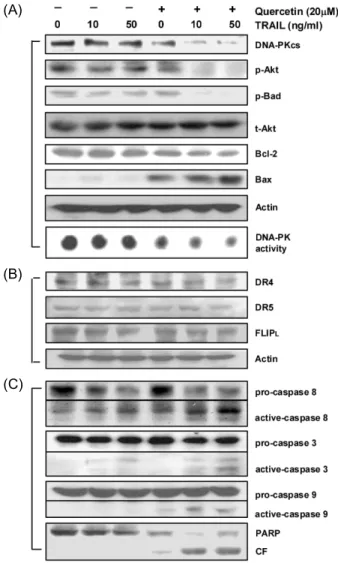
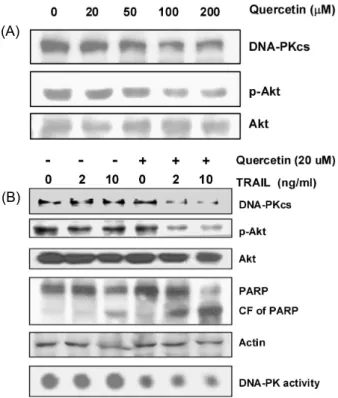
Quercetin Sensitizes Human Leukemic Cells to TRAIL-induced Apoptosis:
Involvement of DNA-PK/Akt Signal Transduction Pathway
Jun-Ik Park, Mi-Ju Kim, Hak-Bong Kim, Jae-Ho Bae, Jae-Won Lee, Soo-Jung Park
1, Dong-Wan Kim
2, Chi-Dug Kang and Sun-Hee Kim*
Department of Biochemistry, Pusan National University School of Medicine, Busan 602-739, South Korea
1
Division of Intractable Diseases, Center for Biomedical Sciences, National Institute of Health, Seoul 122-701, South Korea
2