Characterization of Korean Wheat Line with Long Spike. I. Agronomic Traits and Genetic Variations
10
0
0
전체 글
(2) 220. 韓育誌(Korean J. Breed. Sci.) 47(3), 2015. 소하고 있다(Kang et al. 2014). 재배 농가의 요구에 부응하. 기거나 늦출 수 있다. Vrn-1 과 Ppd-1 등은 각각 파성과 일장. 는 수량성 높은 밀 품종 개발을 위하여 금강밀과 중국밀 자원. 과 관련되어 있는 유전자로서 각각 3개의 상동유전자를 포함. 인 Xian83의 교배를 통해 이삭 길이와 1수립수가 많은 장수. 하고, 5번과 2번 염색체에 위치하고 있다(Kamran et al.. 형 계통을 개발하였다(Kang et al. 2013).. 2014). Vrn-1은 개화 시기와 식물의 키 등에 영향을 미치고. Atsmon과 Jacobs (1977)가 개발한 “Gigas”는 최대 30개. (Stelmakh 1998), Ppd-1는 일장 둔감형 유전자로서 일장에. 의 이삭을 갖는 장수형 밀로 종자 1개의 무게는 63 mg이고. 민감한 유전자보다 개화 시기를 앞당기기 때문에 여름의 고. 한 소수당 9개의 종자가 열린다. 그러나 Gigas는 분얼이 왕성. 온과 건조 조건에서도 수량을 증대시킬 수 있는 중요한 유전. 하지 않고 억제되는 경향을 나타냈는데, 이것은 염색체 1AS. 자이다(Snape et al. 2001).. 에 위치하고 있는 tin 열성유전자가 원인으로 알려져 있다. 국내 품종들에 대한 품질 관련 유전자와 같이 농업형질과. (Richards 1988, Spielmeyer & Richards 2004). tin은 수량. 관련된 TaCwi-A1, TaGW2, TaSux2-2B, Rht, Ppd-1, Vrn-1. 에 영향을 미치는 유전자로서 1수립수는 9% 정도 증가시키. 등 유전자의 연구가 이루어졌지만, 새로 개발된 장수형 밀. 지만, 수수와 립중을 각각 11%와 2% 감소시킨다고 보고되어. 164계통에 대한 연구는 아직 없다. 따라서 다수성 밀 품종 개. 있다(Duggan et al. 2005). 장수형 밀은 국제밀옥수수연구소. 발을 위한 장수형 밀의 농업적인 분석과 유전적인 구성요소. (CIMMYT)의 밀과 비교했을 때, 1수영화수, 1수립수, 개개. 를 함께 분석하여 한국의 밀 육종에서 다수성 밀 품종개발을. 의 립중이 각각 4%, 5%, 10%가 증가하는 것으로 보고되었. 위한 중요한 자원으로 활용하고자 하였다.. 지만, 대비품종인 Bacanora에 비해 수량이 8% 감소하고, 수. 재료 및 방법. 수가 26% 감소하는 것으로 확인 되었다(Gaju et al. 2009, 2014). 수수 및 1수립수 등 수량성과 관련된 유전자 및 다양한 분. 공시재료. 자마커 개발은 밀의 육종에 매우 유용하게 사용될 수 있다. 2005년도에 숙기는 늦지만 이삭이 길고 립수가 많은. (Bernardo 2008). 종실의 무게와 크기는 수량에 직접적으로. CIMMYT의 수집종인 Xian83을 모본으로 하고, 숙기가 빠르. 관련되어 있기 때문에 매우 중요하며 이와 관련된 유전자로. 고 다목적용 밀 품종인 금강밀을 부본으로 인공교배하여. Sucrose synthase 2 (Sus2), grain width (GW2)와 cell-wall. IW20050259 조합을 육성하였다. 국립식량과학원 시험포장. invertase (CWI) 등 3개가 보고되어 있다(Liu et al. 2012).. (전북 익산시)에서 교잡종(F1)의 분리세대(F2) 종자를 국립식. 각각의 유전자형에 따라 TaSus2-2B의 Hap-H과 TaGW2-6A. 량과학원 운봉출장소(전북 남원시)에서 일반적인 집단육종법. 의 Hap-6A-A는 천립중 증가와 매우 밀접하게 연관되어 있는. 에 의해 내한성이 양호한 계통을 선발하였다. 잡종 제3세대. 것으로 보고되어 있다(Jiang et al. 2011, Su et al. 2011). 또. (F3)이후에는 국립식량과학원 시험포장에 집단 재배하여 이. 한 Ma et al. (2012)은 TaCwi-A1의 분석을 통해 TaCwi-A1a. 삭이 길고 립수가 많은 계통을 선발하였다. 본 연구에는 잡종. 대립유전자의 천립중은 TaCwi-A1b 보다 많은 것을 확인하였. 제 8세대(F8) 164계통을 공시하였다. 부본으로 이용된 Xian83. 다. 반왜성 유전자(Rht)는 녹색혁명을 이끈 중요한 유전자로. 은 CIMMYT에서 중국자원에 대한 붉은곰팡이병 평가를 위. 서 서로 다른 대립유전자는 식물의 길이 생장에 서로 다른 영. 해 도입되었지만, 평가 이후 폐기되어 농업형질 및 관련 표지. 향을 미친다(Börner et al. 1996). 특히 밀의 염색체 4B와 4D. 인자와의 비교는 부본인 금강밀과 비교했다.. 에 위치하고 있는 Rht-B1b과 Rht-D1b는 DELLA 단백질을 암호화하고 있고 지베렐린산(GA3)에 둔감한 유전자이다. 농업형질 조사. (Tosovic-Maric et al. 2008). 이들 반왜성 유전자는 간장을. 농업 형질 조사는 농촌진흥청 농업 과학 기술 조사 분석 기. 감소시키고 고온 등 기후 변화 아래에서도 밀의 수량을 증대. 준(RDA 2012)에 준하여 실시하였다. 출수기는 전체 경수 중. 를 기대할 수 있게 하였다(Flintham et al. 1997). 파성. 에 40%가 출수한 날로 하였으며, 성숙기는 대부분의 이삭이. (vernalization)과 일장(photoperiod)은 밀의 개화를 조절하는. 황화한 날로 조사하였다. 간장은 지면에서 이삭 목까지의 길. 주요한 인자이다. 이들은 생장온도와 반응을 통해 개화를 시. 이로 나타내었고, 수장은 이삭 목에서 이삭 끝까지의 길이를. 작하는 것과 다른 환경적인 자극을 통해 개화의 시기를 앞당. 측정하였고, 단위 면적당 수수는 생육이 고른 3개소에서 25.
(3) 221. 장수형 밀의 계통 특성 분석 I. 농업 형질과 관련 유전자 특성 2. cm × 50 cm에 있는 총 수수를 측정하여 1 m 당 경수로 환산. 통계 분석. 하였다. 1수립수는 생육이 균일한 장소에서 무작위로 20수를. 모든 시험은 최소 3회 이상 반복을 두어 실시하였으며, 시. 채취하여 탈곡 후 조사하였다. 리터중은 탈곡한 종자를 2.0. 험성적은 SAS 프로그램(SAS Institute, Cary, NC)을 이용하. mm 종목체를 통과하지 않은 종자를 리터중 측정기를 이용하. 여 분산분석을 실시하고 LSD로 유의성을 검정하였으며, 품종. 여 측정하였으며, 천립중도 탈곡한 종자를 2.0 mm 종목체를. 과 연차간 변이의 상호작용을 보기 위한 ANOVA (Analysis. 통과하지 않은 종자를 1,000립을 측정하여 무게를 조사하였. of variance) 분석은 GLM을 이용하였으며, 품종과 연차 변. 다. 수량은 수확 탈곡 건조 후(수분 14% 기준) 설립을 제외하. 이의 상호작용 성분은 오차항을 이용하였다. 상관관계는 피어. 고 종실을 평량하여 10a당으로 환산하여 조사하였다.. 슨의 상관 계수를 이용하였으며, 통계적 유의 수준은 P<0.05 으로 실시하였다.. Genomic DNA 추출. 결과 및 고찰. 금강밀과 장수형 계통을 온실에 파종하여 3주 간 생육시켰 다. 유식물체의 엽을 채취하여 액체질소로 급속냉동 시킨 후 유발을 이용하여 미세분말로 분쇄한 뒤 -70°C에 보관하였다.. 장수형 계통의 농업 형질. Genomic DNA는 잎 분말(100 mg)로부터 식물용 Genomic. 장수형 계통의 잎색은 녹색이며 폭은 금강밀에 비해 넓고. DNA 분리시약(Solgent, Korea)을 이용하여 추출하였다.. 긴 편이다. 줄기 굵기는 중 정도이며 이삭은 방추형으로 까락 은 중간 정도이고, 종실 크기는 중 정도에서 대립크기의 분포. PCR 분석. 를 보였다(Fig. 1). 장수형 계통은 출수까지 평균 200.8일이. Rht-B1와 Rht-D1 유전자를 이용한 Polymerase chain. 걸렸고, 금강밀(193일)보다 약 7일 정도 많은 일수가 요구되. reaction (PCR)은 Ellis et al. (2002)의 방법으로 수행되었다.. 었다. 장수형의 간장은 88.3 cm로 금강밀보다 약 16 cm 더. Vrn-1 유전자 PCR 분석은 Yan et al. (2004), Fu et al.. 길었다(Table 1). 밀의 수량과 관련된 특징 중 이삭길이, 단위. (2005), Santra et al. (2009), Zhang et al. (2012), Chen et. 면적 당 수수, 1수립수, 리터중, 천립중과 생산량 등은 각각. al. (2013)의 연구방법에 따라 수행되었고, Ppd-1는 Beales et. 11.7 cm, 650.7개, 43.4개, 776.8 g, 44.6 g, 480.4 kg/10a으. al. (2007)과 Bentley et al. (2011)의 방법을 이용하였다.. 로 나타났다. 비교 품종인 금강밀은 7.0 cm, 711.1개, 27.5개,. PCR 생산물은 1.5% 아가로스젤에 전기영동하여 확인하였고,. 843.3 g, 45.1 g, 470.4 kg/10a였는데, 장수형 계통은 단위면. TaSus2-2B, TaGW2-6A, TaCwi-A1 등의 대립유전자 다양성. 적당 수수, 리터중은 금강밀보다 낮았지만 이삭의 길이가 길. 은 Jiang et al. (2011), Su et al. (2011)과 Ma et al. (2012). 고, 1수립수가 많아 전체적인 수량은 금강밀보다 약 10. 의 방법에 의해 분석되었다.. kg/10a 증수되는 것으로 나타났다(Table 1).. Table 1. Agronomic traits in Korean wheat line with long spike and Keumkang. Agronomic traits Days to heading date. Korean wheat population with long spike lengthz (n = 164) 200.8 ± 1.3. Keumkang. (197.3 – 204.0). 193.0. Culm length (cm). 88.3 ± 6.8. (66.1 – 102.3). 72.1. Spike length (cm). 11.7 ± 1.1. (9.1 – 14.4). 7.0. 2. Number of spike per m. Number of kernel per spike Test weight (g) 1000-kernel weight (g) Grain yield (kg/10a) z. 650.7 ± 117.0 (389.0 – 950.0) 43.4 ± 4.3. (30.8 – 54.7). 776.8 ± 16.9 (713.0 – 815.0) 44.6 ± 5.7. (31.6 – 55.9). 480.4 ± 74.7 (243.2 – 654.1). 711.1 27.5 843.3 45.1 470.4. Value indicates average and standard deviation of Korean wheat line with long spike and parenthesis includes ranges of Korean wheat line with long spike.
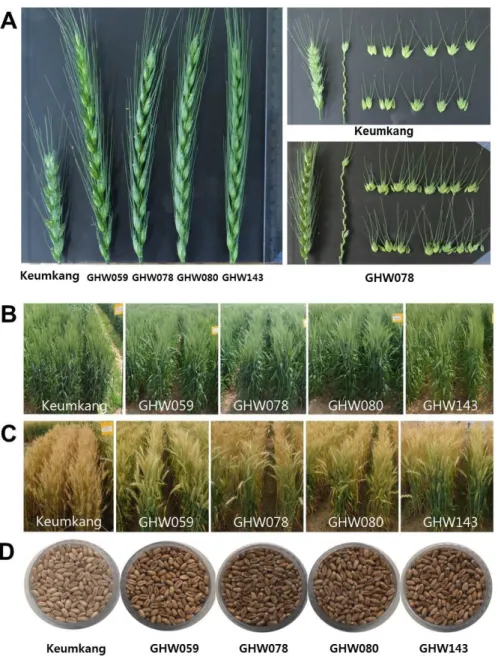
(4) 222. 韓育誌(Korean J. Breed. Sci.) 47(3), 2015. 장수형 계통의 농업적 형질 간의 상관관계 분석을 통해 간. 1수립수의 상관도 매우 높게 나타났다. 리터중은 천립중과 생. 장은 출수까지의 일수와 수장과 매우 높은 상관을 갖는 것으. 산량과 매우 높은 정의상관을 나타냈지만, 출수까지의 일수와. 로 나타났다. 이와 같은 결과는 장수형 계통이 금강보다 긴. 는 부의상관을 나타냈다. 뿐만 아니라 천립중과 생산량도 출. 간장과 수장을 갖고 있기 때문에 그 만큼 이들이 자랄 수 있. 수까지 일수와 고도의 부의상관을 나타냈다(Table 2).. 는 충분한 일장이 요구되기 때문인 것으로 생각된다. 단위면. 잎의 크기는 생산량과 바이오매스 생산에 매우 중요한 요. 적당 수수는 금강에 비해 50개가 적었지만, 1수립수와 전체. 소이다(Gonzalez et al. 2009). 잎의 크기와 관련된 생장은 세. 적인 수량은 금강보다 많았다. 장수형 계통의 종자 크기는 금. 포증식과 세포신장의 중첩된 두 단계로 나뉘어져 있고, 큰 잎은. 강밀과 비슷하고, 긴 이삭에 종자가 많이 형성되기 때문에 수. 세포의 크기와 관련이 있는 것으로 보고되어 있다(Anastasiou. 량이 많게 나온 것으로 생각된다(Fig. 1). 이와 같이 수장과. & Lenhard 2007). Shin et al. (2015)은 금강과 비교한 결과. Fig. 1. Image of spike and spikelet (A), heading stage (B), maturing stage (C) and grain (D) of Korean wheat 164 line with long spike and Keumkang..
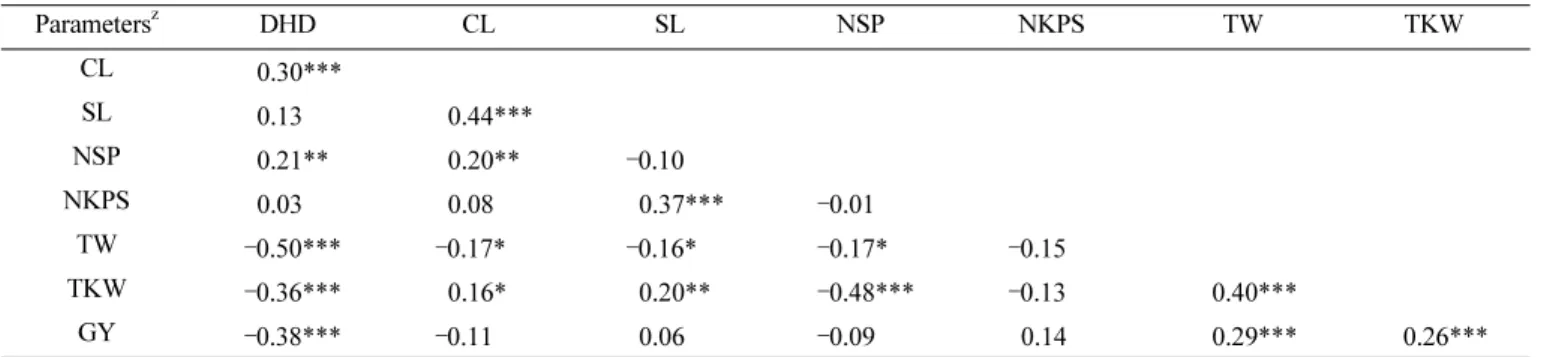
(5) 223. 장수형 밀의 계통 특성 분석 I. 농업 형질과 관련 유전자 특성 Table 2. Correlation coefficients among agronomic traits of Korean wheat line with long spike. z. Parameters. DHD. CL. SL. NSP. NKPS. TW. TKW. CL. 0.30***. . . . . . . SL. 0.13. 0.44***. . . . . . 0.21**. 0.20**. . . . . 0.03. 0.08. . . . NSP NKPS TW. –0.50***. –0.17*. TKW. –0.36***. 0.16*. –0.10 0.37*** –0.16* 0.20**. –0.01 –0.17*. –0.15. . . –0.48***. –0.13. 0.40***. . –0.38*** –0.11 0.06 –0.09 0.14 0.29*** 0.26*** z DHD = days to heading date, CL= culm length, SL= spike length, NSP= number of spike per m2, NKPS= number of kernel per spike, TW= test weight, TKW=1000-kernel weight, GY= grain yield y * indicates significance at 0.05 level, ** at 0.01 level, and *** at 0.001 level GY. 세포의 크기는 장수형 계통이 금강보다 30% 증가했지만 세. 두 Vrn-B1b, Ppd-A1b, Ppd-B1b를 갖고 있었으며, 각각. 포의 수나 광합성 효율은 차이가 없었고, 장수형 계통의 수장. 1,149 bp, 453 bp, 1,292 bp의 크기의 밴드로 증폭되었다. 은 금강보다 6.0 cm 이상 긴 것으로 보고하였다. 본 연구에서. (Fig. 2).. 수장과 1수립수와 고도의 정의상관을 나타내는 것을 알 수 있. Vrn-A1과 Vrn-D1 유전자는 각각 Vrn-A1a와 Vrn-A1b,. 다. 수장이 길수록 립수가 많고, 줄기는 길었지만 도복에 강한. Vrn-D1a와 Vrn-D1b의 유전자 형을 갖고 있었으며 1,068bp. 특성을 나타냈다. 장수형 계통의 줄기 두께는 5.02 cm로서. 와 522 bp의 크기로 증폭되었다(Fig. 2). Vrn-A1a는 금강을. 금강 4.35 cm보다 약 0.8 cm가 더 두껍기 때문에 많은 립수. 포함하여 156 계통에서 확인되었고, Vrn-A1b는 8개 계통에. 에도 불구하고 줄기가 수를 지탱하는 것으로 생각된다(Son et. 서 확인되었다. Vrn-D1은 금강과 157 계통에서 Vrn-D1b. al. 2014). 장수형 계통의 출수기는 금강보다 8일에서 10일. (997 bp), 7개 계통에서 Vrn-D1a (1,671 bp)가 확인되었다.. 정도 늦고, 개화기는 5일에서 6일 정도 늦다고 보고되어 있다. Ppd-D1 또한 Ppd-D1a (288 bp)과 Ppd-D1b (414 bp) 두 가. (Son et al. 2014). 리터중, 천립중과 생산량 등은 출수까지의. 지 유전자형을 갖고 있었고, 금강과 130개 계통이 Ppd-D1a,. 일수와 고도의 부의 상관을 나타내는 것은 출수기와 개화가. 34 계통이 Ppd-D1b를 갖고 있었다. 164 계통의 농업적 특성. 늦기 때문에 수확기까지 요구되는 충분한 시간이 우리나라. 과 비교해 보았을 때, Vrn-A1a와 Vrn-A1b는 유전자형에 따. 기후 여건에서 충족될 수 없는 환경적 영향에 의한 것으로 생. 른 특성차이에 유의성이 없었고, Vrn-D1은 출수기까지 일수,. 각되고, 이들 장수형 계통의 품종과 유전자원으로서 활용을. 수장, 1수립수에서 유의성의 차이가 나타났다. Vrn-D1이. 위해서는 숙기 단축이 매우 중요하게 요구될 것으로 판단된다.. Vrn-D1a를 갖는 계통이 출수까지 일수와 수장, 1수립수가 많 게 나타났다. 일장과 관련이 있는 Ppd-D1 유전자 분석에서. 밀의 농업적 특성과 관련된 유전자 분석. Ppd-D1a와 Ppd-D1b에 따라 출수까지 일수, 단위면적당 수. TaCwi-A1, TaGW2, TaSux2-2B, Rht-B1, Rht-D1, Vrn-A1,. 수, 1수립수, 리터중을 제외한 다른 특성에서 Ppd-D1a를 갖. Vrn-B1, Vrn-D1, Ppd-A1, Ppd-B1, Ppd-D1등 밀의 농업특. 는 계통이 많게 나타났다(Table 3). 파성과 일장 관련 유전자. 성과 관련이 있는 유전자 11개의 다양성을 분석하여 금강밀. 는 earliness per se (Eps) 유전자와 함께 밀의 환경에 대한 적. 과 비교하였다. PCR을 통해 증폭된 밴드는 167에서 1,671. 응을 조절하는 한 요인이다(Kato & Yamagata 1988). 이들. bp 범위 안에서 확인되었다(Fig. 2). Rht-B1, Rht-D1, Vrn-B1,. 유전자는 밀의 개화를 조절하는 것으로서 특정 환경에서 유. Ppd-A1, Ppd-B1 등 5개 유전자는 164개 장수형 계통에서 차. 전자형에 따라 개화 시기를 앞당기거나 늦추는 것을 조절한. 이가 없고 모두 같은 유전자형으로 나타났다. Rht-B1과 Rht-D1. 다(Worland 1996). 특히 파성은 보통밀에서 Vrn-A1, Vrn-B1,. 의 유전자형은 두 가지(237, 264bp)로서 장수형 계통은 Rht-. Vrn-D1, Vrn4, Vrn-B3 등 적어도 5개 대립유전자에 의해 조. B1b와 Rht-D1b (null-type) 금강밀은 Rht-B1a와 Rht-D1a로. 절되는 것으로 알려져 있고(Goncharov 2003, McIntosh et al.. 서로 다른 것을 확인하였다. 장수형 164 계통과 금강밀은 모. 1998, Pugsley 1971), 본 연구에서 Vrn-D1은 7개 계통에서.
(6) 224. 韓育誌(Korean J. Breed. Sci.) 47(3), 2015. Fig. 2. Agarose gel electrophoresis of PCR amplified Rht-B1 and Rht-D1 (A), Vrn-1 (B), Ppd-1 (C) and TaSus2-2B, TaGW2 and TaCwi-A1 alleles (D). M, molecular size marker; 1-12, Korean wheat line with long spike (GHW001~GHW012). Table 3. Difference in agronomic traits of Korean wheat line with long spike carrying different vernalization and photoperiod genotypes using pair-wise t-test. Photoperiod Ppd-D1 vrn-A1 Vrn-A1b vrn-D1 Vrn-D1a Ppd-D1a Ppd-D1b 156 8 7 157 130 34 No. Line y 200.8a 200.4a 200.7b 201.3a 200.5b 201.8a DHD 88.3a 90.0a 88.5a 85.7a 88.0a 89.5a CL (cm) 11.7a 11.8a 11.8a 10.8b 11.8a 11.5a SL (cm) 651.0a 645.8a 673.8a 649.7a 640.3b 690.5a NSP 43.4a 42.6a 43.6a 39.3b 43.7a 41.9b NKPS 777.2a 769.7a 776.3a 787.8a 778.5a 770.2b TW (g) 44.7a 42.4a 44.7a 42.7a 45.6a 40.5b TKW (g) 479.9a 490.1a 481.3a 460.0a 501.1a 401.1b GY (kg/10a) z DHD = days to heading date, CL= culm length, SL= spike length, NSP= number of spike per m2, NKPS= number of kernel per spike, TW= test weight, TKW=1000-kernel weight, GY= grain yield y Values followed by same letters within same characteristics at the same locus are not significantly different at P<0.05 Agronomic z Traits. Vernalization. Vrn-A1. Vrn-D1.
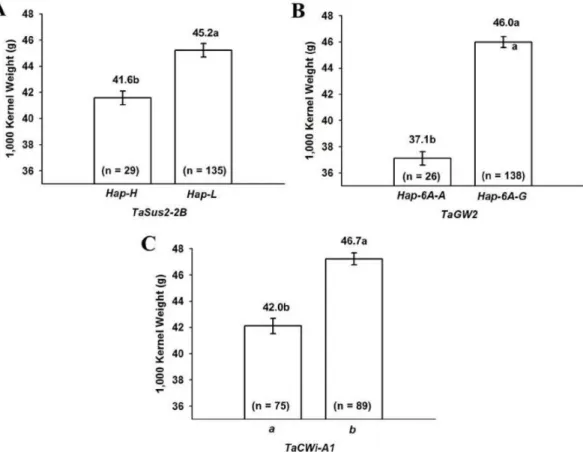
(7) 장수형 밀의 계통 특성 분석 I. 농업 형질과 관련 유전자 특성. 225. 확인되었고, 수장과 1수립수와 매우 깊게 연관되어 있는 것으. 비해 현저하게 높게 나타났다(Fig. 3). 본 연구를 통해 TaCwi-A1,. 로 확인되었다. Ppd-D1b를 가지는 34 계통은 출수기까지 일. TaGW2, TaSus-2B, Vrn-D1, Ppd-D1 등 5개 유전자는 밀의. 수와 수수가 많았지만 전체적인 수량은 Ppd-D1a보다 적었. 농업형질 확인을 위한 분자마커로 매우 유용하게 사용될 수. 다. Ppd-D1a는 1수립수, 리터중, 천립중, 생산량에서 모두 많. 있을 것으로 생각된다. TaCwi-A1 유전자는 저장조직 발달,. 은 것으로 확인되었기 때문에 Ppd-D1a는 수량에 직접적인. 탄소 구배, 종자무게와 깊게 관련되어 있는 효소로 알려져 있. 연관이 있는 것으로 생각된다.. 는 cell wall invertase (CWI)를 암호화 하고 있다(Miller &. TaCwi-A1 유전자 또한 두 가지 유전자형을 가지고 있다.. Chourey 1992, Sturm 1999, Tang et al. 1999). TaCwi-A1. 장수형 164계통 중 75 계통은 a형(402 bp), 89 계통은 b형. 은 밀의 염색체 2번에 위치하고 있으며 open reading frame. (404 bp)을 갖고 있었다. TaGW2 유전자는 Hap-6A-A와. (ORF)는 총 1,767 bp로서 7개의 엑손과 6개의 인트론으로 구. Hap-6A-G의 두 대립유전자를 갖고 있으며, Hap-6A-A는. 성되어 있다(Ma et al. 2012). TaSus2 유전자는 염색체 2A,. 167 bp, Hap-6A-G는 218 bp의 크기로 각각 확인되었다. 24. 2B, 2D에 위치하고 있으며 sucrose synthase 2 (SuS2)를 암. 계통에서 Hap-6A-A가 나타났고 나머지 계통에서 Hap-6A-G. 호화하고 있다(Jiang et al. 2011). TaSus2는 내배유에서 발. 로 나타났다. TsSus2-2B 유전자는 Hap-L과 Hap-H형의 유전. 현되는 유전자로서 유전형에 따라 농업적 형질이 다르게 나. 자형을 갖고 있고, 각각 318 bp와 423 bp의 밴드로 증폭되었. 타나는 것으로 알려져 있다. 이 유전자는 중국 밀 품종 육종. 다. Hap-L과 Hap-H의 두 유전자형은 164계통에서 각각 134. 단계에서 분자표지이용선발(MAS)을 위해 평가되었다(Jiang. 계통과 30 계통에서 확인되었다. Hap-L, TaCwi-A1b, Hap-6A-G. et al. 2011). TaGW2는 종자무게와 연관되어 있는 것으로 보. 등 3개의 유전자형을 갖는 계통의 천립중은 다른 유전자형에. 고되었고, 벼의 OsGw2와 상동성이 높은 것으로 발견되었다.. Fig. 3. Difference in 1,000-kernel weight according to the genotype of TaSus2-2B (A), TaGW2 (B) and TaCwi-A1 alleles (C) in Korean wheat line with long spike. Each bar represents the mean ± standard error. Bars with different letters above are significantly different at p<0.05..
(8) 226. 韓育誌(Korean J. Breed. Sci.) 47(3), 2015. TaGW2-6A의 유전자 코딩 지역의 염기서열 내에서는 변이가. (세부과제명: 용도별 초다수성 신초형 계통 육성, 세부과제번. 발견되지 않았고, 프로모터 지역에서 2개의 single nucleotide. 호: PJ009230012015에 의해 이루어진 것임.. polymorphisms (SNPs)가 발견되어 두 가지 유전자형을 갖게 된 것으로 보고되었다(Su et al. 2011). 본 연구 결과를 통해. REFERENCES. TaSus2는 천립중과 관련이 있는 것을 알 수 있었다. 이들 유 전자형과 농업형질이 깊은 연관이 있음이 확인되었지만 생산. 1. Beales J, Turner A, Griffiths S, Snape JW, Laurie DA.. 량과 관련된 여러 형질은 환경적인 영향도 많이 받기 때문에. 2007. A pseudo-response regulator is mis-expressed in. 장수형 계통에 대한 연구가 더욱 필요할 것으로 생각된다.. photoperiod insensitive Ppd-D1a mutant of wheat (Triticum. 적. 요. 국내 밀의 수량 증진을 위해 이삭 길이가 길고 1수립수가 많은 장수형 164계통(F8)을 육성하였고, 본 연구에서는 이들. aestivum). Theor. Appl. Genet. 115: 721-733. 2. Bentley AR, Turner AS, Gosman N, Leigh FJ, Maccaferri M, Dreisigacker S, Greenland A, Laurie DA. 2011. Frequency of photoperiod-insensitive Ppd-A1a alleles in tetraploid, hexaploid and synthetic hexaploid wheat. 계통의 주요 농업형질과 이들 특성과 관련 있는 표지인자에. germplasm. Plant Breed. 130: 10-15. 3. Bernardo R. 2008. Molecular markers and selection for. 대한 평가를 수행하였다. 장수형 계통은 모본인 금강밀과 비. complex traits in plants: Learning from the last 20 years.. 교했을 때, 단위면적당 수수와 리터중은 적고, 파종 후 출수까. Crop Sci. 48: 1649-1664. 4. Börner A, Plaschke J, Korzun V, Worland AJ. 1996. The. 지 일수가 길었고, 간장, 수장, 1수립수와 단위면적당 생산량 은 높게 나타났다. 농업 특성 관련 표지인자를 평가한 결과, 장수형 계통은 Rht-B1, Rht-D1, Vrn-B1, Ppd-A1, Ppd-B1 유전자 좌에서는 같은 유전자를 지닌 것으로 나타났다. 장수 형 계통 중 Vrn-D1를 지닌 계통은 Vrn-D1a를 지닌 계통에. relationships between the dwarfing genes of wheat and rye. Euphytica 89: 69-75. 5. Brancourt-Hulmel M, Doussinault G, Lecomte C, Berard P, LeBuanec B, Trottet M. 2003. Genetic improvement of. 비하여 출수에 걸리는 시간이 길었지만, 이삭길이와 1수립수. agronomic traits of winter wheat cultivars released in France from 1946 to 1992. Crop Sci. 43: 37-45.. 가 많은 것으로 나타났다. 하지만 Vrn-B1 유전자 변이는 장. 6. Chen F, Gao M, Zhang J, Zuo A, Shang X, Cui D. 2013.. 수형 계통의 농업형질의 차이가 없는 것으로 나타났다. 장수. Molecular characterization of vernalization response genes in bread wheat from the Yellow and Huai Valley of. 형은 Ppd-D1유전자 변이에 따른 간장이나 이삭 길이는 차이 가 없었지만, 출수 일수와 수수는 Ppd-D1b를 지닌 계통이 많 았지만, 1수립수, 천립중, 리터중과 수량은 Ppd-D1a를 지닌 계통이 높았다. 종실 특성을 나타내는 표지인자 변이는 출수 일수, 간장, 수장, 분얼이나 1수립수 및 수량에는 영향을 주지. China. BMC Plant Biol. 13: 199. 7. Duggan BL, Richards RA, van Herwaarden AF, Fettell NA. 2005. Agronomic evaluation of a tiller inhibition gene (tin) in wheat. I. Effect of yield, yield components. 않았지만 TaSus2-2B유전자의 Hap-L타입, TaGW2유전자의. and grain protein. Aust. J. Agric. Res. 56: 169-78. 8. Ellis MH, Spielmeyer W, Gale KR, Rebetzke GJ,. Hap-6A-G타입과 TaCwi-A1a 가 각각의 대립 유전자형보다. Richards RA. 2002. “Perfect” markers for the Rht-B1b. 천립중이 높게 나타났다. 향후 재배환경이 장수형의 농업형질. and Rht-D1b dwarfing genes in wheat. Theor. Appl. Genet. 105: 1038-1042.. 및 품질에 미치는 영향에 대한 평가가 필요하고, 만숙과 분얼 이 적은 장수형의 단점을 보완하기 위한 지속적인 연구가 필 요하다.. 사. 사. 9. Fischer RA. 2007. Understanding the physiological basis of yield potential in wheat. J. Agric. Sci. 145: 99-113. 10. Flintham E, Borner A, Worland A, Gale M. 1997. Optimizing wheat grain yield effects of Rht (gibberellin-insensitive) dwarfing genes. J. Agric. Sci. 128: 11-25. 11. Foulkes MJ, Slafer GA, Davies WJ, Berry PM, Sylvester-. 본 연구는 2015년도 농촌진흥청 국립식량과학원 박사후연. Bradley R, Martre P, Calderini DF, Griffiths S, Reynolds. 수과정지원사업(PJ008756012015)과 농촌진흥청 연구사업. MP. 2011. Raising yield potential of wheat. III. Optimizing.
(9) 장수형 밀의 계통 특성 분석 I. 농업 형질과 관련 유전자 특성. partitioning to grain while maintaining lodging resistance. J. Exp. Bot. 62: 469-486.. 227. 24. McIntosh RA, Hart GE, Devos KM, Gale MD, Rogers WJ. 1998. Catalogue of gene symbols for wheat. pp. 1–. 12. Fu D, Szűcs P, Yan L, Helguera M, Skinner JS, von. 235. In A.E. Slinkard (ed.) Proc. 9th Int. Wheat Genet.. Zitzewitz J, Hays PM, Dubcovsky J. 2005. Large deletions. Symp. Vol. 5. Univ. Extension Press, Univ. of Saskatchewan,. within the first intron in VRN-1 are associated with spring. Saskatoon, SK, Canada.. growth habit in barley and wheat. Mol. Gen. Genom. 273: 54-65. 13. Gaju O, Reynolds MP, Sparkes DL, Foulkes MJ. 2009.. 25. Miller ME, Chourey PS. 1992. The maize invertase-deficient miniature-1 seed mutation is associated with aberrant pedicel and endosperm development. Plant Cell 4: 297-305.. Relationships between large spike phenotype, grain number. 26. Miralles DJ, Slafer GA. 2007. Sink limitations to yield in. and yield potential in spring wheat. Crop Sci. 49: 961-73.. wheat : how could it be reduced? J. Agric. Sci. 145:. 14. Gaju O, Reynolds MP, Sparkes DL, Mayes S, Ribas-. 139-149.. Vargas G, Crossa J, Foulkes MJ. 2014. Relationships. 27. Perry MW, d’Antuono, MF. 1989. Yield improvement. between physiological traits, grain number and yield potential in a wheat DH population of large spike. and associated characteristics of some Australian spring wheat cultivars introduced between 1860 and 1982. Aust.. phenotype. Field Crops Res. 164: 126-35.. J. Agric. Res. 40: 457-472.. 15. Goncharov NP. 2003. Genetics of growth habit(spring vs. winter) in common wheat: Confirmation of the existence. 28. Pugsley AT. 1971. A genetic analysis of the spring-winter habit of growth in wheat. Aust. J. Agric. Res. 22: 21-23.. of dominant gene Vrn4. Theor. Appl. Genet. 107: 768-772.. 29. RDA. 2012. Standard of research and analysis for. 16. Jiang Q, Hou J, Hao C, Wang L, Ge H, Dong Y, Zhang X. 2011. The wheat (T. aestivum) sucrose synthase 2 gene. agricultural technology. pp. 339-365. 30. Reynolds MP, Foulkes MJ, Furbank R, Griffiths S, King. (TaSus2) active in endosperm development is associated. J, Murchie M, Parry M, Slafer G. 2012. Achieving yield. 17. Kamran A, Iqbal M, Spaner D. 2014. Flowering time in. gains in wheat. Plant Cell Enviorn. 35: 1799-1823. 31. Reynolds MP, Pellegrineschi A, Skovmand B. 2005. Sink. wheat (Triticum aestivum L.): a key factor for global. limitation to yield and biomass: a summary of investigations. adaptability. Euphytica 197: 1-26. 18. Kang CS, Seo SJ, Kim KH, Jung YG, Kim HS, Park JC,. in spring wheat. Ann. Appl. Biol. 146: 39-49. 32. Richards RA. 1988. A tiller inhibitor gene in wheat and. with yield traits. Funct. Integr. Genomics 11: 49-61.. Choi ID, Kim KH, Hyun JN, Kim KJ, Park KH, Lee CK,. its effect on plant growth. Aust. J Agric. Res. 39: 749-57.. Park GK, Park CS. 2013. A new plant type wheat “Iksan 370” with long spikes, high yield and hard wheat and. 33. Rosegrant MW, Cline SA. 2003. Global food security: challenges and policies. Science 302: 1917-1919.. breeding method. Korean Patent No. 10-2013-0120136.. 34. Santra DK, Santra M, Allan RE, Campbell KG, Kidwell. 19. Kato K, Yamagata H. 1998. Method for evaluation of chilling requirement and narrow-sense earliess of wheat cultivars. Jpn. J. Breed. 38: 172-186. 20. Ledent JF, Stoy V. 1988. Yield of winter wheat, a comparison of genotype from 1910 to 1976. Cereal Res. Comm. 16: 151-156. 21. Liu Y, He Z, Appels R, Xia X. 2012, Functional markers in wheat: current status and future prospects. Theor. Appl. Genet. 125: 1-10.. KK. 2009. Genetic and molecular characterization of vernalization genes Vrn-A1, Vrn-B1, and Vrn-D1 in spring wheat germplasm from the Pacific Northwest Region of the U.S.A. Plant Breed. 128: 576-584. 35. Shin DH, Choi MG, Kang CS, Park CS, Choi SB, Park YI. 2015. Preferential expression of cell elongation-related genes in leaves of the new elite wheat line Iksan370 with large spikes. Plant Biotechnol. Rep. 9: 97-105. 36. Snape JW, Butterworth K, Whitechurch E, Worland AJ.. 22. Ma D, Yan J, He Z, Wu L, Xia X. 2012. Characterization of a cell wall invertase gene TaCwi-A1 on common wheat. 2001. Waiting for fine times: genetics of flowering time in wheat. Euphytica 119: 185-190.. chromosome 2A and development of functional markers.. 37. Son JH, Cheong YK, Kim KH, Kim HS, Choi I, Park JC,. Mol. Breed. 29: 43-52. 23. MAFRA. 2014. Ministry of Agriculture, Food and Rural. Shin S, Lee CK, Park KG, Kang CS. 2014. Agricultural and physiological characteristics of new elite wheat line. Affairs statistical yearbook. pp.250-293.. “Iksan370” with Long Spike. J. Agric. Life Sci. 43: 92-96..
(10) 228. 韓育誌(Korean J. Breed. Sci.) 47(3), 2015. 38. Spielmeyer W, Richards R. 2004. Comparative mapping. Plant Cell 11: 177-189.. of wheat chromosome 1AS which contains the tiller inhibition. 43. Tosovic-Maric B, Kobiljski B, Obrehti D, Vapa L. 2008.. gene (tin) with rice chromosome 5S. Theor. Appl. Genet.. Evaluation of wheat Rht genes using molecular markers.. 109: 1303-10. 39. Stelmakh AF. 1998. Genetic systems regulating flowering response in wheat. Euphytica 100: 359-369. 40. Sturm A. 1999. Invertase. Primary structures, functions, and roles in plant development and sucrose partitioning. Plant Physiol. 121: 1-8. 41. Su ZQ, Hao CY, Wang LF, Dong YC, Zhang XY. 2011. Identification and development of a functional marker of. Genetika. 40: 31-38. 44. Worland AJ. 1996. The influence of flowering time genes on environmental adaptability in European wheats. Euphytica 89: 49-57. 45. Yan L, Helguera M, Kato K, Fukuyama S, Sherman J, Dubcovsky J. 2004. Allelic variation at the Vrn-1 promoter region in polyploidy wheat. Theor. Appl. Genet. 109: 1677-1686.. TaGW2 associated with grain weight in bread wheat. 46. Zhang J, Wang Y, Wu S, Yang J, Liu H, Zhou Y. 2012.. (Triticum aestivum L.). Theor. Appl. Genet. 122: 211-223. 42. Tang GQ, Lu¨scher M, Sturm A. 1999. Antisense. A single nucleotide polymorphism at the Vrn-D1 promoter region in common wheat is associated with vernalization. repression of vacuolar and cell wall invertase in transgenic carrot alters early plant development and sucrose partitioning.. response. Theor. Appl. Genet. 125: 1697-1704..
(11)
수치


+2
관련 문서
Micro- and nano-sized pores were formed on the surface of the alloy using PEO and anodization methods, and the pore shape change according to the Zr
Days to emergence and heading, duration of growth, leaf traits and stem diameter of sweet sorghum as affected by soils. Soil type Days
2001.. to determine influence of planting space and the number of plants per hill on agronomic characters, forage yield and quality of Jeju Italian millet..
Fourth, for the sub-factors of personality traits according to weight lifters’ education, while there was a high difference in neuroticism and agreeableness
⚫ User-level tools cannot monitor kernel level performance issues.. Performance
Kernel 1, which has an arithmetic intensity of 0.5 FLOPs/byte, is limited by memory bandwidth to no more than 8 GFLOPS/sec. Kernel 2, which has an arithmetic intensity of
- Calculus of variation (COV) and dynamic programming (DP) are companion methods since both are techniques to determine the optimal function y(x).. - A difference between COV and
• RBFN provides a global approximation to the target function, represented by a linear combination of many local kernel functions. The value
