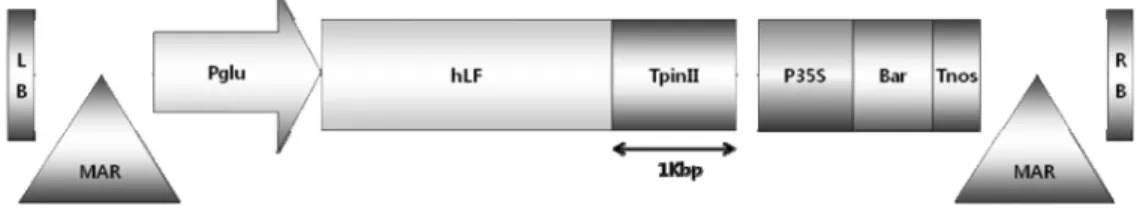
Development of transgenic rice lines expressing the human lactoferrin gene
6
0
0
전체 글
(2)
(3)
(4)
(5)
(6)
수치


관련 문서
