Vitamin D Inhibits Expression and Activity of Matrix Metalloproteinase in Human Lung Fibroblasts (HFL-1) Cells
Seo Hwa Kim, M.D., Moon Seong Baek, M.D., Dong Sik Yoon, M.D., Jong Seol Park, M.D., Byoung Wook Yoon, M.D., Byoung Su Oh, M.D., Jinkyeong Park, M.D. and Hui Jung Kim, M.D.
Department of Internal Medicine, Wonkwang University Sanbon Hospital, Wonkwang University College of Medicine, Gunpo, Korea
Background: Low levels of serum vitamin D is associated with several lung diseases. The production and activation of matrix metalloproteinases (MMPs) may play an important role in the pathogenesis of emphysema. The aim of the current study therefore is to investigate if vitamin D modulates the expression and activation of MMP-2 and MMP-9 in human lung fibroblasts (HFL-1) cells.
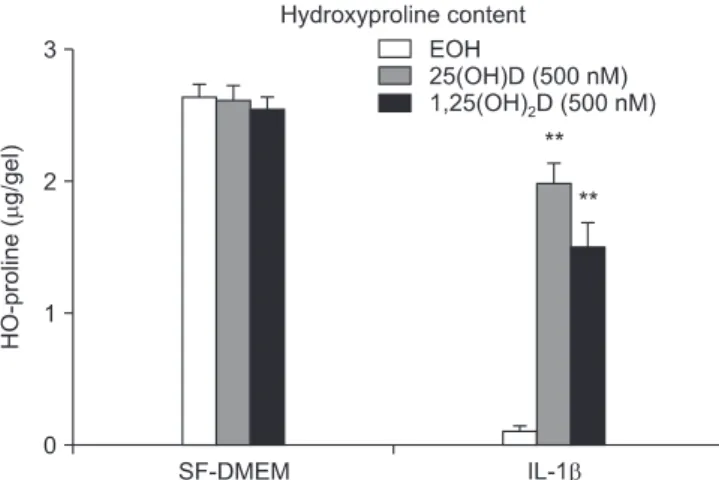
Methods: HFL-1 cells were cast into three-dimensional collagen gels and stimulated with or without interleukin-1β (IL- 1β) in the presence or absence of 100 nM 25-hydroxyvitamin D (25(OH)D) or 1,25-dihydroxyvitamin D (1,25(OH)
2D) for 48 hours. Trypsin was then added into the culture medium in order to activate MMPs. To investigate the activity of MMP- 2 and MMP-9, gelatin zymography was performed. The expression of the tissue inhibitor of metalloproteinase (TIMP- 1, TIMP-2) was measured by enzyme-linked immunosorbent assay. Expression of MMP-9 mRNA and TIMP-1, TIMP-2 mRNA was quantified by real time reverse transcription polymerase chain reaction.
Results: IL-1β significantly stimulated MMP-9 production and mRNA expression. Trypsin converted latent MMP- 2 and MMP-9 into their active forms of MMP-2 (66 kDa) and MMP-9 (82 kDa) within 24 hours. This conversion was significantly inhibited by 25(OH)D (100 nM) and 1,25(OH)
2D (100 nM). The expression of MMP-9 mRNA was also significantly inhibited by 25(OH)D and 1,25(OH)
2D.
Conclusion: Vitamin D, 25(OH)D, and 1,25(OH)
2D play a role in regulating human lung fibroblast functions in wound repair and tissue remodeling through not only inhibiting IL-1β stimulated MMP-9 production and conversion to its active form but also inhibiting IL-1β inhibition on TIMP-1 and TIMP-2 production.
Keywords: Vitamin D; Matrix Metalloproteinase 9; Fibroblasts
Copyright © 2014 The Korean Academy of Tuberculosis and Respiratory Diseases. All rights reserved.
Address for correspondence: Hui Jung Kim, M.D.
Division of Pulmonary and Critical Care Medicine, Department of Internal Medicine, Wonkwang University Sanbon Hospital, 327 Sanbon-ro, Gunpo 435- 040, Korea
Phone: 82-31-390-2300, Fax: 82-31-390-2999, E-mail: [email protected] Received: Mar. 5, 2014
Revised: Mar. 21, 2014 Accepted: Jun. 23, 2014
cc It is identical to the Creative Commons Attribution Non-Commercial License (http://creativecommons.org/licenses/by-nc/3.0/).