ꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏ
<접수일:2003년 9월 18일, 심사통과일:2003년 10월 13일>
※통신저자:배 상 철
서울특별시 성동구 행당동 17번지 한양대학교 의과대학 류마티스병원
Tel:02) 2290-9237, Fax:02) 2298-8231, E-mail:[email protected]
전신성 홍반성 루푸스 신염 활성도와 소변 Transforming Growth Factor-β Induced Gene-h3 (βig-h3)의 연관성
한양대학교 의과대학 류마티스병원, 신장내과*, 병리학교실**, 경북대학교 생화학교실***
김 일․홍철호․조현석․유선진․백창희․이혜순 엄완식․김태환․전재범․유대현․김신규․이창화*
강종명*․박문향**․김인산***․배상철
= Abstract =
Urinary Transforming Growth Factor-β Induced Gene-h3 (βig-h3) as a Marker of Lupus Activity in SLE with Nephritis
Il Kim, M.D., Cheol-Ho Hong, M.D., Hyun-Seok Cho, M.D., Sun-Jin You, M.D., Chang-Hee Paik, M.D., Hye-Soon Lee, M.D., Wan-Sik Uhm, M.D., Tae-Hwan Kim, M.D., Jae-Bum Jun, M.D., Dae-Hyun Yoo, M.D., Think-You Kim, M.D., Chang-Hwa Lee, M.D.*,
Chong-Myung Kang, M.D.*, Moon-Hyang Park, M.D.**, In-San Kim, M.D.***, Sang-Cheol Bae, M.D.
The Hospital for Rheumatic Diseases, Division of Nephrology*, Department of Pathology**, Hanyang University Medical Center, Cell and Matrix Biology National Research Laboratory, Department of
Biochemistry, Kyungpook National University School of Medicine***
Background: TGF-β-induced gene-h3 (βig-h3) is a novel gene induced by active TGF-β and the association with other renal disease is reported. Lupus nephritis is characterized by excessive extracelluar matrix accumulation and the implication that TGF-β is increased in lupus nephritis is known. We measured the urinary βig-h3 in lupus nephritis and sought its association with the activity of lupus nephritis through renal biopsy. The objective of this study was to examine urinary βig-h3 excretion in lupus nephritis and the association with activity of lupus nephritis.
서 론
전신성 홍반성 루푸스(이하 루푸스)는 원인이 불 분명한 질환으로 자가항체와 면역복합체에 의해 세 포와 조직의 손상이 일어나며, 루푸스 신염은 이러 한 면역복합체의 침착으로 인한 염증성, 면역학적 기전에 의해 세포외기질의 축적과 섬유화를 특징으 로 하는 질환이다1).
루푸스 신염의 급성반응은 혈중면역세포와 신실질 세포들의 활성화에 의한 사이토카인(cytokine)과 성장 인자들의 생성이 특징적이며, 이후의 만성반응에는 reactive oxygen species (ROS)와 TGF-β에 의해 세포외 기질이 과도하게 축적되고 말기 신질환으로 진행된 다1,2).
TGF-β는 신질환에서 사구체 경화증과 세뇨관간질 의 섬유화를 초래하는 중요한 사이토카인(cytokine) 으로3), 조직 내에서는 비활성화된 형태로 존재하다 가 효소성4) 또는 비효소성5) 과정에 의해서 활성화되 어 생물학적인 기능을 하게 된다. 또한, 다른 인자들 에 의해서 생물학적 활성도가 조절되기 때문에6,7), 단 순히 TGF-β mRNA, 단백질의 측정만으로는 그 생물 학적 활성도를 알기 어려우며 기존의 생물학적 분석 법은 임상에 적용하기 어려운 점이 있다8,9).
최근에, 인간 선암종세포에서 TGF-β에 의해서 유 도되는 TGF-β-induced gene h3 (βig-h3)가 발견되었
으며, TGF-β의 생물학적 활성도를 나타내는 지표로 서 연구되고 있다. 이미, 당뇨병성 신병증과 cyclo- sporin A 신병증에서 신장내의 βig-h3의 발현이 증가 되어 있고, TGF-β와 βig-h3의 발현이 상관관계가 있 음이 보고되었다10,11).
지금까지 루푸스 신염의 활성도를 반영하는 지표 로서 보체(C3, C4), 항dsDNA 항체(anti-dsDNA Ab)와 같은 비침습적인 방법과 신장조직검사를 통해 활성 도(Activity Index), 만성도(Chronicity Index)를 측정하 는 침습적인 방법이 사용되었으나, 이러한 방법들로 는 루푸스의 활성도를 완전하게 평가하기 어렵다. 저 자들은 루푸스 신염 환자에서 소변 βig-h3을 측정하 여 각종 루푸스 활성도 인자와의 연관성을 비교 연 구하였다.
대상 및 방법 1. 대상 환자
2002년 11월부터 2003년 2월 사이에 한양대학교 류마티스병원에서 진단 받은 15명의 루푸스신염 환 자를 대상으로 하였다. 전체 대상자의 평균연령은 32.6±2.9세였고 성별은 여자가 13명, 남자 2명이었 다. 모든 환자는 1997년 미국 류마티스학회에서 정 한 진단기준에 적합하였다12).
Methods: Fifteen patients (median age 32.6±2.9 years, range 18∼64) who developed lupus nephritis underwent renal biopsy. At the time of biopsy, they showed significant pro- teinuria. Total urinary βig-h3 concentration was assayed by enzyme-linked immunoabsor- bent assay and expressed as a ratio to urinary creatinine concentration.
Results: There were correlations between urinary βig-h3 and the reduction of C3 (r=-0.566, p=0.028<0.05), the magnitude of proteinuria (r=0.531, p=0.042<0.05). The Activity Index, Chronicity Index in the renal biopsy, C4, anti-dsDNA Ab titer were not significantly cor- related with urinary βig-h3 excretion, but the patients with high Activity Index had the in- creased level of urinary βig-h3. Five patients who had fibrinoid necrosis in renal biopsy showed higher level of urinary βig-h3 than the others (107.78±43.02 vs. 50.21±10.12 ng/
ml, p=0.061)
Conclusion: In this study, There is some correlation between urinary βig-h3 and the ac- tivity of lupus nephritis. Urinary βig-h3 may play a role in predicting the active lupus ne- phritis. A further study is needed in large population and in situ expression of βig-h3.
ꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏ Key Words: βig-h3, Lupus nephritis, TGF-β
2. 소변내 βig-h3의 측정
소변은 신장조직검사 시에 같이 채취되었으며, 소 변내 βig-h3의 측정은 ELISA법으로 측정하였다(Reg- en Biotech, Seoul Korea). 방법을 간단히 설명하면 다 음과 같다13). 96-well ELISA 플레이트를 0.5μg/ml βig- h3 단백질로 4oC에서 12시간 코팅한 후 PBS-0.05%
Tween 20 (PBST) 용액으로 3회 세척하였다. 별도의 96-well 플레이트에서 소변을 βig-h3 항체(PBST로 1:2,000 희석)와 37oC에서 90분 반응시킨 후 βig-h3 이 코팅된 플레이트로 옮겼다. 30분간 실온에서 반 응시킨 후 PBST용액으로 3회 세척하고 peroxidase- conjugated antirabbit IgG 항체(1:2,000희석)와 실온 에서 90분 반응시켰다. 다시 PBST용액으로 3회 세척 후 200 ul의 기질용액(100μg/ml ο-phenylenediamine, 0.003%
H2O2)을 넣고 어두운 실온에서 60분간 발색시켰다.
50 ul의 8 NH2SO4 용액으로 반응을 정지시키고 490 nm에서 흡광도를 읽었다.
3. 신장조직검사
신장조직검사는 소변채취 시에 같이 시행되었다.
WHO분류에 의해서 구분되었으며 Austin 등이 제시 한 방법에 의거하여 활성도(Activity Index), 만성도 (Chronicity Index)를 계산하였다14). 활성도는 glomer- ular hypercelluarity, cellular crescents, leukocyte exu- dation, karyorrhexis/fibrinoid necrosis, hyaline deposits, interstitial inflammation, 만성도는 glomerular sclero- sis, fibrous crescents, tubular atrophy, interstitial fibro- sis의 합으로 측정되었다.
4. 임상 검사
소변 내 βig-h3 측정과 함께 보체 C3, C4, 항 ds-DN A항체, 24시간 단백뇨 등에 대하여 검사를 시행하였 다.
5. 통계 분석
결과는 산술평균±평균의 표준오차(standard error of mean, SEM)로 표시하였고, 피어슨 상관계수 (Pearson's correlation coefficient)를 사용하여 상관분 석을 하였다. t-test를 통하여 βig-h3의 평균값을 비 교하였으며, p<0.05일 때 통계적으로 유의하다고
판정하였다. 통계처리는 Window용 SPSS 11.0 프로 그램(SPSS for window, 2001)을 사용하여 분석하였 다.
결 과 1. 환자의 임상적 특성
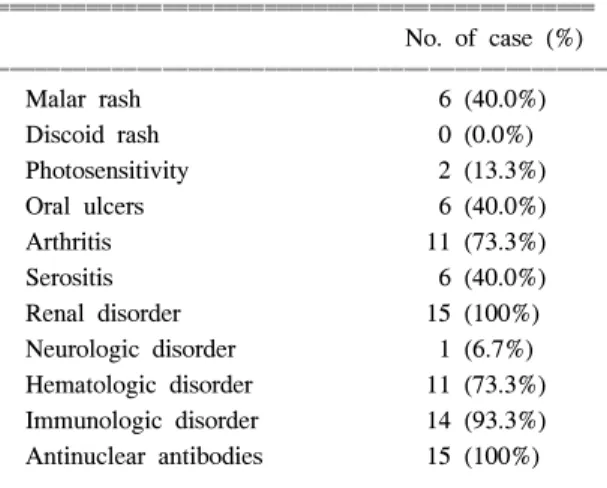
대상환자 중 협부발진(malar rash)은 6명(40.0%), 광과 민증(photosensitivity)은 2명(13.3%)에서 있었으며, 원반 상발진(discoid rash)을 보인 환자는 없었다. 구강궤양 (oral ulcer)은 6명(40.0%), 관절염은 11명(73.3%), 장막 염(serositis)은 6명(40.0%)에서 보였고, 단백뇨는 평균 2561±397 mg/day였고, 3.0 g/day 이상의 단백뇨를 보인 환자는 5명이었다. 신경질환은 1명(6.7%), 혈액질 환은 11명(73.3%), 면역질환(immunologic disorder)은 14명(93.3%), 항핵항체는 15명(100%)의 환자에서 양성소 견을 나타냈다(표 1). 신장조직검사에서 WHO cla- ssification II형이 1명, III형이 1명, III to IV형이 1명, IV형이 3명, V형이 3명이었고, V+III형이 5명, V+
IV형이 1명이었다(표 2).
2. 소변 βig-h3과 Activity Index, Chronicity Index 와의 관계
소변 βig-h3은 Activity Index (r=0.479, p=0.071), Chronicity Index (r=-0.106, p=0.706)와 통계적으로 유의한 상관관계는 없었으나 Activity Index가 높을수
Table 1. The ACR criteria of SLE patients
ꠧꠧꠧꠧꠧꠧꠧꠧꠧꠧꠧꠧꠧꠧꠧꠧꠧꠧꠧꠧꠧꠧꠧꠧꠧꠧꠧꠧꠧꠧꠧꠧꠧꠧꠧꠧꠧꠧꠧꠧꠧꠧꠧꠧꠧꠧꠧ
No. of case (%) ꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏ
Malar rash 6 (40.0%)
Discoid rash 0 (0.0%)
Photosensitivity 2 (13.3%)
Oral ulcers 6 (40.0%)
Arthritis 11 (73.3%)
Serositis 6 (40.0%)
Renal disorder 15 (100%) Neurologic disorder 1 (6.7%) Hematologic disorder 11 (73.3%) Immunologic disorder 14 (93.3%) Antinuclear antibodies 15 (100%) ꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏ
록 소변 βig-h3이 증가되는 경향을 보였고(그림 1, 2), 루푸스 신염의 Activity Index의 매개변수(glomerular hypercelluarity, leukocyte exudates, hyaline deposit, fibri- noid necrosis, cellular crescents)와도 통계학적인 유의성은 없었으나, fibrinoid necrosis를 보인 환자(n=3)에서 소변 βig-h3 level이 증가되어 있었다(107.78±43.02 vs. 50.21±
10.12 ng/ml, p=0.061) (그림 3, 표 3).
3. 소변 βig-h3과 보체 C3, C4, 항ds-DNA항체, 단백 뇨와의 관계
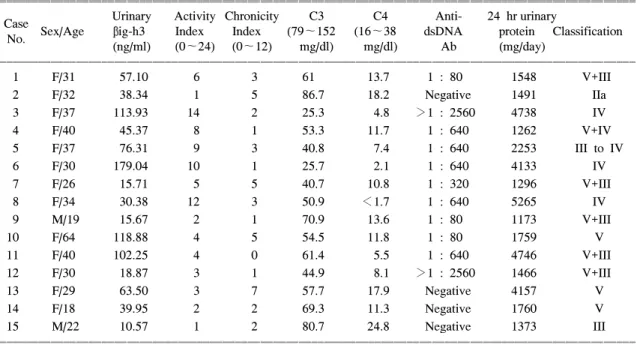
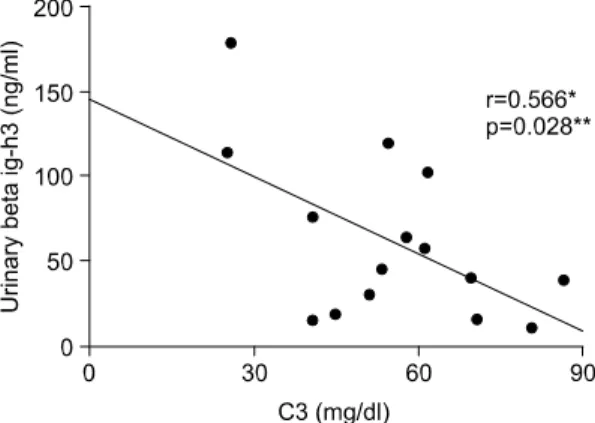
소변 βig-h3과 루푸스 신염의 활성도를 비교하기 위해 보체 C3, C4, 항ds-DNA항체, 단백뇨와 비교하 였다. 보체 C3는 소변 βig-h3과 음의 상관관계를 보 였으나(r=-0.566, p=0.028<0.05) (Fig. 4), 보체 C4 (r=-0.511, p=0.052), 항ds-DNA항체(r=0.271, p=0.329) Table 2. Clinical and histologic data of lupus nephritis
ꠧꠧꠧꠧꠧꠧꠧꠧꠧꠧꠧꠧꠧꠧꠧꠧꠧꠧꠧꠧꠧꠧꠧꠧꠧꠧꠧꠧꠧꠧꠧꠧꠧꠧꠧꠧꠧꠧꠧꠧꠧꠧꠧꠧꠧꠧꠧꠧꠧꠧꠧꠧꠧꠧꠧꠧꠧꠧꠧꠧꠧꠧꠧꠧꠧꠧꠧꠧꠧꠧꠧꠧꠧꠧꠧꠧꠧꠧꠧꠧꠧꠧꠧꠧꠧꠧꠧꠧꠧꠧꠧꠧꠧꠧꠧꠧꠧꠧꠧꠧ
Urinary Activity Chronicity C3 C4 Anti- 24 hr urinary
CaseNo. Sex/Age βig-h3 Index Index (79∼152 (16∼38 dsDNA protein Classification (ng/ml) (0∼24) (0∼12) mg/dl) mg/dl) Ab (mg/day)
ꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏ
1 F/31 57.10 6 3 61 13.7 1 : 80 1548 V+III
2 F/32 38.34 1 5 86.7 18.2 Negative 1491 IIa
3 F/37 113.93 14 2 25.3 4.8 >1 : 2560 4738 IV
4 F/40 45.37 8 1 53.3 11.7 1 : 640 1262 V+IV
5 F/37 76.31 9 3 40.8 7.4 1 : 640 2253 III to IV
6 F/30 179.04 10 1 25.7 2.1 1 : 640 4133 IV
7 F/26 15.71 5 5 40.7 10.8 1 : 320 1296 V+III
8 F/34 30.38 12 3 50.9 <1.7 1 : 640 5265 IV
9 M/19 15.67 2 1 70.9 13.6 1 : 80 1173 V+III
10 F/64 118.88 4 5 54.5 11.8 1 : 80 1759 V
11 F/40 102.25 4 0 61.4 5.5 1 : 640 4746 V+III
12 F/30 18.87 3 1 44.9 8.1 >1 : 2560 1466 V+III
13 F/29 63.50 3 7 57.7 17.9 Negative 4157 V
14 F/18 39.95 2 2 69.3 11.3 Negative 1760 V
15 M/22 10.57 1 2 80.7 24.8 Negative 1373 III
ꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏ
Fig. 1. Correlation between the urinary βig-h3 and Activity Index.
Fig. 2. Correlation between the urinary βig-h3 and Chronicity Index.
와는 상관관계가 없었다. 단백뇨는 소변 βig-h3과 양의 상관관계를 보였으며(r=0.531, p=0.042<0.05) (Fig. 5), WHO classification III, IV인 환자군(n=5)과 class II, V인 환자 군(n=10)간의 소변 βig-h3을 비교 하였을 때 통계적인 유의성은 없었으나(82.05±30.20
vs. 51.56±11.19 ng/ml, p=0.264) class III, IV 환자 군에 서 소변 βig-h3의 증가되었다(Fig. 6).
고 찰
루푸스 신염의 활성도를 나타내는 신장조직검사상 의 Activity Index와 소변 βig-h3은 비록 통계적으로 유의한 상관관계는 없었으나(r=0.479, p=0.071) Acti- vity Index가 높은 환자에서 소변 βig-h3이 증가된 경향을 보였다. Fibrinoid necrosis를 보인 3명의 환자 에서는 통계적인 유의성은 없었으나 소변 βig-h3이 증가되었고(107.78±43.02 vs. 50.21±10.12 ng/ml, p=
0.061), WHO class III, IV를 보인 5명의 환자에서도 역 Fg. 3. Urinary βig-h3 in patients with (A) and without
(B) fibrionoid necrosis.
Table 3. Urinary βig-h3 and fibrinoid necrosis in renal biopsy
ꠧꠧꠧꠧꠧꠧꠧꠧꠧꠧꠧꠧꠧꠧꠧꠧꠧꠧꠧꠧꠧꠧꠧꠧꠧꠧꠧꠧꠧꠧꠧꠧꠧꠧꠧꠧꠧꠧꠧꠧꠧꠧꠧꠧꠧꠧꠧꠧ
Fibrinoid No fibrinoid
necrosis necrosis p value (n=3) (n=12)
ꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏ Urinary βig-h3
107.78±43.02 50.21±10.12 0.061 (mean, ng/ml)
ꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏ
Fig. 4. Correlation between the urinary βig-h3 and C3.
*r>0.5, **p<0.05.
Fig. 5. Correlation between the urinary βig-h3 and proteinuria *r>0.5, **p<0.05.
Fig. 6. Urinary βig-h3 in patients with class III, IV versus class II, V. *r>0.5, **p<0.05.
시 통계적인 유의성은 없었으나 소변 βig-h3이 증가 되어 있었다(82.05±30.20 vs. 51.56±11.19 ng/ml, p=
0.264). 그리고, C3는 소변 βig-h3과 음의 상관관계 를 보였으며(r=-0.566, p=0.028), 단백뇨는 소변 βig- h3와 양의 상관관계를 보였다(r=0.531, p=0.042). 다 른 인자들은 상관관계를 보이지 못했다.
루푸스 신염은 면역복합체의 침착에 의해 염증성, 면역성 반응이 초래되어 세포외기질이 과도하게 침 착되고 사구체, 세뇨관, 혈관, 간질의 섬유화가 나타 나는데1), 이러한 과정은 혈중면역세포와 신실질세포 들이 활성화되어 사이토카인(cytokine) 등과 같은 염 증매개성 물질과 자유산소라디칼, 산화질소, matrix metalloproteinase 등과 같은 독성물질을 분비되어 신 조직 손상이 일어나는 급성반응과 아교질과 세포외 기질이 과도하게 침착되어 말기신질환으로 진행되는 만성반응을 보이는데, TGF-β는 만성 반응 시에 주 된 역할을 한다1,2,15).
TGF-β는 다양한 기능을 갖는 사이토카인(cytoki- ne)으로, 세포증식을 억제 또는 자극함으로써 조직치 유와 재생에 관여한다3). 이러한 기능은 TGF-β가 기 질성분인 섬유결합소, 아교질16), 프로테오글리칸의17) 합성을 자극하고, 기질분해에 작용하는 단백분해효 소의 합성을 저해하며, 단백분해효소억제제를 증가 시킴으로써9), 나타나게 된다. 또한, TGF-β는 사구체 신염18), 폐섬유화19), 간경화20), 안질환21)과 피부질환22) 에서 반흔형성 시 세포외기질의 침착과 섬유화를 촉 진시키고, 항섬유용해제(plasminogen activator inhibit- or-1: PAI-1)와 응고인자(tissue factor: TF)의 유전자발 현을 유도하여 섬유소용해를 억제한다.23,24).
신장질환에서 TGF-β는 사구체경화증과 세뇨관간 질의 섬유화를 일으키는데3), 이것은 사구체신염의 동물모델에서뿐만 아니라 인체에서도 당뇨병성 신병 증21), IgA 신병증, 증식사구체염25), 반월형 사구체염, 루푸스신염 같은 증식성 사구체 질환에서2) 사구체내 TGF-β의 발현이 증가되어 있음이 여러 연구에서 보 고되었다. 또한, 동물실험에서 TGF-β 투여로 신장내 세포외기질의 병적증가와 조직섬유화가 일어나고3), 혈관간기질의 확대에도 TGF-β가 관여하며26), TGF-β 에 대한 중화항체의 투여로 사구체 내 세포외기질의 축적을 감소시킬 수 있다는 연구 결과들은27) TGF-β 가 사구체질환을 진행시킨다는 것을 보여준다.
그리고, 루푸스 신염에서의 TGF-β에 관한 연구들 을 보면 루푸스 신염의 만성반응으로 TGF-β가 지속 적인 과도한 세포외기질의 침착을 일으켜 루푸스 신 염을 진행시키는 역할을 하고1), 루푸스 신염을 포함 한 증식성 사구체 질환에서 사구체 내 TGF-β수용체 의 발현이 증가된 경우 심한 증식성 사구체병변과 상관관계가 있음이 보고되었으며28), TGF-β mRNA가 미만성 루푸스 신염에서 증가되어 있다는29) 연구 결 과들은 TGF-β가 루푸스 신염의 진행에 있어서 중요 한 역할을 한다는 것을 보여준다. 또한, 본 연구에서 사구체질환의 만성반응에 관여하여 세포외기질의 축 적을 일으키는 TGF-β의 활동성 지표인 βig-h3의 활 성도가 높은 루푸스 신염 환자에서 증가되는 경향을 보인 것은 TGF-β가 급성반응 시에도 신장 내에서 발현되는 것으로 생각된다. 신장 내에서 TGF-β의 분포에 대해서는 잘 알려져 있지는 않지만, 루푸스 마우스 모델에서(MRL lpr/lpr) 이루어진 연구결과를 보 면 사구체세포와 침윤된 염증세포에서 TGF-β mRNA 의 발현을 보고하였다2).
TGF-β는 391개의 아미노산으로 구성되며 포유류 에서 3가지 형태로 존재하는데 이 중 TGF-β1이 섬 유화와 가장 관련이 있다3). TGF-β는 TGF-β binding protein과 결합된 비활성화된 형태로 세포표면과 세 포외기질에 저장되었다가 효소성4) 또는 비효소성5) 과 정에 의해 latency-associated peptide (LAP)가 분할되 어 활성화된다.3) 또한 TGF-β의 생물학적 기능은 proteoglycan decorin7), α2-macroglobulin8)에 의해서 조절된다. 따라서, 조직 내의 TGF-β1의 mRNA, 단 백질로는 TGF-β1의 생물학적 활성도를 충분히 반영 하지 못하며 TGF-β1의 활성도를 반영할 수 있는 생 물학적 분석법이 사용되고 있으나, 기존의 방법인 mink 폐 상피세포에 대한 TGF-β의 억제능을 측정 하는 것은 hepatocyte growth factor (HGF)에 의해서 그 효과가 반감될 수 있고9), TGF-β가 섬유결합소 전사의 splicing 형태에 미치는 영향력을 측정하는 것도 다른 성장인자(retinoic acid, 1, 25-dihydroxy vita- mine D3)에 영향을 받을 수 있다는 한계점이 있다10). 최근, 인간 선암종세포(A549)에서 TGF-β에 의해 서 유도된 새로운 유전자인 βig-h3에 대한 연구가 이루어지고 있다. βig-h3은 683개의 아미노산으로 구성되며 N-terminal에는 분비성 아미노산 서열이 있고,
C-terminal은 integrin을 인식할 수 있는 Arg-Gly-Asp (RGD) 아미노산 서열을 갖는다. 이외에 Drosophilia fasciclin-I과 동종의 짧은 아미노산 부위와 4개의 동종의 internal domain으로 구성되어 있다30). βig-h3의 기능에 대한 연구는 최근에 시작되어 많이 알려져 있지 않지만 세포부착과 암 형성에 관여할 것으로 생각되고 있으
며30,31), 이외에도 여러 보고에서 세포성장, 세포분화32),
상처치유33)에 관여하고, 세포외기질의 microfibril-ass- ociated protein (MP 78/80)로서 microfibrillar proteins fibrillin-I, MAGP-1과 결합한다34). TGF-β에 의한 βig- h3의 발현증가는 다양한 세포계에서 발견되는데, 생 체 외에서는 유방상피세포, 섬유모세포, 각질세포, 흑색 종세포33)에서 발견되었고, 생체 내에서는 유두진피31), 각막상피35), 혈관36)에서 TGF-β에 의한 βig-h3의 발현 증가가 보였다.
신장질환에서는 당뇨병성 신병증과 cyclosporin A 신병증에서 TGF-β와 βig-h3 사이에 유의한 상관관 계가 있으며, 정상군에 비해서 βig-h3이 증가되었고, 신장내에서 βig-h3가 발현된다고 보고하였다10,11). 그 리고, 신증후군에서 소변을 통한 TGF-β의 배설이 증가되어 있고37), IgA 신염과 초점성 사구체신염과 같이 세포외기질의 축적이 특징인 질환들에서 소변 TGF-β가 신간질의 섬유화 정도, 혈관간기질의 증가 와 상관관계가 있다는 연구 결과들은38) 세포외기질 의 축적을 보이는 루푸스 신염에서도 소변 βig-h3과 상관관계를 가질 수 있음을 시사한다. 본 연구에서 도 Activity Index가 증가된 경우, class IV, fibrionid necrosis를 보인 환자에서 소변 βig-h3의 증가소견을 보임으로써 루푸스 신염의 활성도를 반영하는 지표 로서 사용될 수 있다는 가능성을 보여주었다. 하지 만, 급성반응 시에 신장 내에서 βig-h3이 발현되는 지에 대한 연구가 필요하다. 또한 βig-h3의 발현이 TGF-β의 생물학적 활성도를 반영하지만 이러한 βig- h3의 발현이 TGF-β에 의해서만 조절되는지는 밝혀 져있지 않으며, βig-h3의 측정이 기존에 사용되고 있는 TGF-β의 생물학적분석보다 우월한지에 대한 비교연구가 필요할 것으로 생각된다.
결 론
본 연구에서는 루푸스 신염 환자에서 TGF-β의 활
성도를 나타내는 소변 βig-h3측정이 루푸스 신염의 활성도를 반영하는 하나의 지표로서 사용될 수 있을 것으로 생각된다.
REFERENCES
1) Grande JP. Mechanisms of progression of renal da- mage in lupus nephritis: pathogenesis of renal sca- rring. Lupus 1998;7:604-10.
2) Yamamoto K, Loskutoff DJ. Expression of transfo- rming growth factor-beta and tumor necrosis factor- alpha in the plasma and tissues of mice with lupus nephritis. Lab Invest 2000;80:1561-70.
3) Border WA, Noble NA. Transforming growth factor beta in tissue fibrosis. N Engl J Med 1994;331:1286- 92.
4) Schultz-Cherry S, Chen H, Mosher DF, Misenheimer TM, Krutzsch HC, Roberts DD, et al. Regulation of transforming growth factor-β activation by discrete sequences of thrombospondin 1. J Biol Chem 1995;
270:7304-10.
5) Oreffo RO, Mundy GR, Seyedin SM, Bonewald LF.
Activation of the bone-derived latent TGF-β complex by isolated osteoclasts. Biochem Biophys Res Com- mun 1989;158:817-23.
6) Yamaguchi Y, Mann DM, Ruoslahti E. Negative re- gulation of transforming growth factor-β by the proteoglycan decorin. Nature 1990;346:281-4.
7) O'Connor-Mccourt MD, Wakefield LM. Latent tran- sforming growth factor β in serum. A specific com- plex with alfa 2-macroglobulin. J Biol Chem 1987;
262:14090-9.
8) Magnuson VL, Young M, Schattenberg DG, Mancini MA, Chen DL, Steffensen B, et al. The alternative splicing of fibronectin pre-mRNA is altered during aging and in response to growth factors. J Biol Chem 1991;266:14654-62.
9) Taipale J, Keski-Oja J. Hepatocyte growth factor rele- ases epithelial and endothelial cells from growth fa- ctor arrest by transforming growth factor-β1. J Biol Chem 1996;271:4342-8.
10) Langham RG, Egan MK, Dowling JP, Gilbert RE, Thomson NM. Transforming growth factor-beta1 and tumor growth factor-beta-inducible gene-H3 in nonre- nal transplant cyclosporine nephropathy. Transplant- ation 2001;72:1725-6.
11) Gilbert RE, Wilkinson-Berka JL, Johnson DW, Cox
A, Soulis T, Wu LL, et al. Renal expression of tra- nsforming growth factor-beta inducible gene-h3 (beta βig-h3) in normal and diabetic rats. Kidney Int 1998;
54:1390-1.
12) Hochberg MC. Updating the American College of Rheumatology revised criteria for the classification of SLE. Arthritis Rheum 1997;40:1725.
13) Lee SH, Bae JS, Park SH, Lee BH, Park RW, Choi JY, et al. Expression of TGF-β-induced matrix pro- tein βig-h3 is up-regulated in the diabetic rat kidney and human proximal tubular epithelial cells treated with high glucose. Kidney International 2003;64:1012- 21.
14) Austin HA III, Muenz LR, Joyce KM, Antonovych TA, Kullick ME, Klippel JH, et al. Progonstic factors in lupus nephritis. Contribution of renal histologic data. Am J Med 1983;75:382-91.
15) Kuroiwa T, Lee EG. Cellular interactions in the pa- thogenesis of lupus nephritis: the role of T cells and macrophages in the amplication of the inflammatory process in the kidney. Lupus 1998;7:597-603.
16) Ignotz RA, Massague J. Transforming growth factor- β stimulates the expression of fibronectin and colla- gen and their incorporation into the extracellular ma- trix. J Biol Chem 1986;261:4337-45.
17) Bassols A, Massague J. Transforming growth factor β regulates the expression and structure of extracellu- lar matrix chondroitin/dermatan sulfate proteoglycans.
J Biol Chem 1988;263:3039-45.
18) Okuda S, Languino LR, Ruoslahti E, Border WA.
Elevated expression of transforming growth factor-β and proteoglycan production in experimental glome- rulonephritis. Possible role in expansion of the mesa- ngial extracellular matrix. J Clin Invest 1990;86:453-62.
19) Broekelmann TJ, Limper AH, Colby TV, McDonald JA. Transforming growth factor-β1 is present at sites of extracellular matrix gene expression in human pulmonary fibrosis. Proc Natl Acad Sci USA 1991;88:
6642-6.
20) Czaja MJ, Weiner FR, Flanders KC, Giambrone MA, Wind R, Biempica L, et al. In vitro and in vivo associ- ation of transforming growth factor-β1 with hepatic fibrosis. J Cell Biol 1989;108:2477-82.
21) Yamamoto T, Nakamura T, Noble NA, Ruoslahti E, Border WA. Expression of transforming growth factor β is elevated in human and experimental diabetic ne- phropathy. Proc Natl Acad Sci USA 1993;90:1814-8.
22) Shah M, Foreman DM, Ferguson MW. Control of scarring in adult wounds by neutralizing antibody to
transforming growth factor β. Lancet 1992;339:213-4.
23) Sawdey MS, Loskutoff DJ. Regulation of murine type 1 plasminogen activator inhibitor gene expression in vivo: Tissue specificity and induction by lipopolysa- ccharide, tumor necrosis factor-α, and transforming growth factor-β. J Clin Invest 1991;88:1346-53.
24) Ranganathan G, Blatti SP, Subramaniam M, Fass DN, Maihle NJ, Getz MJ. Cloning of murine tissue factor and regulation of gene expression by transforming gro- wth factor type β1. J Biol Chem 1991;266:496-501.
25) Yoshioka K, Takemura T, Murakami K, Okada M, Hino S, Miyamoto H, et al. Transforming growth factor-βprotein and mRNA in glomeruli in normal and diseased human kidneys. Lab Invest 1993;68:154-63.
26) Tomooka S, Border WA, Marshall BC, Noble NA.
Glomerular matrix accumulation is linked to inhibition of the plasmin protease system. Kidney Int 1992;42:
146-29.
27) Border WA, Okuda S, Languino LR, Sporn MB, Ru- oslahti E. Suppression of experimental glomerulonephritis by antiserum against transforming growth factor β1. Nat- ure 1990;346:371-4.
28) Onetti Muda A, Feriozzi S, Rahimi S, Faraggiana T.
Expression of TGF-beta receptors type I and II in human glomerulonephritis. Nephrol Dial Transplant 1998;13:279-84.
29) Iwano M, Akai Y, Fujii Y, Dohi Y, Matsumura N, Dohi K. Intraglomerular expression of transforming growth factor-beta1 (TGF-beta1) mRNA in patients with glomerulonephritis: quantitative analysis by com- petitive polymerase chain reaction. Clin Exp Immunol 1994;97:309-14.
30) Skonier J, Neubauer M, Madisen L, Bennett K, Plow- man GD, Purchio AF. cDNA cloning and sequence analysis of beta ig-h3, a novel gene induced in a human adenocarcinoma cell line after treatment with transforming growth factor-beta. DNA Cell Biol 19 92;11:511-22.
31) LeBaron RG, Bezverkov KI, Zimber MP, Pavelec R, Skonier J, Purchio AF. βig-h3, a novel secretory pr- otein inducible by transforming growth factor-β, is present in normal skin and promotes the adhesion and spreading of dermal fibroblasts in vitro. J Invest Dermatol 1995;104:844-9.
32) Dieudonne SC, Kerr JM, Xu T, Sommer B, DeRubeis AR, Kuznetsov SA, et al. Differential display of human marrow stromal cells reveals unique mRNA expression patterns in response to dexamethasone. J Cell Biochem 1999;76:231-43.
33) Skonier J, Bennett K, Rothwell V, Kosowski S, Plo- wman G, Wallace P, et al. Beta ig-h3: a transforming growth factor-beta-responsive gene encoding a secre- ted protein that inhibits cell attachment in vitro and suppresses the growth of CHO cells in nude mice.
DNA Cell Biol 1994;13:571-84.
34) Gibson MA, Hatzinikolas G, Kumartilake JS, Sanberg LB, Nicholl JK, Sutherland GR, et al. Further cha- racterization of proteins associated with elastic fiber microfibrils including the molecular cloning of MA GP-2 (MP-25). J Biol Chem 1996;271:1096-103.
35) Escribano J, Hernando N, Ghosh S, Crabb J, Coca- Prados M. cDNA from human ocular ciliary epithe- lium homologous to βig-h3 is preferentially expre- ssed as an extracellular protein in the corneal epithe-
lium. J Cell Physiol 1994;160:511-21.
36) O'Brien ER, Bennett KL, Garvin MR, Zderic TW, Hinohara T, Simpson JB, et al. Beta ig-h3, a tra- nsforming growth factor-beta-inducible gene, is overe- xpressed in atherosclerotic and restenotic human vas- cular lesions. Arterioscler Thromb Vasc Biol 1996;
16:576-84.
37) Goumenous DS, Tsakas S, El Nahas AM, Alexandri S, Oldroyd S, Kalliakmani P, et al. Transforming growth factor-beta (1) in the kidney and urine of patients with glomerular disease and proteinuria. Ne- phrol Dial Transplant 2002;17:2145-52.
38) Murakami K, Takemura T, Hino S, Yoshioka K. Uri- nary transforming growth factor-beta in patients with glomerular diseases. Pediatr Nephrol 1997;11:334-6.