτTGCτAGACGCCAATGAAAAτGCCCATGGτCCτACτCCAGTτGAATIGAG As pAspPhe IhrG1uG1γI1eLe uL euA spAlaAsnG1 uAs nAla Ei sGly?ro IhrProVa1GluI, euSer
160 Sau3A
CAAGACCAAτTTACATC G'τTACCCAGATC Lys IhrAs nL e u?~sArgTyrP~oAsp
Fig.2. DNA sequence of 5' upstream region of the HIS 5 gene.
The numbering is relative to the transla tion initiation codon which is designated + 1 . The amino acid sequence of the HIS 5 coding region is printed above the DN A sequence. The locating of two reading frames upstream from HIS 5 are indicated by underlining, with arrow heads marking
‘ the stop codons. Directed repeats are designated by underlining arrows and labeled with capital letters.lnverted repeat pairs are underlined with dotted arrows and labeled with small letters, such as y, y' and p, p' where y; z' were the complement to y, z. HB box are Hogness box and CAP are Ininitiotion of transcription.
의 발현조절에는 몇 개의 아미노산에 공통으로 하 류부는 Fig. 4 와 같이 9 엽기쌍(base pair) 의 반복 여 작용하는 일안조절계(general control system ) 배열이 존재한다고 지적하였다. 이 공통적 배열은 가 있다 효모에서도 arginine, tryptophan, histidine, 바로 5' -A-~'-G-T-G-A-C-T-C-3' 이다.
A
lysine, leucine, isoleucine, valine 에 공통 의 계 가 있다 HIS 5 gene 도 Fig. 1, Fig. 4 와 같 이 -336, - Struhl (15,161 등은 이 일반조절계에 지배되는 아 275 그리고 -205 위치의 세 곳에 반복배열이 있다 미노산 합성효소구조 유전자의 5' 상류영역의 최상 특히 다른 유전자도 상류부에 반복된 공통배열이
Genes A TG environment
10 9 8 7 6 5 4 3 2 -1+1 2 3 4 5 6 7 8 9 H1S3
H1S4 H1S5
Fig.3. Comparison of the ATG enviroment region on HIS3. HIS 4 and HIS 5 gene of yest.
존재하며 그 분포도는 H1S 1, H1S 4 는네 곳 , H1S 3, H1S 5 는 셰 곳이 있다. 따라서 H1S 5 gene 도 역 시 general amino acid control 을 받은 것 23 .. 생각 할 수 있다. General amino acid control 을 받는다
Position of genes. Sequences
HIS 1 -450 A G G T G A C T A 7/9 -435 A T G T G A C T A 8/9 -286 G C G T G A C T C 7/9 -189 A A G T G A G T C 8/9 HIS3 -182 T T A T G C C T C 6/9 -140 Q. Q. A T G A C T C 6/9 - 74 A T G T G A T T T 7/9 HIS 4 -257 A A C T G A C T C 8/9 -245 T A G T G A C T C 8/9 -201 C A G T G A C T C 8/9 - 1 5 0 A T A T G A C T 좌 7/9 HIS 5 - 3 3 6 T A A T G A C T A 6/9
-275 A G A T G A C T A 6/9 -205 T G T T G A C T C 6/9 TRP5 -235 T T G T G A C T G 7/9 -110 으 A A T G A C T k 6/9
* * * *
Consensus: AAG T G A C T C T
Matches/15 7 10 8 15 15 14 13 15 8
LEU2 absent
TRPl absent
Fig.4. A compilation of I'epea ted seperated seq- uences ;in .the 5' flan king region of co-
‘ \
regulatèil amino acid biosynthetic genes in Sac씨aromgces cere8iviae.
Positions which deviate from the consen- sus sequence are underline. The asterrisks
。ver the consensus sequence mark the in- variant positions for the set the number of matches between the consensus seque- nce and the set of 12.
고 생각되는 유전자에는 3 - 4 개의 consensus se- quence 가 존재하는 것을 알 수 있다.
한편 Fig.5 와 같이 general amino acid controJ 을 받지 않는 유천자 예로서 leucine 에 작용하는 L- EU2 유전자에 있어서는 640 염기쌍의 상류에 이르 기까지에나 또 tryptophan 의 합성에 작용하는 TR.-
Pl 유천자에 있어서도 440 염기쌍 상류에 이르거까 지 조사하여도 이와 같은 9 염기배열은 발견되지 않
는 것이 특칭이다. ‘
이상 Fig.4 와 같이 H1S 1, H1S 2, H1S 3, H1S 5 그리고 TRP5의 consensus 는 -A .. ~ .. G- 단프판 도콰 C- 인 것을 알 수 있다. 따라서 9 엽 기쌍 중
.. 600 ... 00 .. 400 .300 .200 .. 100 ATG -4-100 • 200
。
• • •
願J뭔않+
1636-따↓혈h;웅3
1320
- 616 톨-- 톨-..l 떤같쫓+180
- 450 +2493
- 640 다냥 새 떼 ?i
- 440
때
蠻 --
Fig.5. Repeated sequences in the 5' f1anking regions of co .. regulated amino acid bio- syn thetic genes in Saccharomgce8 cere- Vl8lae.
The 9 bp repeats Jisted in Fig. 3 are depicted by soJid squares. The mapped 5' termini of the transcripts are shown as arrows. For H1S 1, the fractionaJ amount of total H1S 1. mRNA areindicated above the arrows for the three major H1S 1 m ..
RNA species. The distance in base pairs from the begining of the coding regions
。 f the four genes is given at the bottom, where+ 1 marks the first nucleotide po- sition of each ATG.
Vol.13, No.l,
적어도 6 엽기짱은 같으며 괄호내의 염기배열 순서 는 15 엽기 중 적어도 13 엽기는 같고 15 엽기가 꼭 같 은 것이 두 곳이나 된다.
그러냐 하냐의 구조유전자가 딸현함에 있어서 그 상류에 있는 모든 반복배열 9 염기쌍이 필요한 것 윤 아니다. 그러나 이들이 결실되면 발현에 이상
{異常)이 생 낀다고 한다(''1 한 예로서 HIS 4 gene
;은 4 개의 반복배열을 가지나 그 중 두 개가 결설되 면 general amino acid concrol 계에 기능을 잃는다 고 한다. 그러나 세개 모두가 결실되연 발현은 정 ι~쩌되고 만다고 한다. 반대로 발현이 행해지는 돌연
변이체에서는 유사한 배열이 생기게 되고, general amino acid control 계에 강응하여도 회복되고 말 것으로믿어 진다.
요 약
Saccharomyces cerevi&iae HIS 5 유전자는 hist- idinol phosphate aminotransferase (EC: 2, 6, 1,9) 를 code 하는 아미노산 합성유전자이다. 이 유천자 는 plasmid pSH 530 에 cloning 되어 E. coli 와 Sacc- lmromyces cereν Ulae 숙주에서 promoter 로서 전 사하였다.
HIS 5 유천자의 총엽 기 수는 7367>> 이 였고 5' 상 류영 역 에 는 긴 reading frame, directed repeat, 천 사개시점, 그리고 Pribnow box 엽기배열이 있었다.
특히 HIS 5 유전자의 ATG 주변 영기배열은 -A-A-A -T-T-A-C-A-C-T-A-T-G-G-T-T-T-T-T-G-A-T- 였으며 C block 윤 없었다.
사 시
25
참고문헌
1. Maxam, A. M.and W.Gilbert: Proc.Natl.Acad.
Sci. , U SA, 74, 560 (1977). USA,
2. Maxam, A. M. and W. Gi\gert Methods Enz- ymol. ,65, 499 (1980)
3. Sanger F., S. Sicklen and A. R. Coulson: Pro.
Natl. Acad. Sci. , USA, 74, 5463 (1977) 4. Favalora, J. ,R. Treisman and R. Kamen; Me-
htods Enzνmol., 65, 718 (1980)
5. Hinnebusch, A. G. and G. R. Fink; J. o{ Biol.
Chem. ,258,5238 (1983)
6. Struhl, K.; Proc. Natl. Acad. Sci., USA, 78, 4461 (1981)
7. Donahue. T. F., P. J. Farabaugh and G. R. Fink Gene, 18,47(1982)
8. Donahue, T. F. , R. S. Daves, G. Lucchini and G.
R. Fink; Cell, 32, 89 (1983)
9. Zalkin, H. and C. Yanofsky; J.o{ Biol. Chem.’ 257, 1491 (1982)
10. Andreadis, A., Y. P. Hsu, G. B. Kohlhaw and P.
Schimmel; Cell, 31, 319(1982)
11. Dobson, M. J., M. F. Tuite, N. A. Robert, A. J.
Kingsman and S. M. Kingsman; Nuc/eic acids Res., 10,2525 (1982)
12. Kaput, J. , S. Goltz and G. Blobel; J.o{ Biol.
Chem. ,257, 15054 (1982)
13. Taussing, R. and M. Cerlson; Nuc/eic acid Res., 11, 1943 (1983)
14. Harashima, S.,R. S. Si에1U, A. Toh-e and Y.Os- 본 연구는 1983 년 9 월부터 1984 년 8 월까지 국 hima.: Gene, 16,335 (1981)
비 연구교수 (운교부)로 파견되어 오샤카대학 공학부 15. Struhl, K.; Proc. Natl. Acad. Sci. USA, 78, 발효공학과에서 공동으로 이루어진 결과의 일부분 4461 (1981)
임 을 밝혀 둔다 16. Donahue, T. F. , P. J. Farabaugh and G. R. Fink 연구바플 지급해 준 운교부와 본 연구수행에 지 Gene, 18,47 (1982)
도와 편말을 하여주신 *嗚泰治교수. 高田信男 조 17. 入江新司 : 碩士좋位쏠文, (大限大좋工좋部 훌 교수, 原島 俊 조교에게 감사를 드리는 바이다. 홈工좋科 (1983) )