유전자가위 사용설명서와 이를 이용한 선천성 면역결핍환자 대상 유전자 치료
서론
1970년에 ‘제한효소’가 처음 발견된 이후 8년 만에 3명의 과학자들이 (Werner Arber, Daniel Nathan, & Hamilton Smith) 노벨 생리의학상을 공동 수상하였다. 지금은 많은 연구실에서 흔히 사용하는 효소이지만, 46년전인 그 당시 에는 획기적으로 새로운 분자생물학적 도구가 이 분야를 엄 청난 속도로 발전 시키는 밑거름이 되었고, 현재에도 여전 히 우리는 이 분자생물학적 도구를 이용하여 모든 생명체의 유전자를 우리가 원하는 대로 자르고 붙이면서 다양한 연구 를 수행하고 있다. 이 발견 이후, 37년 만인 2007년에 미국 의 바이오벤처 기업인 ‘SANGAMO’ 에서 또 다시 분자생물 학에 커다란 영향을 미치는 기술을 발표하게 되는데, 그것은 zinc finger nuclease (ZFN)라는 기존에 없었던 새로운 방 법의 유전자 조작을 자유자재로 할 수 있는 일명 ‘유전자가
위’ 를 개발해 내었다. 물론 제한효소도 유전자를 자르는 가 위 역할을 하지만, 그 대상이 극히 제한되어 있고, 주로 작 은 사이즈의 DNA (5,000-10,000 bp)를 다룬다면, ZFN 는 게놈전체를 대상으로 우리가 원하는 특정 부위만을 절단 하는 기능이 있어 기존의 제한효소와는 다른 신기술인 것이 다. 그 후 2011년에 Dr. Daniel F. Voytas가 transcription activator-like effector nuclease (TALEN)를 그리고, 2013년에 Dr. George M. Church가 clustered regularly- interspaced short palindromic repeats (CRISPR/Cas9) system과 같은 제2, 제3의 유전자가위를 세상에 발표하면 서 유전자가위의 다양한 적용 범위와 그 중요성이 계속 부각 되고 있다. 특히 CRISPR/Cas9 system은 지금까지 나온 유 전자가위중 가장 성능이 뛰어나고, 널리 알려진 지 3년 밖에 되지 않았지만, 이미 전세계의 많은 과학자들이 다양한 방법 과 목적으로 사용하고 있다. 이러한 유전자가위의 사용 범 최 의 묵
Laboratory of Host Defense, NIAID, NIH, USA [email protected]
위, 사용시 꼭 알아두어야 할 중요한 요인들(CRISPR/Cas9 system을 주로 언급할 것임), 마지막으로 임상에서 실제 활 용방법에 대해 간략하게 소개해 보려고 한다.
유전자가위의 사용범위
유전자가위 (ZFN, TALEN, CRISPR/Cas9)가 나오기 전 인 2006년도까지만해도, 과학자들은 유전자 조작(Knock- out, knock-in, transgenic animal 등)을 유전자가위 없 이도 꾸준히 해오고 있었다. 물론 엄청난 시간과 노력이 뒤 따르는 고행의 연속이었다. 예를 들어 유전자가위 없이 knock-out mouse, 또는 knock-in mouse를 만들어 내는 데, 보통 1년에서 2년의 시간이 필요했고, 비용도 $10,000-
$30,000이상 소요되었다. 지금은 많은 연구실에서 유전자 가위를 이용해서, 5-6개월이면, 같은 모델의 생쥐를 만들 고, 비용도 25% 이하로 드는 것이 일반적이다. 동물모델뿐 만이 아니라, 유전자가위는 모든 종류의 배양세포들의 유전 자 조작을 빠르게는 한달 안에 해낼 수 있다. 10년 전만 해 도 유전자 조작을 한 배양세포주를 모델로 해서 저명한 국 제저널(임펙트10)에 낼 수 있었던 것과는 다르게, 현재 그와 비슷한 저널의 일부로 사용되어 유전자 조작은 더 이상 그 자체가 목적이 될 수 없는 일반화된 실험 방법이 되었다. 가 장 일반적이고, 그 효과도 가장 좋은 유전자가위의 사용분야 는 표적유전자 knock-out이다. 그 다음으로 많이 사용되는 분야는 원하는 유전자를 원하는 세포에 과발현 시키는 것인 데, 이때 safe harbor라고 하는 게놈상의 특정 부위에 원하 는 유전자카세트(mini-gene; left homologous arm, gene to express like GFP, positive selection marker like puromycin resistant gene, right homologous arm and negative selection marker like HSV-tk gene so called suicide gene)를 삽입하는 방법을 쓰는데, 인간의 경우 염색 체 19번에 있는 AAVS1 과 생쥐의 경우는 염색체 6번에 있 는 ROSA26이 각각의 좋은 예이다. 이와 같은 세포주 제작 을 위해서는 safe harbor부위를 자를 수 있는 유전자가위와 위에서 언급한 mini-gene을 함께 배양세포나 생쥐의 수정 란에 넣으면 제작이 가능하다. Safe harbor에 원하는 카세 트를 삽입하면, 어떤 조직의 세포든 어떤 상태에 있든 원하 는 유전자를 항상 안정적으로 발현하게 할 수 있기 때문에,
최근 이 방법을 이용해서 발표되는 저널이 증가하는 추세 에 있다. 또 다른 사용범위는 유전자가위에서 가위역학을 하 는 nuclease를 없애고 대신 유전자발현을 도와 주는 활성제 를 가위가 있던 부위에 넣어 주면, 우리가 원하는 유전자만 을 더욱 활성화 시킬 수 있다. 가장 최근에 발표된 저널에 의 하면, acetyltransferase (p300 core)의 핵심부위만을 Cas9 의 가위부분과 바꾸어준 결과, 정확하게 표적으로 하는 유전 자만을 10에서 100배까지의 과발현을 유도해 낼 수가 있었 다[1]. 마지막으로 소개할 유전자가위의 사용범위는 single strand oligo donor(SSOD)를 이용한 knock-in이다. 단일 유전자의 단일 염기가 변형되어 일어나는 유전병(X-SCID, X-CGD, LAD etc.)을 치료할 수 있는 이 방법은 가장 어렵 고 성공확률도 낮은 방법이다. 하지만 많은 질병의 가장 이 상적인 치료방법이 될 수 있기 때문에 지속적으로 발전시켜 나가야 할 영역이다.
유전자가위 사용 시 고려사항
원하는 유전자를 knock-out하는 방법에 있어서는 특별 히 숙지해야 할 사항이 없으나 몇 가지를 말하자면, 가능한
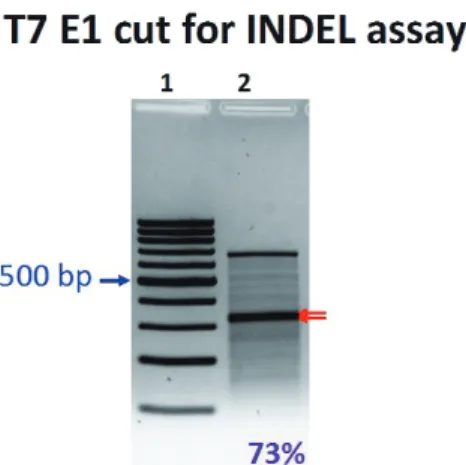
그림 1. INDEL assay in K562 cells transfected with DNA
유전자의 앞쪽인 exon1 이나 exon2를 표적부위로 삼고 실 험을 설계 하는 것이 유리 하다. 그래야 완전한 knock-out 을 효율적으로 유도해 낼 수 있다. 필자는 주로 crispr.mit.
edu 사이트에서 CRISPR/Cas9 system을 디자인 하고 모 든 정보를 다운로드 해놓는다. 적어도 3개의 추천 gRNA 를 만들어서 insertion or deletion (INDEL)assay [그 림1]를 통해서 가장 효과적인 gRNA를 선택하여 본 실험 에 사용하는 것이 안전하다. CRISPR/Cas9 system에 필 요한 모든 종류의 plasmid DNA들은 www.addgene.org 에서 쉽고 저렴하게 얻을 수가 있다. 보통 발현 벡터를 제 작하는데 이틀 정도가 소요되고, 최초 디자인에서 최종 INDEL assay까지 일주일이면 가능하다. 가장 쉽게 그리 고 효과적으로 사용할 수 있는 것 중 한가지를 소개 하자 면, plentyCRISPR V2라는 plasmid DNA backbone을 권 하고 싶다[그림2]. 원하는 gRNA를 cloning site인 BsmBI 에 넣어주면, trRNA가 gRNA와 연결이 되고, SpCas9단백 질은 EF1-alpha promoter에 의해 항시 많은 양이 발현을
하고 FLAG tag이 있어서, 발현 정도를 확인 할 수 있다. 또 한 puromycin resistant cDNA가 self-cleavage peptide (P2A)로 SpCas9에 연결 되어있기 때문에 transfection이나 transduction후에 puromycin-selection 을 하면, 살아남 는 세포들은 모두가 knock-out이 되어있기 때문에 single cell cloning을 따로 하지 않아도 된다. 또한 이 모든 구조 가 HIV lenti viral vector안에 있기 때문에, 바이러스를 만 들어서 사용하면 더욱 효과적으로 어떤 세포든 원하는 표적 유전자를 knock-out시킬 수가 있다. 일반적으로 knock- in rate은 knock-out rate의 1/100 또는 1/50 정도에 그친 다. 그 이유는 CRISPR/Cas9 system으로 원하는 부분을 자 르면 세포 안에 있는 DNA ligase IV가 곧바로 잘라진 가닥 의 DNA를 다시 복원하기 때문이다. 물론 이때 잘라진 곳을 기준으로 몇 개의 염기가 없어지거나, 더해지면서 결국에는 형질전환의 돌연변이를 유발하고, 이에 따라 표적 유전자를 knock-out하는 효과를 가져온다. 이때 DNA ligase IV의 복원능력은 생각보다 빨라서 유전자가위가 표적부위를 자 그림 2. Plasmid DNA map for CRISPR/Cas9 system
른 후 곧바로 homologous recombination (HR)을 유발할 SSOD가 옆에 없으면, knock-in은 발생하지 않는다. 그 이 유는, 복원 부분이 이미 변형이 되어 있어서 다시 CRISPR/
Cas9 system에 의해서 잘라질 수가 없기 때문이다. 그렇 기 때문에 SSOD를 이용한 knock-in 은 recombination 이 일어나는 순간의 ‘타이밍’이 가장 중요한 요인이다. 또한 safe harbor에 mini-gene 을 이용한 knock-in의 경우, positive selection 과 negative selection 을 통해서 우리 가 원하는 상태의 세포만 살기 때문에 그 확률이 낮아도 결 국엔 원하는 세포를 얻을 수가 있다. 이와 달리, SSOD를 이 용한 knock-in은 selection marker의 사용이 불가능 하기 때문에 성공확률이 낮을 수 밖에 없는 문제점을 안고 있다.
그렇기 때문에 본 필자가 경험에 의해 얻어진 몇 가지 유용 한 정보를 소개하고자 한다. DNA ligase IV 기능을 방해하 는 SCR7이라는 화학물질을 쓰면, 유전자가위로 자른 DNA 가 조금 더 오래 잘라진 상태를 유지 하기 때문에 5-10배의 knock-in 효율의 상승 효과를 볼 수가 있다[2]. 위에서 언 급한 것과 같이 knock-in 은 '타이밍'이 중요하기 때문에, 정제된 상품의 Cas9 단백질을 사용하면 SSOD가 사라지기 전에 효과적으로 표적 유전자를 잘라 주어 그 효과가 상당히 상승되는 것을 볼 수가 있다. 또한, gRNA나 SSOD의 반감 기를 연장 시킬 수 있도록 각각의 말단 부분에 있는 염기의 화학적 구조를 변형 시키는 방법이 있다. 예를 들어서, ‘메틸’
을 첨가하든가, ‘황’과’메틸’을 같이 첨가하면 반감기가 50배 이상 증가하는 것을 보여준다[3]. SSOD를 디자인 할 때, 가 장 효과적인 HR을 유도해 내는 공식은 다음과 같다. 유전자 가위의 절단 부위를 기준으로 upstream쪽으로 91 base pair 그리고, downstream쪽으로 36 base pair 로 설계하여 양 말 단이 127 bases 만큼의 비대칭 올리고머를 제작하고, SSOD 는 sense strand DNA sequence를 취하면 된다[4]. 지금껏 여러 가지 의견이 분분 했지만, 최근에 발표된 논문에서 다양 한 경우의 조건들을 검증하고서 내린 결론으로 본 방법을 사 용하면 가장 높은 효율을 얻을 것으로 기대된다. 2013년에 CRISPR/Cas9 system이 처음 나왔을 때부터 지금까지 계속 되는 논쟁 중 하나로, 상당히 높은 효율에 비해 ‘off-target’
비율 또한 높다는 문제가 있다. 게놈 전체를 대상으로 작용하 므로 원하지 않는 부위의 절단과 그로 인한 원하지 않는 유전 자 변형의 가능성을 안고 사용을 해야만 한다. 2016년 1월에
‘Science’에 이러한 문제를 줄여줄 수 있는 방법이 소개 되었
는데, Cas9의 아미노산 서열을 재 정돈 하여, 양성전기를 띄 어 DNA에 강하게 결합할 수 있는 Lysine 과 Arginine을 무 극성의Alanine으로 바꾸어주어, off-target에 부정확하게 붙을 기회를 낮추는 방법이다[5]. 논문에 의하면 off-target 이 극적으로 감소하고, 연구상 원하는 목표부위만 변형시키 고 다른 부분은 정상을 유지한다. 이와같이 지금 이 순간에도 CRISPR의 단점들이 많은 연구진들에 의해 활발히 보완되고 있는 상태이다.
선천성 면역결핍환자를 위한 유전자가위 기반 유전자치료의 임상 적용 사례
X-linked chronic granulomatous disease (X-CGD) 는 출생 시부터 우리 몸을 보호해 주는 다형핵 백혈구의 주 요 기능중 하나인 활성산소를 만들어 내지를 못해 박테리아 나 곰팡이균에 감염되어 결국 목숨을 잃는 유전병중 하나이 다. 환자들의 75%가 gp91phox[그림3]라는 유전자에서 하 나의 염기가 잘못 되어서 병을 유발 하는데, 유전자가위로 잘못된 하나의 염기만 정상 염기로 바꿔주면 되기 때문에 유 전자가위를 이용한 치료 대상으로 최적의 모델이 될 수 있 다. 또한 전체 다형핵 백혈구 중 5% 만 정상적 기능을 해도
그림 3. Diagram of phox gene family in Neutrophil
생명에 지장 없이 살 수 있기 때문에 100%의 치료를 목표 로 하지 않아도 되는 장점이 있다. 이와 더불어, 모든 혈액 세포들은 골수에 있는 골수 조혈모 줄기세포로부터 유래 되 고, 이 줄기세포를 이용해 유전자가위를 이용한 치료를 하 면 최종적으로 원하는 다형핵 백혈구세포의 기능을 지속적 으로 정상화 할 수 있어 더욱 효과적이라고 할 수 있다. 먼저 X-CGD환자의 조혈모 줄기세포를 Granulocyte-Colony Stimulating Factor (G-CSF)라는 사이토카인을 주사해서 골수에 있는 세포를 몸 전체 혈관으로 끌어낸 뒤 ‘Apheresis’
라는 과정을 거쳐서 백혈구만을 피 속에서 분리한 후, 나노 미터 단위의 작은 금속이 결합된 CD34 (조혈모 줄기세포의 표면에 존재하는 항원)항체를 이용해서 치료에 사용할 진짜 조혈모 줄기세포 만을 골라낸 다음 액화질소탱크에 잘 보관 해 놓는다. 세포가 확보되면 이번엔 CRISPR/Cas9 system
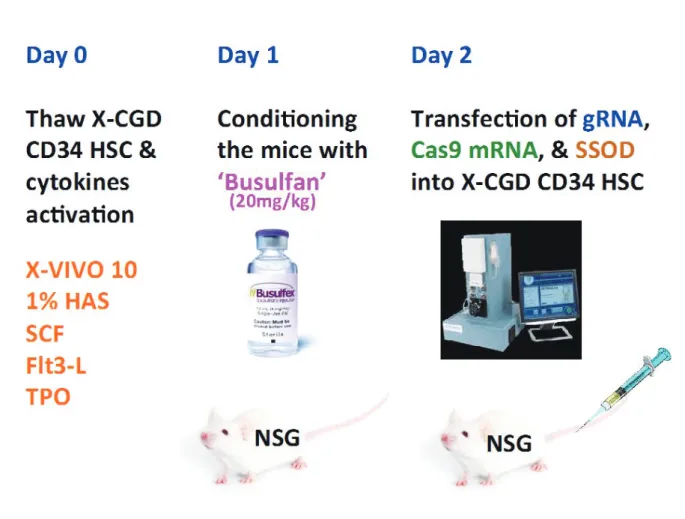
과 SSOD를 위에서 언급한 바와 같이 준비하고, 여러 가지 조합으로 최상의 결과가 나올 때까지 실험을 통해서 최고의 조건을 찾아낸다. 이때 gRNA와 Cas9 mRNA 그리고 SSOD 를 환자의 조혈 모 줄기세포에 넣어주는 과정이 가장 중요한 기술로 작용하는데 기존의 전기충격 방법으로는 많은 세포 들이 죽기 때문에 적합하지 않고, 임상용으로 개발된 새로운 전기충격 기계인 ‘MaxCyte GT’을 이용하면, 거의 모든 세 포들이 살아남고, 그 중 살아있는 세포의 25~50% 정도의 잘못된 염기를 정상인 염기로 바꾸어 줄 수가 있다. 이렇게 처리한 환자의 조혈모 줄기세포는 면역기능이 완전히 제거 된 실험용 생쥐인 NOD scid gamma (NOD.Cg-Prkdcscid Il2rgtm1Wjl/SzJ), NSG mouse에 먼저 Busulfan (20mg/
kg)으로 생쥐의 골수를 일부 제거한 후 정맥 주사한다[그림 4]. 두달 후에 골수와 혈액에서 샘플을 수거해 얼마나 환자
그림 4. In vivo experiment of CRISPR/Cas9 & SSOD with CD34 HSC in NSG mouse
의 백혈구에서 치료효과가 있는지를 flow cytometry 를 통 해서 확인한다. 최근에 본 연구실에서 시도한 동물 실험결 과는 상당히 고무적이다. 거의 정상세포의 1/3에 해당하는 10%정도의 환자세포가 치료효과를 보였다[그림5]. 당초 목 표였던 5%의 2배의 효과를 보인 것이다. 물론 off target에 의한 게놈의 손상여부를 검사하는 과정이 반드시 수반되어 야 하며, 이는 deep sequencing을 통해 이미 확인하였다. 검 사결과 off target에 의한 손상은 존재 하지 않은 것으로 보 여진다. 현재 이 방법을 이용한 유전자치료 임상 프로토콜이 미국립보건원 (NIH)에서 통과가 되어 올해 안에 환자에게 적 용할 계획에 있다.
결론
제한효소가 발견된 지 거의 반세기가 지난 현재 우리는 유 전자가위라는 또 하나의 발견을 통해 생명공학의 발전과 이
를 통한 인류의 건강 증진 도모라는 중요한 분기점에 놓여 있다. ‘부뚜막의 소금도 넣어야 짜다’는 우리말 속담이 있듯 이, 아무리 좋은 도구를 가지고 있어도 이를 잘 이용하지 않 고 바라만 보면서는 아무런 변화도 생기지 않는다. 필자의 바램은 21세기를 살아가는 우리들이 모든 두려움을 떨쳐 버 리고 포기 할 줄 모르는 마음 가짐으로 최선을 다해 꾸준 히 정진할 때 비로서 우리는 우리가 원하는 것들을 이루어 낼 수 있으리라 굳게 믿는다. 필자는 새로 오는 학생이나 박 사 후 과정에 있는 분들에게 늘 하는 말이 있다. 우리가 하 는 ‘Research’는 ‘Search’ 앞에 Re (다시)를 붙여서 만든 단 어이다. 즉 다시 말해 한 번에 되면 ‘Search’이고 우리가 하 는 ‘Research’는 한 번에 되지 않기 때문에 다시 찾는다는 의 미의 ‘Re-Search’ 라고 말한다. 다시 말해서, 실패를 두려워 하지 말라는 말이다. 마지막으로 하고 싶은 말은 유전자 가 위는 결코 어렵거나, 비싼 돈을 지불해야만 얻을 수 있는 것 이 아니라, 어느 랩에서나, 누구든지 원하는 사람은 손쉽게 만들어 쓸 수 있는 일반적인 도구라는 것을 다시 한 번 힘주 그림 5. Gene marking of corrected gp91phox gene in X-CGD CD34+ HSC in chimeric population of NSG mouse BM.
어 강조하는 바이다. 이 글을 접하고, 관심 있는 분들은 연락 을 주시기 바라며, 제가 아는 한도 내에서 최선을 다해 도와 드릴 것을 약속 드립니다. 감사합니다.
1. Isaac B Hilton et al. Epigenome editing by a crisPr-cas9-based acetyltransferase activates genes from promoters and enhancers. Nat.
Biotechnol. 33, 510 (2015)
2. Van Trung Chu et al. Increasing the efficiency of homology-directed repair for CRISPR-Cas9-induced precise gene editing in mammalian cells. Nat. Biotechnol. 33, 543 (2015)
3. Ayal Hendel et al. Chemically modified guide RNAs enhance CRISPR- Cas genome editing in human primary cells. Nat. Biotechnol. 33, 985 (2015)
4. Christopher D Richardson et al. Enhancing homology-directed genome editing by catalytically active and inactive CRISPR-Cas9 using asymmetric donor DNA. Nat. Biotechnol. 34, 339 (2016)
5. Ian M. Slaymaker et al. Rationally engineered Cas9 nucleases with improved specificity. Science 351, 84 (2016)
참고문헌
최 의 묵
1983–1990 중앙대학교 문리과대학 생물학과, 학사 1990–1992 연세대학교 암연구소 연구원
1995– 1999 State University of New York at Buffalo, Microbiology/Immunology Dept., 박사 1999 – 2008 Laboratory of Host Defense, National
Institute of Allergy and Infectious Disease, in NIH박사후 과정 및 선임연구원
2009 – 현재 Staff Scientist in LHD, NIH