재조합 대장균의 고농도 배양과 유도조건 최적화를 통한 Bacillus 유래 esterase의 생산
강승훈†, 민병혁†, 최홍열, 김동일*
인하대학교생명공학과
Received: March 16, 2017 / Revised: May 8, 2017 / Accepted: May 17, 2017
서 론
Esterase, peroxidase, α-amylase를비롯한효소자원은 식품, 의약품및섬유산업에이르기까지광범위한분야에서 사용되고있다[1, 2]. Esterase의주요기능은생체내에서다 양한지방질의가수분해에관여하지만, 효소반응조건에따 라역반응인에스터합성반응과트랜스에스터반응을촉매 하는특성을가지고있다[3, 4]. 산업적관점에서의 esterase 는주로아미노산합성, steroid 전환, ester 전환및합성공 정등에서효소광학분활법(enzymatic optical resolution)에의해
D 형과 L 형이혼합된물질을 D 형또는 L 형으로분리하 는데이용된다[5, 6]. 현재까지다양한종류의 esterase가발 견되었는데, 주로미생물에서유래한 esterase가기질에대 한특이성, 내알칼리성, 유기용매내성, 내열성그리고광학 선택성(enantioselectivity) 등의특성을갖고있는것으로밝
혀져다양한산업분야에서널리이용되고있다[7, 8]. 특히
L-ofloxacin인 levofloxacin과D-ofloxacin이 1:1로섞여있는 광학이성질체에서 esterase가갖는광학선택성촉매반응
을 통해 D-ofloxacin으로부터 항균활성이 월등히 뛰어난
levofloxacin을생전환하기위한시도가이루어지고있다[9].
대장균은유전자재조합단백질생산에가장널리이용되 는균주로서인터페론, 인터루킨, CSF (colony stimulation factor), 성장호르몬, IGF (insulin like growth factor) 등의
생산에사용되고있다[10]. 뿐만아니라대장균은산업적으
로유용한효소의발현을위해서도사용되고있으며, 재조합 Optimization of Induction Conditions for Bacillus-derived Esterase Production by High-cell Density Fermentation of Recombinant Escherichia coli
Seung-Hoon Kang†, Byung-Hyuk Min†, Hong-Yeol Choi, and Dong-Il Kim*
Department of Biological Engineering, Inha University, Incheon 22212, Republic of Korea
To increase the efficiency of esterase production by Bacillus, high cell-density culture of recombinant Escherichia coli through fed batch fermentation was tested. Cells were cultured to OD600 of 76 (35.8 g/l DCW) with dissolved oxygen level controlled to least above 30% air saturation by supplying pure oxygen.
Cells were cultured to an OD600 of 90 (42.4 g/l DCW) with glucose feeding controlled to at least 1 g/l. How- ever, the cells reached stationary phase at the late stage of culture, despite glucose being supplied. Cells were cultured to an OD600 of 185 (87.3 g/l DCW) by supplying additional medium with fortified yeast extract.
To increase the productivity of the recombinant protein, cell growth and esterase productivity based on induction time were evaluated. Late exponential phase induction for esterase production in fed batch fer- mentation resulted in maximum optical density OD600 of 190 (89 g/l DCW) and maximum esterase activity of 1745 U/l, corresponding to a 5.8-fold enhancement in esterase production, compared to the early exponen- tial phase induction. In this study, we established fermentation methods for achieving maximum produc- tion of Bacillus-derived esterase by optimizing IPTG induction time in high-cell density culture by supplying pure oxygen and a nitrogen source.
Keywords: Esterase, high cell density fermentation, isopropyl β-D-1-thiogalactopyranoside induction, Escherichia coli
*Corresponding author
Tel: +82-32-860-7515, Fax: +82-32-872-4046 E-mail: [email protected]
†These authors contributed equally to this work as the first author.
© 2017, The Korean Society for Microbiology and Biotechnology
효소가발현된대장균세포를고정화하여생물학적촉매로 서이용하기위한연구가이루어지고있다[11]. 이러한고정 화세포의이용은재조합효소의분리정제과정이없이도해 당효소의사용이가능하기때문에비용측면에서장점을가 지고있으며, 고정화과정을통해단위공간당높은세포농
도의유지가가능하여생전환효율을높일수있다[12].
대장균에서단백질생산성을향상시키기위해서는단위시 간, 단위세포당재조합단백질의생산성향상과함께세포 의고농도배양을수행하여목적단백질이발현된균체를대
량으로확보하는것이유리하다[13]. 대장균의고농도배양
은재조합단백질의생산성높일수있을뿐만아니라폐기
물질과배양비용을감소시킬수있는장점이있다[14]. 대
장균고농도배양을위해서는유가식배양이일반적이며기
질저해, 산소의고갈, acetate의생성에따른대장균의성장
저하뿐만아니라재조합단백질의생산성저하를극복할수 있는배양방법의확립이중요하다[15, 16].
유가식배양에서추가배지의공급속도는배양과정의주 요공정변수인 pH, DO, 포도당농도등의변화에기초하여 결정되며, 공급방식에따라 constant rate feeding, gradual feeding 그리고 exponential feeding 방법들이적용된다[17,
18]. 추가배지에대한공급속도조절을통해대장균의비
성장속도를제어할수있으며, 비성장속도에따라서재조
합단백질의생산성에영향을미친다[19]. 따라서세포의성
장과단백질발현량을모두만족할수있는최적화된추가 배지공급방식의선택이중요하다.
따라서, 본 연구에서는 유가식 배양에서 Bacillus 유래
esterase를발현할수있는재조합대장균의고농도배양을
위해, 용존산소농도및포도당공급조절과추가배지조성 의강화를시도하였다. 추가적으로단백질발현시기의조절 에따른 esterase 생산성평가를통해최적 induction 시기
를확인하여최대 esterase 생산성을갖는배양조건을확립
하고자하였다.
재료 및 방법
사용 균주 및 배지
본연구에는 E. coli BL21/pET24b를기반으로 Bacillus pumilus 유래의 esterase (DQ339137.1)가 cloning된균주 를보령제약중앙연구소로부터분양받아사용하였다. 종배 양발효는 LB 배지를사용하였으며, 본배양발효를위한기 본 배지(substrate medium) 조성은 (NH4)2HPO4 2 g/l, KH2PO4 6.75 g/l, Citric acid 0.85 g/l, MgSO4·7H2O 0.7 g/l, Glucose 10 g/l, Yeast extract 2 g/l, trace metal solution 5 ml (FeSO4· 7H2O 10 g/l, ZnSO4· 7H2O 2.25 g/l, CuSO4· 5H2O 1 g/l, MnSO4·5H2O 0.5 g/l, Na2B4O7·10H2O 0.23 g/l,
CaCl2· 2H2O 2 g/l)이며, 추가 기질 공급 배지는 glucose 500 g/l, MgSO4· 7H2O 15 g/l, yeast extract 2 g/l를사용하
였다. 질소원의강화를목적으로사용된강화배지(fortified
medium)의조성은기본배지조성을기반으로 yeast extract 의농도를 2 g/l에서 20 g/l로상향조절하였으며, 추가기질 공급 배지는 glucose 500 g/l, MgSO4· 7H2O 15 g/l, yeast extract 20 g/l를사용하였다.
발효기 운전 조건
1차종배양을위해 kanamycin 40 mg/l가포함된 LB 배지 5 ml에 1%(v/v)로접종후 12시간동안 250 rpm, 37℃에서 진탕배양하였다. 2차종배양은 50 ml의배지에접종하여 1차종배양과동일조건으로진행하였다. 본배양을위해 5-L fermentor (Kobiotech, Korea)에 2 L의 working volume으 로배지를준비한후 2차종배양액을 5% (v/v)로접종하였 다. 본배양을위한발효기운전조건은 37℃, 통기속도 4 vvm,
교반속도 600 rpm을유지하였으며, 산소농도가감소하는시
점에서교반속도를 800 rpm까지단계적으로증가시킨후
100% 산소를간헐적으로공급하여 30% 이상의산소농도가
유지되도록설정하였다. 유가식배양을위해 30분마다포도
당농도를확인하고포도당농도가 1 g/l 이하를유지하도록
feeding 배지의공급속도를조절하였다. 본배양중 pH의조 절은암모니아수(28%)를사용하였고, 배양중발생하는 foam 의제거를위해 antifoam A (Sigma, USA)와 Adecanol LG 109 (Asahi Denka, Japan)를첨가하였다. Esterase의발현을 위해 IPTG (isopropyl-β-D-thiogalactopyranoside)를 1 mM로 첨가하여 induction하였다.
세포 성장 및 glucose 농도 측정
균체량의측정은 spectrophotometer (Agilent 8453, Agilent Technologies, USA)을사용하여 OD600에서의흡광도를측 정하였고, 흡광도와건조균체량의표준곡선에의해결정하 였다. 흡광도와건조균체량의표준곡선을구하기위해대장 균배양액을 LB 배지를사용하여단계적으로희석한후각 각의흡광도를측정하였고, 8,000 ×g에서 15분간원심분리 후침전된균체를 80℃ dry oven에서 24시간건조시켜건조 중량을측정하였다. 배양액내의 glucose는 glucose analyzer (YSI2000 select, USA)를사용하여측정하였다.
세포 내 esterase 활성 측정
Induction에 의해 대장균 세포 내에서 발현된 esterase activity 측정을 위해 세포를 OD600 5로 희석한 후 그 중 200μl를취하여원심분리한후상등액을제거하고, ethanol 과 50 mM sodium phosphate buffer (pH 6.8)가각각 4:95 의비율로혼합된 reaction buffer 500 μl를첨가하였다. 효
소 반응을 위한 기질로 acetonitrile에 용해된 10 mM p- nitrophenyl caprylate (PNPC) 10 μl를첨가하여반응을개
시하고상온에서 1분간반응후 405 nm에서흡광도를측정
하였다. 효소의활성측정은효소반응을통해생성된산물인 pNP (p-nitrophenol)의표준곡선을통해, 1분당 1 μmole의 pNP를생성하는효소활성을 1 unit으로정의하였다. 발효
부피당 esterase의생산성은시간별로측정된세포중량당
esterase의활성값에해당시점의세포농도를반영하여산
출하였다.
결과 및 고찰
용존산소 농도 조절에 의한 세포 성장
호기성균주의발효시배지내의용존산소는미생물의산 소요구량을만족시키기위한중요인자중하나이다. 산소 는수용액상에서용해도가낮기때문에미생물의산소요구 량을만족시킬수있도록통기속도와교반속도를증가시키 거나, 추가적인순수산소(pure oxygen)의공급또는산소의 분압을조절하여고압환경을유지함으로써용존산소를조절
한다[20, 21]. 산소소비속도는결국세포생장속도와비례
하기때문에산소의부족은미생물대사를저해하고성장속 도를저하시키는원인으로작용할뿐만아니라, 재조합단백
질의발현에도중요한인자로작용한다[22].
세포농도가증가함에따라나타나는산소고갈의문제를
해결하기위해교반속도 600 rpm, 통기속도 4 vvm의발효
조건을기반으로추가적으로순수산소를 solenoid valve와 연결하여배양액중용존산소농도를 air saturated condition
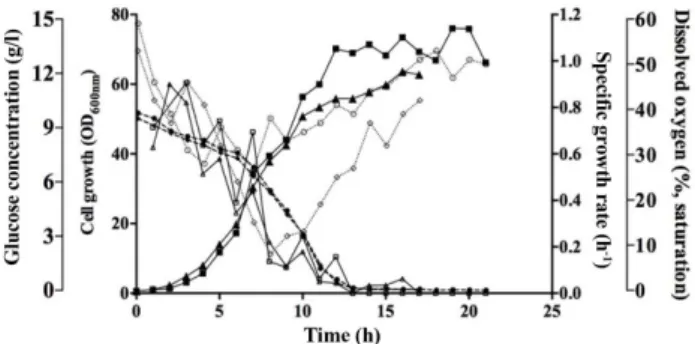
의 30% 이상으로유지하여세포성장을비교하였다(Fig. 1).
실험결과순수산소를공급하지않은경우배양 17시간 째산소부족이관찰되었으며, 이때최종 OD600는 63 (29.7 g/l DCW)로확인되었다. 반면용존산소농도가 30% 이상이유 지되도록순수산소를공급하는경우최종 OD600는 76 (35.8
g/l DCW)으로세포농도가증가함을확인하였고, 배양시간
은 20시간이상유지되었다. 그러나순수산소공급에의한 세포성장은유의한증가율을보이지않았는데, 이는당을 비롯한영양원의공급에제한이발생하기때문인것으로판 단되었다.
포도당과 yeast extract 공급에 의한 세포 성장
대장균고농도배양을위해서추가적인당의공급과공급
속도는세포성장에있어서매우중요한인자가된다[23]. 당
의공급이세포의생장을따라가지못하면영양원의부족으 로세포생장과단백질합성이원활하게이루어질수없고, 과량의당이공급되면대사과정의부산물로서 acetic acid 생 산이증가하여, 세포생장과단백질생산에부정적인결과를 초래하므로, E. coli 고농도배양을위해서는당공급속도의 조절이필수적이다[24]. 따라서배양중포도당농도를확인
하여포도당농도가 1 g/l 이하가되도록공급속도를조절하
여배양을진행하였다. 포도당공급조절을통해얻은최대 OD600은 90 (42.4 g/l DCW)이었으며이전실험과비교시활 발히생장이이루어지는초기세포생장을크게향상시킬수
있었다(Fig. 2). 그러나배양후반인 10시간이후에는세포
의생장이정체되는현상을확인할수있었다.
배양후반세포생장의정체를극복하기위해공급배지의
Fig. 1. Time course changes of cell growth, specific growth rate, glucose concentration and dissolved oxygen with and without pure oxygen supply during fermentation. ▲, cell growth profiles without pure oxygen supply; ■, cell growth pro- files with pure oxygen supply; △, specific growth rate without pure oxygen supply; □, specific growth rate with pure oxygen supply; ◆, glucose concentration without pure oxygen supply;
●, glucose concentration with pure oxygen supply; ◊, dissolved oxygen without pure oxygen supply; ○, dissolved oxygen with pure oxygen supply.
Fig. 2. Time course changes of cell growth, specific growth rate and glucose concentration with substrate medium or fortified medium as an additional feeding media. ▲, cell growth profiles with substrate medium; ■, cell growth profiles with fortified medium; △, specific growth rate with substrate medium; □, specific growth rate with fortified medium; ◆, glu- cose concentration with substrate medium; ●, glucose concen- tration with fortified medium.
yeast extract를강화하여배양을진행하였고, 공급배지로사 용된 yeast extract 성분이배양후반에필요한당이외의영 양원으로사용되어세포성장을극대화시킬수있기를기 대하였다. 다른연구의경우에도 yeast extract의첨가를통 한질소원의공급이세포의생장을향상시키고, 외래단백질
생산성을높였다는결과가보고된바있다[25]. 따라서배양
후반의 생장 저하를 극복하기 위해 본배양 배지의 yeast
extract 농도를 2 g/l에서 20 g/l로강화하였고, feeding media에 도 20 g/l의 yeast extract를첨가하여배양을 진행하였다. Yeast extract의강화에의해확인된최대세포성장은 OD600
185 (87.3 g/l DCW)로서포도당농도의조절을통해얻은세 포성장에비해세포농도가 2배이상증가할뿐만아니라지 수성장기의비성장속도역시도 0.759 h-1에서 1.146 h-1로증 가함을확인할수있었다(Fig. 2).
IPTC 첨가 시기에 의한 esterase 생산성
대장균배양에서 IPTG에의한목적단백질생합성의유
도는세포안에제한적으로존재하는자원을세포의기능유 지를위한단백질의합성과목적단백질의합성에동시에사 용함으로써대사과정에부하를유발하여세포의생장을저
해하는효과를나타낸다[26]. 따라서목적단백질의생산을
위한 induction 시기는세포의성장시기와농도를고려하
여결정되어야하며, 최대의생산성을확보할수있는적절 한시기에이루어져야한다. 본연구에서는앞서확립된고 농도배양이가능한유가식배양조건을기반으로최적의
IPTG induction 시기를결정하기위해세포의지수성장기
초기, 중기, 후기에해당하는 3 h, 9 h, 12 h에 IPTG induction
을진행하여발현유도시기에따른세포의성장과 esterase
생산성을확인하였다.
세포의배양조건은앞서확립된고농도세포배양을위 한유가식배양조건을기초로진행하였고, 1 mM의 IPTG 를첨가하여 induction을시도하였다.
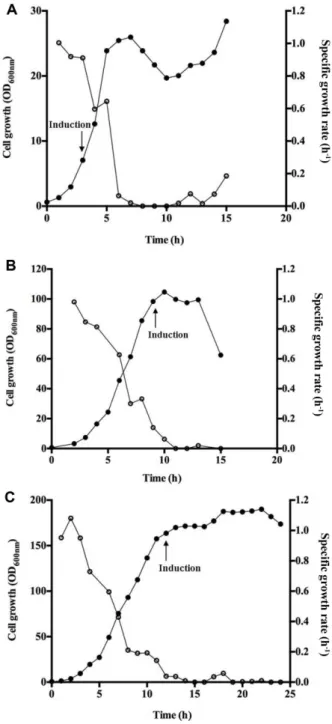
지수성장기초반인 OD600 7 (3.2 g/l DCW)에서 induction을 유도한경우최대 OD600 28 (13.2 g/l DCW)로가장낮은세 포성장을나타내었다(Fig. 3A). 세포의비성장속도는접종 후 induction 직전까지 0.911 h-1 이상을 유지하였으나, induction 후 0.596 h-1으로감소하고 induction 3시간이후 부터는세포의성장이정체되는것으로확인되었다. 지수성 장기중반인 OD600 98 (46.2 g/l DCW)에 induction을유도 한경우 IPTG 첨가후세포의비성장속도가 0.627 h-1에서 0.301 h-1로감소하였고, induction 4시간후에는세포의성 장이 정체되었으며, 최대 세포성장은 OD600 104 (49 g/l DCW)로확인되었다(Fig. 3B). 특히 induction 유도후 6시
간째에는급격한세포사멸이이루어져 induction이세포의
성장에심각한영향을미치는것으로확인되었다. 지수성장
기후반인 OD600 163 (77 g/l DCW)에서 induction을유도한 경우 induction 전후의비성장속도는각각 0.191 h-1, 0.193 h-1
으로 induction에도불구하고세포의성장속도에는큰영향
을미치지않았으며, induction 후 13시간까지점진적으로
Fig. 3. Time course changes of cell growth and specific growth rate with induction during the early exponential phase (A), mid exponential phase (B) and late exponential phase (C). ●, cell growth profiles; ○, specific growth rate;
arrows indicate the induction times using IPTG.
세포가성장하여최대 OD600 190 (89 g/l DCW)까지증가하 였다(Fig. 3C).
Induction 이후시간에따른 esterase의상대활성변화를 확인한결과, 지수성장기초기에 induction을유도한경우 4시 간에 26.8 mU/g DCW로 esterase의단위세포당활성이증 가하였으나, 배양시간이길어질수록점차감소하는경향을 보였다(Fig. 4A). 그러나배양후반 induction에의해정체되 었던세포의성장이다시개시되었음에도불구하고, 단위세
포당 esterase 활성은오히려감소하는것을확인할수있
었다(Fig. 3A, Fig. 4A). 또한지수성장기초기에 induction을
유도한경우단위세포당 esterase의활성이가장높았으나,
부피당활성측면에서는가장낮은것으로확인되었다. 지수
성장기중반에 induction을유도한경우에는지수성장기초
반에 induction을유도한경우에비해세포농도측면에서
는약 3.7배정도높게 성장하였으나, 단위 세포당활성은 15.5 mU/g DCW로오히려낮아졌다(Fig. 4A). 그러나부피 당활성측면에서는 2.03배의생산성증가가확인되었다(Fig.
4B). 지수성장기후반에 induction을유도한경우유도후 7시간까지단위세포당 esterase의활성이증가하였으며, 최
고상대활성은 19.7 mU/g DCW로확인되었다. 단위세포
당상대활성측면에서는지수성장기초반에 induction을 유도한경우와비교하여약 1.4배낮은수치를보였으나, 부 피당생산성측면에서는지수성장기초반대비 5.8배, 지수 성장기중반대비 2.8배의생산성증가가확인되었다. 선행 연구에서회분식배양에서의대장균발현시스템을사용한
재조합 EcoRI 단백질의발현유도에있어서지수성장의중
간시점에서 1 mM의 IPTG를사용한조건이최적생산성을
나타내는것으로확인하였다[27]. 그러나본연구에서적용 한유가식배양조건에서는지수성장기후반에 induction을 하는경우최대생산성을갖는것으로확인되었고, 이는유 가식배양을통한고농도배양에있어서균체량의증가가재 조합단백질의생산성에중요한영향을미침을시사한다.
이상의결과를통해 Bacillus 유래 esterase의생산을위 한대장균배양에있어서, 최고 OD600 185 (87.3 g/l DCW)까 지고농도배양이가능한유가식배양조건을확립하였고, 단백질발현유도시기에따른 esterase의생산성을확인하 여, 지수성장기후반에단백질발현을유도하는경우최대 생산성을나타냄을확인하였다.
요 약
본연구에서는 Bacillus 유래 esterase를생산할수있는 재조합대장균을사용하여유가식배양을이용한고농도균
체배양을통해 esterase 생산성을극대화하고자하였다. 유
가식배양중순수산소의공급을통해용존산소를 30% 이 상유지한경우와포도당농도를 1 g/l 이상유지한경우각 각 OD600 76 (35.8 g/l DCW)과 OD600 90 (42.4 g/l DCW) 까지균체량을증가시킬수있었다. 포도당의공급에도불구 하고배양후반에세포의성장이정체되는현상을극복하기
위해 yeast extract가강화된추가배지의공급을시도하였
으며, 그결과 OD600 185 (87.3 g/l DCW)까지고농도균체 배양이가능함을확인하였다. 단백질생산수율의향상을위 해성장시기에따라 induction에의한세포성장과 esterase 생산성을평가하였고, 그결과대수성장기후반에 induction 을 유도한 경우 세포 성장 측면에서는 최대 OD600 190
(89 g/l DCW)까지고농도균체배양이가능함을확인하였다.
Esterase 생산성측면에서는대수성장기초반에 induction 을유도한경우에비해최대 5.8배생산성이증가됨을확인 Fig. 4. (A) Time course profiles of relative esterase activity
after induction during early exponential phase (gray), mid exponential phase (dark gray) and late exponential phase (black) and (B) Volumetric productivity of recombinant esterase production depending on induction time.
할수있었다. 따라서본연구를통해순수산소와질소원의 공급을통해확립된대장균고밀도배양방법을기초로 IPTG 유도시간을최적화함으로써 Bacillus 유래 esterase의최대 생산성을확보할수있는배양방법을확립하였다.
Acknowledgments
This work was supported by the National Research Foundation of Korea (NRF) grant (No. NRF-2013M3A9B6075887).
References
1. Adrio JL, Demain AL. 2014. Microbial enzymes : tools for bio- technological process. Biomolecules 4: 117-139.
2. Souza PM, Magalhaes PO. 2010. Application of microbial α- amylase in industry – A review. Braz. J. Micribiol. 41: 850-861.
3. Lowe ME. 2002. The triglyceride lipases of the pancreas. J. Lipid Res. 43: 2007-2016.
4. Yee LN, Akoh CC, Phillips RS. 1995. Terpene ester synthesis by lipase catalyzed transesterification. Biotechnol. Lett. 17: 67-70.
5. Patel RN. 2013. Biocatalytic synthesis of chiral alcohols and amino acids for development of pharmaceuticals. Biomole- cules 3: 741-777.
6. Elmi F, Lee HT, Huang JY, Hsieh YC, Wang YL, Chen YJ, et al.
2005. Stereoselective esterase from Pseudomonas putida IFO12996 reveals alpha/beta hydrolase folds for D-beta-acet- ylthioisobutyric acid synthesis. J. Bacteriol. 187: 8470-8476.
7. Ghati A, Paul G. 2015. Purification and characterization of a thermo-halophilic, alkali-stable and extremely benzene toler- ant esterase from a thermo-halo tolerant Bacillus cereus strain AGP-03, isolated from ‘Bakreshwar’ hot spring, India. Process Biochem. 50: 771-781.
8. Kaiser P, Raina C, Parshad R, Johri S, Verma V, Andrabi KI, et al.
2006. A novel esterase form Bacillus subtillis (RRL 1789) : purifi- cation and characterization of the enzyme. Protein. Expr. Purif.
45: 262-268.
9. Kim HK, Na HS, Park MS, Oh TK, Lee TS. 2004. Occurrence of ofloxacin ester-hydrolyzing esterase from Bacillus niacin EM001. J.
Mol. Caltal B: Enzym. 27: 237-241.
10. Baeshen MN, Al-Hejin AM, Bora RS, Ahmed MM, Ramadan HA, Saini KS, et al. 2015. Production of biopharmaceuticals in E. coli : Current scenario and future perspective. J. Microbiol. Biotech- nol. 25: 953-962.
11. Djokic L, Spasic J, Jeremic S, Vasilievic B, Prodanovic O, Prodanovic R, et al. 2015. Immobilization of Escherichia coli cells expressing 4-oxalocrotonate tautomerase for improved bio- transformation of β-nitrostyrene. Bioprocess Biosyst. Eng. 38:
2389-2395.
12. Zajkoska P, Rebros M, Rosenberg M. 2013. Biocatalysis with immobilized E. coli. Appl. Microbiol. Biotechnol. 97: 1441-1455.
13. Shiloach J, Fass R. 2005. Growing E. coli to high cell density – a historical perspective on method development. Biotechnol.
Adv. 23: 345-357.
14. Nakagawa S, Oda H, Anazawa H. 1995. High cell density culti- vation and high recombinant protein production of Esche- richia coli strain expressing uricase. Biosci. Biotech. Biochem. 59:
2263-2267.
15. Lee SY. 1996. High cell density culture of Escherichia coli. Trends Biotechnol. 14: 98-105.
16. Luli GW, Strohl WR. 1990. Comparison of growth, acetate pro- duction, and acetate inhibition of Escherichia coli strains in batch and fed-batch fermentations. Appl. Environ. Microbiol.
56: 1004-1011.
17. Castan A, Enfors SO. 2000. Characteristics of DO controlled fed batch culture of Escherichia coli. Bioprocess Eng. 22: 509-515.
18. Yazdani SS, Shakri AR, Chitnis CE. 2004. A high cell density fer- mentation strategy to produce recombinant malarial antigen in E. coli. Biotechnol. Lett. 26: 1891-1895.
19. Kovarova-Kovar K, Egli T. 1998. Growth kinetics of suspended microbial cells : From single-substrate-controlled growth to mixed-substrate kinetics. Microbiol. Mol. Biol. Rev. 62: 646-666.
20. Belo I, Mota M. 1998. Batch and fed batch cultures of E. coli TB1 at different oxygen transfer rate. Bioprocess Eng. 18: 451-455.
21. Liu YC, Chang WM, Lee CY. 1999. Effect of oxygen enrichment aeration on penicillin G acylase production in high cell density culture of recombinant E. coli. Bioprocess Eng. 21: 227-230.
22. Ukkonen K, Veijola J, Vasala A, Neubauer P. 2013. Effect of cul- ture medium, host strain and oxygen transfer on recombinant Fab antibody fragment yield and leakage to medium in shaken E. coli cultures. Microb. Cell Fact. 12: 73.
23. Eiteman MA, Altman E. 2006. Overcoming acetate in Esche- richia coli recombinant protein fermentations. Trends Biotech- nol. 24: 530-536.
24. Suarez DC, Kilikian BV. 2000. Acetic acid accumulation in aero- bic growth of recombinant Escherichia coli. Process Biochem.
35: 1051-1055.
25. Kweon DH, Han NS, Park KM, Seo JH. 2001. Overproduction of Phytolacca insularis protein in batch and fed-batch culture of recombinant Escherichia coli. Process Biochem. 36: 537-542.
26. Kilikian BV, Suarez ID, Liria CW, Gombert AK. 2000. Process strategies to improve heterologous protein production in Escherichia coli under lactose or IPTG induction. Process Bio- chem. 35: 1019-1025.
27. Yildir C. Onsan ZI, Kirdar B. 1998. Optimization of starting time and period of induction and inducer concentration in the pro- duction of the restriction enzyme EcoRI from recombinant Escherichia coli 294. Turk J. Chem. 22: 221-226.