Note
Analysis of quaternary structure of leucine-responsive regulatory protein (Lrp) by crosslink experiments
Euiho Lee†, Robert Pokoo†, Loi Thuan Nguyen, and Chan Yong Lee*
Department of Biochemistry, Chungnam National University, Daejeon 34134, Republic of Korea
교차결합 실험을 통한 루신 대응 조절 단백질의 4차 구조 분석
이의호† ・ 로버트 포쿠† ・ 로이 트엔 응구엔 ・ 이찬용*
충남대학교 생화학과
(Received August 23, 2017; Revised November 21, 2017; Accepted November 22, 2017)
†These authors contributed equally to this work.
*For correspondence. E-mail: [email protected];
Tel.: +82-42-821-5482; Fax: +82-42-822-7548
Leucine-responsive regulatory protein (LRP) is a regulatory protein of molecular weight 18.8 kDa and is widely known to regulate many metabolic and functional activities of operons in Escherichia coli. The gene for Lrp from Escherichia coli in pQE system of 6 × His-tagging was expressed and 3H-labeled protein, as well as the wild type Lrp, was purified. The crosslink experiments were performed to analyze the quaternary structure of Lrp at high of 5 μM and at low concentrations below 0.3 μM with cross linkers, such as glutaraldehyde, 1, 2, 3, 4-diepoxy- butane (DEB), and ethylene glycol bis (succinimidyl succinate) (EGS). In the experiments, we found that the Lrp protein can be formed higher conformation states of tetramer, hexamer, octamer, as well as dimeric state when incubated with the above cross linkers.
Keywords: crosslink, global regulatory protein, leucine, Lrp
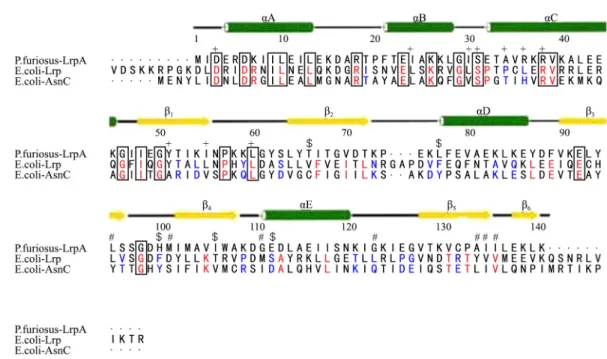
Leucine responsive regulatory protein (Lrp) from Escherichia coli has been identified to consist of 164 amino acids; it crystallizes as an octamer, but assembles as a tetramer of dimers (Ernsting et al., 1992). The Lrp monomer is shown to consist of three regions (Brinkman et al., 2003) namely, The N-terminal HTH domain (Hart et al., 2011), the C-terminal
domain (Chen et al., 2001), and the flexible linker peptide (colored red, green, and blue, respectively, in Fig. 1). They also show that only about 10% relationship with well known regulatory proteins such as Crp or Fnr and to other DNA binding proteins, such as Integration Host Factor (IHF) or the Histone like regulatory proteins (H-NS) (Hart and Blumenthal, 2011). The only similarity between Lrp protein and that of the above named regulatory proteins is that they all bind to DNA to regulate the transcription of their respective operons (Shimada et al., 2015).
Despite this difference, Lrp is related to AsnC, an E. coli protein that regulates the expression of asnA which encodes asparagine synthetase A (Ettema et al., 2002). Previously, it has been confirmed that amino acid sequences of Lrp and AsnC are 25% identical and another 25% of them show a close similarity (Philip et al., 2001). This confirmation has led to the classification of Lrp/Asn family of regulatory proteins (Deng et al., 2011; Hart and Blumenthal, 2011).
Lrp is a global regulatory protein that affects the expression of multiple genes and operons in bacteria (Newman et al., 1992, 1996). Previous research on Lrp protein has demonstrated that it has an isoelectric point (pI) of 9.3 and a monomeric molecular mass of 18.8 kDa (Calvo and Matthews, 1994). It has also been postulated that, at the concentration of 10 μM, Lrp protein
Fig. 1. Secondary structure of three domains of Lrp. The positions of the E. coli Lrp DNA binding mutants, activation mutants and leucine response mutants are indicated by the symbols +, $ and #, respectively (Barton, 1993; Parry-Smith et al., 1998). Elements of secondary structure in LrpA are shown as labeled cylinders (helices) and arrows (strands). [The figure was produced using CINEMA (Parry-Smith et al., 1998) and ALSCRIPT (Barton, 1993)]. Residues conserved across all three sequences (Escherichia coli Lrp, Escherichia coli AsnC, and Pyrococcus furiosus LrpA) have been boxed. Those residues that are conserved between Lrp and AsnC are shaded red and the residues switched from a hydrophobic side chain in Lrp to a hydrophilic side chain in AsnC are shaded blue.
exists as a dimer in solution and binds to double stranded DNA in a homodimer conformation state (Wang and Calvo, 1993).
A compilation of operons has been confirmed to be regulated by Lrp protein directly or indirectly (Cho et al., 2008). However, this compilation is devoid of operons whose expressed poly- peptides are yet to be confirmed. In some cases, Lrp dependent activation or repression is not affected by the presence or absence of leucine (Ettema et al., 2002). This amino acid influences the oligomeric state of Lrp, with the dimer-dimer interaction being favored in its presence (Chen et al., 2001, 2005; de los Rios and Perona, 2007; Peterson et al., 2007).
In the present study, we purified the 6 × His tagged-Lrp and prepared 3H-labeled Lrp to perform the crosslink experiments to analyze the quaternary structure of Lrp, especially under the micromolarity concentration of Lrp. E. coli XL 1 blue competent cell and BL21 expressing strains were purchased from RBC Bioscience Co.; gel extraction kits were obtained from Solgent Company. Bronsted Lowry protein concentration determination reagents and protein markers were purchased from BioRad Co.
The cross linker of glutaraldehyde and 1, 2, 3, 4-diepoxy- butane (DEB) were obtained from Sigma, and ethylene glycol
bis (succinimidyl succinate) (EGS) was purchased from Pierce Chemical Co.
Single colonies from recombinant E. coli strains transformed with wild type gene of Lrp inserted in pQE 30 vector were cultured in 2 L of terrific broth media (tryptone 12 g, yeast- extract 24 g, 4 ml glucose, 2.31 g KH2PO4, and 12.54 g K2HPO4) containing ampicillin (100 μg/ml) at 37°C in shaking volumetric flasks until the optical density of 0.7 was recorded at the wavelength of 600 nm. Isopropyl-β-D-thiogalactopyranoside (IPTG) was added to the concentration of 1 mM. This was followed with incubation at 25°C for 10 h.
The construction of the wild type (pCV 294) genes coding for the 6 × His-tagged wild type Lrp was described in our previous paper (Lee et al., 2010). Frozen cells of mass 3 g, obtained from the culture, were thawed 30 min on ice and then resuspended in 25 ml of lysis buffer (50 mM NaH2PO4, 300 mM NaCl, 10 mM imidazole at pH 8.0). The detailed condi- tions for the expression and purifications of proteins were the same as reported previously (Lee et al., 2010; Pokoo et al., 2014).
A high protein expression of molecular weight 18 kDa that
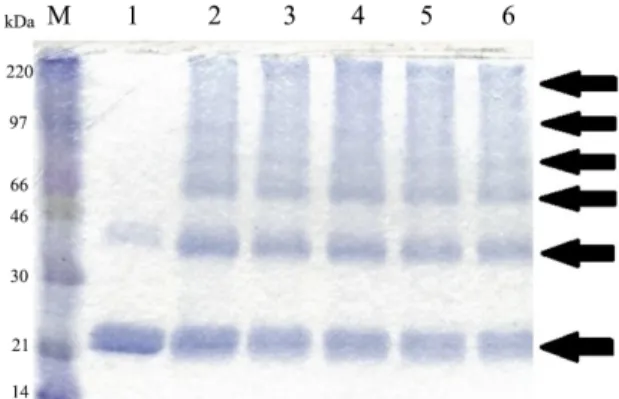
Fig. 2. SDS-PAGE analysis of Lrp protein after the final step of purification.
Lanes: 1, Protein size marker (15, 20, 30, 40, 50 kDa); 2~4 eluted fractions of 0.1 M imidazole; 5~9, eluted fractions of 0.25 M imidazole. The flow through fractions were collected and later analyzed on SDS-PAGE. The column was then washed with lysis buffer and wash buffer (50 mM NaH2PO4, 10% glycerol, and 300 mM NaCl at pH 6.0), respectively. The purified protein was eluted with an elution buffer (50 mM NaH2PO4, 300 mM NaCl, 250 mM imidazole at pH 8.0). Collected eluate fractions were each analyzed on 12.5% SDS-gel to confirm the molecular weight of the protein and their level of purity. The fractions containing purified protein of interest were pooled together and concentrated. The purified protein was then dialyzed in Lrp protein storage buffer (10 mM Tris base, 50%
glycerol, 0.1 mM EDTA, 0.1 mM DTT, and 0.1 M NaCl).
Fig. 3. Oligomeric state of Lrp protein by forming of cross-linking experi- ment of with increasing concentrations of glutaraldehyde. Lanes: M, protein size marker (14, 21, 30, 46, 66, 97, 220 kDa); 1, no glutaraldehyde; 2, 2 mM glutaraldehyde; 3, 4 mM glutaraldehyde; 4, 6 mM glutaraldehyde; 5, 8 mM glutaraldehyde; 6, 10 mM glutaraldehyde. The purified Lrp protein of concentration 8 μM was pre-incubated in a reaction solution containing HEPES and 1.5 mM MnSO4 for 10 min. Afterwards, the resultant solution was incubated with glutaraldehyde for 20 min. This was finally followed by a 10-min incubation in 1 M Tris buffer. Each incubated sample was run on 12% SDS-PAGE.
corresponds to the calculated value of Lrp was identified on 12% SDS-PAGE. It was confirmed that there was no protein expressed by all genes in the absence of IPTG (data not shown).
The recombinant proteins obtained from the supernatant fraction were higher than those in the pellet after centrifugation was performed several minutes after sonication. Hence, a large culture of 2 L of terrific broth media was inoculated with an overnight culture at 37°C. Lysis of the cells and purification of the proteins from the culture yielded appreciably a high con- centration of protein (see Fig. 2).
In order to prepare a lower concentration of Lrp, we prepared
3H labeled 6 ×-His tagged Lrp, uniformly labelled with tritium leucine. L-leucine amino acid has been identified as the most abundant and readily available building protein (Hung et al., 2002; Brinkman et al., 2003). Tritium labeled Lrp was prepared as follows: 2 ml of a stationary phase culture of strain CV1509 (leu-83::Tn10) (Landgraf et al., 1996) were inoculated into 100 ml of M9 minimal medium containing thiamine, ampicillin, kanamycin, tetracycline, and 5 μg/ml L-[4,5-3H] leucine (134 μCi/μmole; Amersham).
The culture was induced with IPTG at the A600 of 0.8 and, after further incubation for 5 h, tritium labeled Lrp was isolated by the same method as for Lrp purification described above.
Only a single radioactive band was observed by SDS-PAGE
that corresponded in size to Lrp (data not shown). The specific activity of Lrp was calculated as the specific activity of leucine in the medium leucines/monomer Lrp. The specific activity of the protein was also determined experimentally by adding known amounts of the tritium protein (Bio-Rad protein assay) to the 0.35 cm3 slice of 6% polyacrylamide and counting as described previously (Cui et al., 1996).
Initially, we checked the quaternary structure of Lrp by performing crosslink experiment with glutaraldehyde which is space arm of 0 Å. It was observed that the Lrp protein formed high oligomeric forms according to the increasing concentrations of glutaraldehyde in 12% SDS-PAGE (Fig. 3). The protein sample that did not contain glutaraldehyde cross linker showed a visibly strong band that corresponded to the Lrp monomeric state. Also, a visibly low band was observed at the position that corresponds to the dimer conformation (see Lane 1 in Fig. 3).
This is because the concentration of the 8 μM Lrp protein used in the cross-linking experiment is almost equal 10 μM at which Lrp forms a dimer (Wang and Calvo, 1993). From 2 mM of glutaraldehyde, in addition to dimeric proteins the bands corresponding to tetrameric, hexameric, even in octameric conformations were shown at those positions (see Lanes 2~6 in Fig. 3). However, the bands of hexameric and octameric proteins
(A) (B)
(C) (D)
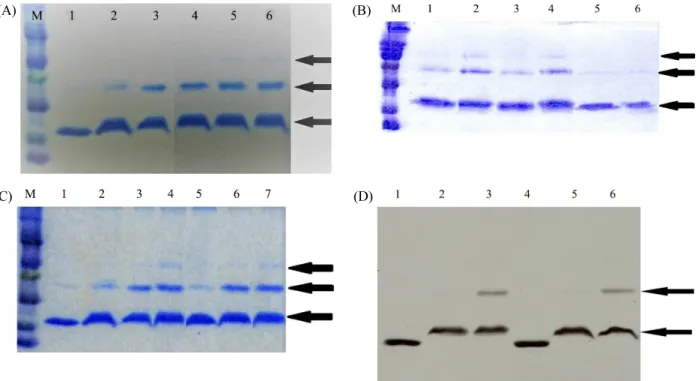
Fig. 4. Crosslink experiment of Lrp with 1,2,3,4-diepoxy-butane (DEB). (A) Effect of DEB concentration on the forming of multimeric Lrp. The reaction solution consisted of 10X TMN buffer, Lrp of 4.5 μM was incubated for 5 min after the addition of DEB in 50 mM tri-ethanolamine at 37°C. Afterwards, 1 M Tris of pH 7.6 was added to stop incubation for the period of 30 min and the resultant mixture was mixed with the sample buffer and together incubated for 2 min. Lane M, protein size marker (14, 21, 30, 46, 66, 97, 220 kDa, respectively). The concentration of DEB was as follows: Lane 1, no DEB; Lane 2, 8 mM; Lane 3, 16 mM; Lane 4, 32 mM; Lane 5, 80 mM; and Lane 6, 168 mM. The arrows indicate the conformers of Lrp. (B) Comparison of the formation of multimeric Lrp in the absence and presence of DTT. Crosslink experiments were analyzed by Coomasie blue-stained 12% SDS-PAGE. Lane M, protein size marker (14, 21, 30, 46, 66, 97, 220 kDa); Lanes 1~4, 4.5 μM Lrp adding 80 mM DEB in the absence or presence of 2 mM DTT in reaction mixture; Lanes 5 and 6, Lrp without DEB in the absence or presence of DTT. The concentration of DEB in lanes 1~6 were 80 mM DEB. 2 mM DTT were added in Lanes 2, 4 and 6. (C) Comparison of the formation of multimeric Lrp in the absence and presence of DNA. Lane M, marker (14, 21, 30, 46, 66, 97, 220 kDa); Lane 1, 9 μM Lrp itself, Lanes 2~4 Lrp added DEB without DNA, Lanes 5~7 Lrp added DEB with 9 μM DNA. Concentrations of DEB are 52 mM in Lanes 2 and 5, 210 mM in Lanes 3 and 6, 420 mM in Lanes 4 and 7, respectively. (D) Fluorography of the 3H-labeled Lrp in the crosslink experiments. Formation of dimeric 3H labelled Lrp in the absence DNA are shown in Lanes 1–3 and the presence of was 0.30 μM DNA in Lanes 4–6. The concentration of 3H Lrp was 0.28 μM. Lanes 1 and 4, 3H Lrp without DEB; Lanes 2 and 5, adding 8 mM DEB; Lanes 5 and 6, adding 32 mM DEB.
appeared to be smeared due to the low degree of space arm in the glutaraldehyde cross linker.
To get a more precise quaternary structure of Lrp, we per- formed cross link experiments with DEB (1, 2, 3, 4-diepoxy- butane), a cross linker with space arm of length 4 Å. As shown in Fig. 4A, the dimeric Lrp state was increased by adding DEB.
At a high DEB concentration, the band corresponding to the tetrameric state shown in Lanes 5 and 6 in Fig. 4A was observed.
There are two cysteine amino acid residues at the position of 45 and 106 in Lrp. Therefore, we checked the possibility of the formation by a disulfide bond between the two cysteines.
Dithiothreitol (DTT) was added to dissolve the disulfide bonds that can be formed in intra-subunit or inter-subunits of Lrp. The reagent prevents any form of natural oligomeric conformation
that could be formed as a result of interactions between cysteine residues within the Lrp.
As shown by the crosslink experiments (see Fig. 4B), the reaction mixture containing Lrp and sample buffer were incu- bated together by adding DEB. The cross linking solutions were prepared in the absence and presence of DTT. The incubated samples at Lanes 5 and 6 in Fig. 4B that did not contain the cross linker DEB only showed the monomer con- formation. This signifies that, in the absence of the cross linker and presence of dithiothreitol, the Lrp remains in its monomer state (see Fig. 4B).
The samples at Lanes 2 and 4 which contains DTT showed a high oligomeric conformation of dimer and tetramer, respec- tively. Especially, the intensity of the bands corresponding
tetrameric state are shown to have a much high intensity as compared to those from the samples in the absence of DTT in Lanes 1 and 3. This result indicates that the preventing of disulfide bond from the two cysteine residues by adding DTT may be helpful for the formation of tetrameric Lrp confomer by the cross linker of DEB. From these results, it can be inferred that Lrp could form high oligomeric conformations dependent on the interactions between the cysteine residues present in its amino acid sequence.
In previously reported data (Cui et al., 1996), the purified 6 × His-tagged Lrp which contains 11 amino acid extended at the N-terminus did not appear to have major negative effects on the function that binds to six binding sites upstream of the ilvIH promoter without the same affinity as does Lrp. The two com- plementary DNA (5ʹ-AAGGAGAATATTATGCTATGG-3ʹ and 5ʹ-CCATAGCATAATATTCTCCTT-3ʹ) were designed from the specific Lrp binding DNA sequences from ilvIH operon; furthermore, it was confirmed that the protein very strongly binds the double strand DNA by the gel mobility shift assay (Lee et al., 2010).
Two HPLC purified complementary oligonucleotides (21-mer DNA obtained from Bioneer Co.) were annealed in the TE buffer to synthesize double stranded 80 μM DNA. Lrp were bound together with the complementary double stranded DNA formed by thermal annealing. We checked the cross-linking experiments in the presence of the DNA. As shown in Fig. 4C, there were no differences in the formation of dimeric and tetrameric confomers of Lrp by DEB in the absence (Lane 2~4) and presence (Lanes 5~7) of the DNA.
This result can be attributed to the fact that the DNA binding site of Lrp and oligomerization site of Lrp are different, although the DNA binds tightly to the Lrp (Brinkman et al., 2003). The N-terminal domain, referred to Helix-Turn-Helix domain (Hart et al., 2011), is the DNA binding unit of the Lrp protein, a middle domain responsible for the transcription of activation, a C-terminal domain of Lrp consists of a fold similar to that of RAM (regulation of amino acid metabolism) domain (Calvo and Matthews, 1994). In additions, studies of C-terminal deletion mutant suggest that the C-terminal domain of Lrp is involved in dimerization and oligomerization of dimer (Chen and Calvo, 2002).
In order to check the conformer of Lrp below micromolar
concentrations, we prepared the tritium leucine labeled Lrp described above. The protein concentration of below 0.3 μM of Lrp was used. As shown in Fig. 4D, the monomer and dimer of Lrp were clearly shown in the SDS-PAGE. At lower Lrp concentration, the patterns of crosslink experiments in the presence or absence of DNA were the same as in the high Lrp experiment shown in Fig. 4C. Interestingly, we observed that the Lrp monomer state without DEB has a high mobility, similar to that of Lrp monomer state in the presence of Lrp in Fig. 4A~4D. It can be explained that the conformation state of monomer Lrp is differs from the Lrp one with DEB cross linker.
We also tested the oligomeric state of Lrp by experiment with the bi-functional crosslinking agent of ethylene glycol bis (succinimidyl succinate) (EGS) which is space arm of 6 Å. In this experiment, about 7.5 mg/ml of desalted Lrp was mixed with HEPES buffer, increasing concentrations of EGS dissolved in 20 mM dimethyl sulfoxide (DMSO). The resultant mixture was incubated at room temperature for 1 h after which the incubation reaction was quenched using 1 M Tris for about 30 min. The incubated samples were then analyzed on 12.5%
SDS-PAGE to identify the oligomers formed.
It was observed that distinct and high oligomeric confor- mations were formed by the samples located in lanes 2, 3, and 4 in the 12% destained gel (see Fig. 5A). Samples in Lane 1, however, showed a visibly clear band that represents the monomeric conformation at 21 kDa and a less clear at 42 kDa that represents the dimeric form (see Fig. 5A). The reason attributed to this feature is that, incubated sample 1 did not contain the cross linker EGS and, hence, the extent of cross- linking interactions to form higher oligomeric forms are very low.
However, it was also observed from Fig. 5A that, as the concentration of the EGS cross linker increased, the number of oligomeric forms formed by the incubated samples increased correspondingly. This can be clearly identified from Lane 1, where the sample did not contain the cross linker up to lane 4, where the sample had the highest cross linker concentration. In conclusion, cross linking experiments with EGS show that the wild type protein of Lrp tends to be associated to a protein dimer-dimer interaction and forms higher oligomeric confor- mations.
In order to check the conformer of Lrp at a lower con-
(A)
(B)
(C)
Fig. 5. Crosslink experiments of Lrp with ethylene glycol bis (succinimidyl succinate) (EGS). (A) 12 μM Lrp incubated with EGS. Lane 1, absence of EGS; Lane 2, 0.4 mM of EGS; Lane 3, 0.6 mM of EGS; Lane 4, 0.8 mM of EGS; Lane M, protein size marker (14, 21, 30, 46, 66, 97, 220 kDa). The 12 μM concentration Lrp was incubated in a reaction solution consisting of HEPES and EGS of concentrations 0.4, 0.6, 0.8 mM, respectively at room temperature for 1 h. The reaction mixture was then quenched for 30 minutes with a 1 M Tris buffer. About 5 μl of each sample were run on 12% SDS-PAGE to determine the oligomeric states formed due to cross-linking with EGS. (B) Crosslink experiment of 3H Lrp with EGS.
Lrp concentration was 0.28 μM. Concentrations of EGS were as follows;
Lane 1, 0; Lane 2, 0.1 mM; Lane 3, 0.2 mM; Lane 4, 0.3 mM; Lane 5, 0.4 mM; Lane 6, 0.6 mM. (C) Cross-linking of 3H Lrp with EGS in the absence and presence of DNA. Lane 1, 0.28 μM 3H Lrp; Lane 2, Lrp with 0.04 mM EGS; Lane 3, Lrp with 0.4 mM EGS; Lane 4, Lrp with 0.04 mM EGS plus 0.6 μM DNA; Lane 5, Lrp with 0.4 mM EGS plus 0.6 μM DNA. The positions of migration of standards are marked on the Lane M (14, 21, 30, 46, 66, 97 kDa, respectively).
centration lower to micromolar concentrations, we used the tritium leucine labeled Lrp described above. The protein concentration of below 0.3 μM of Lrp was used. As shown in Fig. 5B, the monomeric and dimeric states of Lrp were clearly shown in the SDS-PAGE. As shown in Fig. 5B, the dimeric Lrp
state increased by adding EGS. At a high EGS concentration, higher bands corresponding to the dimeric state were observed (Lanes 5 and 6 in Fig. 5B).
To check the conformer of low concentrations Lrp in the presence of the doubled stranded DNA, we also tested crosslink experiments with 3H Lrp below 0.3 μM with EGS in the absence and presence of DNA. As shown in Fig. 5C, the monomer and dimer of Lrp were clearly shown in the SDS- PAGE. At a lower Lrp concentration, the pattern of crosslink experiments in the absence or presence of DNA are different shown in Lanes 3 and 5 in Fig. 5C. 3H Lrp crosslink experi- ments with 0.4 mM EGS plus 0.6 μM DNA show the tetrameric state and the octameric state of Lrp show in Lane 5 in Fig. 5C.
These results are different from the pattern obtained the cross- linking with DEB in Fig. 4D and may be due to the fact that the EGS is a bifunctional cross linker and its space arm is wider than the cross linker of DEB.
In conclusion, crosslink experiments show that low or high concentrations Lrp tend to be associated to a protein dimeric state and form higher oligomeric conformations with various cross linkers.
적 요
루신-대응 조절 단백질(Lrp)은 18.8 kDa의 분자량을 갖는 글 로벌 조절 단백질로서 대장균과 같은 장내세균과에서 많은 대 사작용 오페론의 기능적 활성도를 조절한다. 단백질의 4차 구 조를 규명하기 위한 목적으로 Lrp 단백질 코드하는 유전자가 삽 입된 재조합 플라스미드 pQE vector를 발현시킨 6 × His-tag Lrp 야생형과 3H로 표지된 Lrp를 분리・정제한 후 cross linker들인 glutaraldehyde, 1,2,3,4-diepoxy-butane (DEB), ethylene glycol bis (succinimidyl succinate) (EGS)으로 cross link 실험을 수행 하여 Lrp가 0.3 μM 이하의 낮은 농도에서나 5 μM의 높은 농도 에서 이량체, 사량체, 육량체, 팔량체로 존재할 수 있음을 확인 하였다.
Acknowledgements
This work was supported by the research grant from Chungnam National University.
References
Barton, G.J. 1993. ALSCRIPT: a tool to format multiple sequence alignments. Protein Eng. 6, 37–40.
Brinkman, A.B., Ettema, G.J.T., de Vos, W.M., and Van der Oost, J.
2003. The Lrp family of transcriptional regulators. Mol. Microbiol.
48, 287–294.
Calvo, J.M. and Matthews, R.G. 1994. The leucine-responsive regulatory protein, a global regulator of metabolism in Escherichia coli.
Microbiol. Rev. 58, 466–490.
Chen, S. and Calvo, J.M. 2002. Leucine-induced dissociation of Escherichia coli Lrp hexadecamers to octamers. J. Mol. Biol.
318, 1031–1042.
Chen, S., Rosner, M.H., and Calvo, J.M. 2001. Leucine-regulated self-association of leucine-responsive regulatory protein (Lrp) from Escherichia coli. J. Mol. Biol. 312, 625–635.
Chen, S., Iannolo, M., and Calvo, J.M. 2005. Cooperative binding of the leucine-responsive regulatory protein (Lrp) to DNA. J. Mol.
Biol. 345, 251–264.
Cui, Y., Midkiff, M.A., Wang, Q., and Calvo, J.M. 1996. The leucine responsive regulatory protein (Lrp) from Escherichia coli.
Stoichiometry and minimal requirements for binding to DNA. J.
Biol. Chem. 271, 6611–6617.
Cho, B.K., Barett, C.L., Knight, E.M., Park, Y.S., and Palasson, B.O.
2008. Genome scale reconstruction of the Lrp regulatory network in Escherichia coli. Proc. Natl. Acad. Sci. USA 105, 19462– 19467.
Deng, W., Wang, H., and Xie, J. 2011. Regulatory and pathogenesis roles of Mycobacterium Lrp/AsnC family transcriptional factors.
J. Cell Biochem. 112, 2655–2662.
de los Rios, S. and Perona, J.J. 2007. Structure of the Escherichia coli leucine-responsive regulatory protein (Lrp) reveals a novel octameric assembly. J. Mol. Biol. 366, 1589–1602.
Ernsting, B.R., Atkinson, M.R., Matthews, R.G., and Ninfa, A.J.
1992. Characterization of the regulon controlled by the leucine responsive regulatory protein in Escherichia coli. J. Bacteriol.
174, 1109–1118.
Ettema, T.J.G., Brinkman, A.B., Tani, T.H., Rafferty, J.B., and van der Oost, J. 2002. A novel ligand-binding domain involved in regulation of amino acid metabolism in prokaryotes. J. Biol.
Chem. 277, 37464–37468.
Hart, B.R. and Blumenthal, R.M. 2011. Unexpected coregulatory range for the global regulator Lrp of Escherichia coli and Proteus mirabillis. J. Bacteriol. 193, 1054–1064.
Hart, B.R., Mishra, P.K., Listner, R.E., Hinerman, J.M., Herr, A.B., and Blumenthal, R.M. 2011. Recognition of DNA by the helix-turn-helix global regulatory protein Lrp is modulated by the amino-terminus. J. Bacteriol. 193, 3794–3803.
Hung, S., Pierre, B., and Wesley, H.G. 2002. Global gene expression profiling in Escherichia coli K-12. The effects of leucine- responsive regulatory protein. J. Biol. Chem. 277, 40309–40323.
Landgraf, J.R., Wu, J., and Calvo, J.M. 1996. Effects of nutrition and growth rate on Lrp levels in Escherichia coli. J. Bacteriol. 178, 6930–6936.
Lee, C.Y., Kim, S.C., and Seo, C.H. 2010. Purification and fluorometric analysis of leucine-responsive regulatory protein from Escherichia coli. Korean J. Microbiol. 46, 104–108.
Newman, E.B., D’Ari, R., and Lin, R.T. 1992. The leucine-Lrp regulon in Escherichia coli: A global response in search of a raison d'etre.
Cell 68, 618–620.
Newman, E.B, Lin, R.T., and D’Ari, R. 1996. The leucine/Lrp regulon.
Annu. Rev. Microbiol. 49, 1513–1525.
Parry-Smith, D.J., Payne, A.W.R., Mitchie, A.D., and Attwood, T.K.
1998. CINEMA—a novel colour interactive editor for multiple alignments. Gene 221, 57–63.
Peterson, S.N., Dahlquist, F.W., and Reich, N.O. 2007. The role of high affinity non-specific DNA binding by Lrp in transcriptional regulation and DNA organization. J. Mol. Biol. 369, 1307–1317.
Philip, M.L., Sander, H.J.S., Svetlana, E.S., Arie, B.B., Willem, M.D., Van der Oost, J., David, W.R., and John, B.R. 2001. Crystal structure of the Lrp-like transcriptional regulator from the archaeon Pyrococcus furiosus. EMBO J. 20, 990–997.
Pokoo, R., Lee, E.H., and Lee, C.Y. 2014. Fluorescence characteristics of a tryptophan mutant of leucine-responsive regulatory protein.
Korean J. Microbiol. 50, 275–280.
Shimada, T., Saito, N., Maeda, M., Tanaka, K., and Ishihama, A. 2015.
Expanded roles of leucine responsive regulatory protein in transcription regulation of the Escherichia coli gemome: Genomic SELEX screening regulation targets. Microb. Genom. 1, e000001.
Wang, Q. and Calvo, J.M. 1993. Lrp, a global regulatory protein of Escherichia coli, binds cooperatively to multiple sites and activates transcription of ilvIH. J. Mol. Biol. 229, 306–318.