류마티스 관절염의 활막 섬유모세포에서 Hypoxia/
reoxygenation에 의한 종양 억제 인자 PTEN 발현의 조절
전북대학교 의과대학 내과학교실, 임상연구소 이 학 용․류 완 희
= Abstract =
Hypoxia/reoxygenation Regulates Expression of Tumor Suppressor PTEN in Rheumatoid Synovial Fibroblasts
Hak Yong Lee, M.D., Wan Hee Yoo, M.D.
Department of Internal Medicine, and Research Institute of Clinical Medicine, Medical School, Chonbuk National University, Jeonju, Korea
Objective: The lack of phosphatase and tensin homolgue deleted on chromosome ten (PTEN) expression was described in rheumatoid synovial tissues and synovial fibroblasts, but exact cause of that in RA is not well known. Hypoxic conditions are thought to exist in inflamed arthritic synovium and therefore, this study was designed to investigate the effects of hypoxia/
reoxygenation on the expression of PTEN in synovial fibroblasts of rheumatoid arthritis.
Methods: Synovial fibroblasts were isolated from synovial tissues of patients suffering from RA and hypoxic culture was performed by incubating cells in 5% CO2 incubator held at 3%
oxygen by the addition of nitrogen gas for 24 hours. Then synovial fibroblasts were cultured for 10 min under normoxic condition for reoxygenation. To know the expression of PTEN and phosphorylated Akt (p-Akt) in synovial fibroblasts, Western blotting analysis was performed. The expression of PIP3 kinase and PTEN was analyzed by immunocytochemical staining.
Results: There were less PTEN expression in rheumatoid synovial fibroblasts than that of healthy controls. Hypoxic/reoxygenation stimuli induced down-regulation of PTEN expression in the rheumatoid synovial fibroblasts. In contrast, the expression of PIP3 and p-Akt was increased after stimulation with hypoxia/reoxygenation.
Vol. 11, No. 4, December, 2004
ꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏ
<접수일:2004년 4월 13일, 심사통과일:2004년 9월 15일>
※통신저자:류 완 희
전북 전주시 덕진구 금암동 634-18번지 전북대학교 의과대학 내과학교실, 임상연구소
Tel:063) 254-1609, Fax:063) 250-1672, E-mail:[email protected] 이 논문은 전북대학교병원 임상연구소의 학술연구비 지원에 의하여 연구되었음.
서 론
류마티스 관절염은 전신성 자가면역질환으로 대칭 성 관절염과 이로 인한 관절의 비가역적 파괴를 초 래하며 조직학적으로는 활막의 비후와 염증세포의 광범위한 침착을 특징으로 한다1,2). 류마티스 관절염 에서 염증성 활막은 지속적으로 증식하여 판누스 (pannus)를 형성하며, 이는 류마티스 관절염에서 관 절의 파괴에 중요한 역할을 하는 것으로 알려져 있 다. 판누스의 활막 섬유모세포는 관절연골에 유착하 고 파괴성 성장(invasive growth)을 하는 종양 조직과 유사한 특성을 갖고 있으며 관절의 파괴에 관여하는 것으로 알려져 있다3).
PTEN은 종양 억제 유전자로서 tyrosine phospha- tase activity를 가지며 세포골격 단백질(cytoskeletal proteins)인 tensin과 auxilin에 상동성(homology)을 갖 는 물질이다4,5). 여러 가지 악성 종양에는 PTEN의 변이(mutation)가 보고되어 있으며 이는 암의 침습성 (invasiveness)과 전이와 관련되는 것으로 알려져 있 다4-6). 류마티스 관절염에서는 염증성 활막에 특징적 으로 PTEN이 감소되어 있으며, 이는 관절의 파괴에 중요한 역할을 하는 활막조직의 활막 섬유모세포의 침습성과 파괴성의 원인으로 알려져 있다7). 그러나 이러한 PTEN의 발현에 미치는 조절인자에 대해서는 잘 알려져 있지 않으며, 류마티스 관절염에서도 그 원인이 잘 알려져 있지 않다.
류마티스 관절염의 염증성 활막은 현저한 저산소 증을 동반하며8,9), 관절 운동 등에 의한 관절 내의 산소분압의 변화는 저산소증 재관류의 순환을 초래 한다10). Hypoxia/reoxygenation에 의해 생성된 산화반 응의 대사물질과 생화학적 변화는 대부분의 염증성 질환의 병인에 중요한 역할을 하는 것으로 알려져 있
으며, 류마티스 관절염에서도 관절의 구성 성분인 결 체조직 등에 직접적으로 산화적 손상(oxidative dam- 을 주고, 면역성 염증반응에 관련된 다양한 단백질의 유전자 발현에 영향을 주어 염증반응에 관련되는 것 으로 알려져 있다11,12). 특히, 염증 전구물질, 주요 주조 직적합체의 항원과 유착분자의 발현을 증가시키며13), vascular endothelial growth factor (VEGF)의 증가를 통 해서 맥관형성(angiogenesis)에 관여하고14,15), matrix metalloproteinase의 발현을 증가시켜16) 관절 연골과 주위 조직의 파괴에 관련하는 것으로 알려져 있다.
따라서 류마티스 관절염에서 활막의 hypoxia/reoxy- genation은 활막 섬유모세포의 PTEN 발현에 영향을 줄 수 있는 것으로 추정되지만 이에 대한 연구는 없 는 실정이다.
저자는 정상인 및 류마티스 관절염 환자에서 활막 섬유모세포를 배양하여 PTEN의 발현에 차이가 있는 지를 알아보고, 류마티스 관절염 환자의 활막 섬유 모세포에서 hypoxia/reoxygenation이 PTEN 발현에 미 치는 영향을 조사하고자 하였다. 또한 hypoxia/reoxy- genation이 PTEN에 미치는 영향을 규명하기 위하여 류마티스 관절염 환자의 활막 섬유모세포에서 PTEN 억제가 PIP3와 p-Akt 발현에 미치는 영향을 알아보 고자 하였다.
대상 및 방법 1. 재료
활막 섬유모세포에서 단백질 발현 상태를 관찰하 기 위하여 1차 항체로는 anti-PTEN monoclonal anti- body와 anti-p-Akt monoclonal antibody를 Santa Cruz Biotechnology (Santa Cruz, CA, USA)로부터 구입하 였으며, anti-PIP3 monoclonal antibody는 Echelon (Salt Lake City, UT, USA)에서 구입하였다. 2차 항체로는 Conclusion: These studies suggest that hypoxia/reoxygenation could cause the reduced expression of PTEN in rheumatoid synovial fibroblasts and thus it might thereby contribute to the invasive behaviour of rheumatoid synovial fibroblasts by maintaining their aggressive phenotype in RA.
ꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏ Key Words: Hypoxia/reoxygenation, Phosphatase and tensin homolgue deleted on chro-
mosome ten, Synovial fibroblasts
goat anti-mouse IgM-TRITC를 Santa Cruz Biotechno- logy (Santa Cruz, CA, USA)에서 구입하였다. 또한 세포배양을 위하여 DMEM와 fetal bovine serum은 Gibco-BRL (Gaithersburg, MD, USA)에서 구입하였으 며, HBSS (Hanks' balanced salt solution)와 EDTA은 Sigma Chemical Co. (St. Louis, MO, USA)로부터 구 입하였다.
2. 활막 섬유모세포의 분리와 배양
미국류마티스학회(American College of Rheumato- logy)의 진단기준에17) 적합한 류마티스 관절염 환자 와 염증성 관절 질환이 아닌 다른 질환으로 관절 수 술이 필요한 환자에서 무릎 관절의 활막 제거술이나 관절 치환술이나 기타 수술 중에 얻은 조직으로부터 Matsuoka 등18)이 기술한 방법을 적용하여 활막 섬유 모세포를 분리하였다. 활막조직을 가위를 이용하여 잘게 자른 후에 collagenase (Sigma Chemical Co. St.
Louis, Missouri, USA)로 37oC에서 45분간 처리하였 다. 세포부유액을 나일론 망에 통과시키고 남은 세 포를 DMEM-10% FBS가 담긴 culture flask에 배양하 였다. 비부착세포를 PBS로 3회 세척하여 제거하고 남아 있는 부착세포를 계대 배양하였다. 배양세포가 활막 섬유세포임을 규명하기 위하여 anti-CD3 단일 클론 항체, anti-CD68 단일클론 항체, anti-CD20 단일 클론 항체, anti-vWF 항체 등을 사용한 flow cytmetry (Becton-Dickinson, CA, USA)를 이용하여 배양세포 내에 T 림프구, 대식세포, B 림프구, 내피세포 등이 1% 미만임을 입증하였다. 또한 활막 섬유모세포는 uridine diphosphoglucose dehydrogenase 염색을 이용하 여 확인하였으며, 계대배양 3∼7회의 세포를 사용하 였다.
3. 활막 섬유모세포의 hypoxia/reoxygenation 처리
저산소 조건을 만들기 위하여 5% CO2 incubator에 nitrogen gas를 주입하여 incubator의 조건을 5% CO2,
>95% N2, <3% O2가 되도록 하였다. 37oC에서 배 양하였으며, 24시간 동안 저산소증에 노출시키는 동 안에는 incubator 내의 O2 농도를 O2 분석기를 이용 하여 지속적으로 감시하였다. 활막 섬유모세포의 re- oxygenation을 위해서는 정상 산소 상태에서 10분간 배양하였다. 저산소증 상태에서 활막 섬유모세포의
생존율을 조사하기 위하여 trypan blue exclusion 방 법19)을 이용하였다.
4. PTEN과 p-Akt의 Western blot analysis
활막 섬유모세포에서 PTEN과 p-Akt의 발현을 조 사하기 위하여 Western blot analysis를 시행하였다.
활막 섬유모세포를 3회 PBS로 세척한 후 100 mm plate에 1 mL의 PBS-TDS (1×PBS, 1% Triton X-100, 0.05% sodium deoxycholate, 0.01% SDS, 0.5µg/mL leupeptin, 1 mM EDTA, 1µg/mL pepst atin, 0.2 mM PMSF) 용액을 첨가, 15분간 얼음 위에서 반응시킨 후 13,000 rpm에서 5분간 원심하여 상층액을 얻었 다. 단백질 농도는 BSA (bovine serum albumin)을 표 준단백으로 Bio-Rad Protein Assay Kit (BIO-RAD Co.)를 사용하여 정량하였다. 10µg의 세포 용해액을 10% gel (BIO-RAD Co.) SDS-PAGE (Polyacrylamide Gel Electrophoresis)로 변성 분리하였고, 이를 nitro- cellulose membrane (Hybond-C Amer sham Co.)에 80 Voltage로 2시간 전기영동을 하였다. Membrane를 5%
탈지유가 함유된 TBS (PBS+0.1% Tween 20) 용액 으로 상온에서 1시간 정도 반응시킨 후 비특이적 결 합을 차단하였다. PTEN과 p-Akt의 발현 양을 측정 하기 위한 1차 항체로는 anti-PTEN monoclonal anti- body와 anti-p-Akt monoclonal antibody (Santa Cruz, CA, USA)를 1:1,000의 TBS 용액에 희석하여 상온에서 1시간 반응시킨 후 TBS로 3회 세척하였다. 2차 항체 로는 horseradish peroxidase가 결합된 anti- mouse IgG (Santa Cruz, CA, USA)를 1:5,000으로 희석하여 상 온에서 1시간 동안 반응시켰다. TBS로 3회 세척 후 ECL 기질(Amer sham Co.)과 1분 30초간 반응시킨 후 image reader LAS-1000 lite로 측정하였으며 image gauge로 도표화하였다(Fujifilm, Japan).
5. PTEN과 PIP3의 Immunocytochemistry
PTEN과 PIP3의 발현을 조사하기 위하여 면역화학 염색(immunocytochemistry)을 시행하였다. 세포 1×103 개를 6-well plate에 저산소 상태에서 24시간 동안 배 양하였다. PBS로 3회 세척한 후에 4% paraformal- dehyde로 10분간 고정하였다. 고정 후 슬라이드를 PBS로 3회 세척한 후 0.01% H2O2와 0.05% DAB (dia- minobenzidine tetrahydrochloride)의 용액에 5분간 반
응시킨 후 30분간 항체의 비특이적 결합을 제거하기 위하여 blocking 용액(3% bovine serum albumin, 0.5%
Triton X-100 in PBS)으로 반응시켰다. 1차 항체로는 anti-PTEN monoclonal antibody와 anti-PIP3 monoclonal antibody (Santa Cruz, CA, USA)를 500배 blocking 용 액에 희석하여 20시간 정도 실온에서 반응시킨 후 PBS로 3회 세척하였으며, biotin이 결합된 anti-rabbit 항체를 blocking용액에 실온에서 10분간 반응시킨 후 PBS로 3회 세척하였다. Avidin 용액과 biotin과 per- oxidase가 결합된 항체(Vectast ain)의 용액에 슬라이 드를 5분간 반응시키고 PBS로 3회 세척하여 반응을 정지시켰다. 슬라이드에 커버슬라이드를 봉합한 후 Axiovert S100 fluorescence microscope (carlZeiss, Jena, Germany)를 이용하여 관찰하였다.
결 과 1. 활막 섬유모세포에서 PTEN 발현
류마티스 관절염 환자와 정상인의 활막 섬유모세 포에서 종양 억제 유전자인 PTEN의 발현 양상을 알 아보기 위하여 Western blotting을 시행하였다. 정상 인 대조군의 활막 섬유모세포에 비해(C1∼C3 in lanes 1∼3) 류마티스 관절염 환자군의 활막 섬유모세포에 서(RA1∼RA3 in lanes 4∼6) PTEN의 발현이 현저하 게 감소되어 있음을 관찰할 수 있었다(그림 1).
2. Hypoxia/reoxygenation이 활막 섬유모세포에서 PTEN 발현에 미치는 영향
Hypoxia/reoxygenation이 류마티스 관절염 환자의 활막 섬유모세포에서 감소된 PTEN 발현의 원인인지 를 알아보기 위하여 류마티스 관절염 환자로부터 분
Fig. 1. PTEN expression in the synovial fibroblasts.
The PTEN expression was examined by West- ern blotting in synovial fibroblasts of healthy controls (C1∼C3, n=3) and patients with rheumatoid arthritis (RA1∼RA3, n=3). PTEN expression was less in the synovial fibroblasts of patients with rheumatoid arthritis than that of healthy controls. The analysis of PTEN was examined triplicates and the density of PTEN electrophoretic band was performed with the image reader LAS-1000 lite (Fujifilm, Japan).
The results shown above are representative of three separate experiments.
PTEN/Actin ratio
0 0.5 1 Actin PTEN
C1 C2 C3 RA1 RA2 RA3
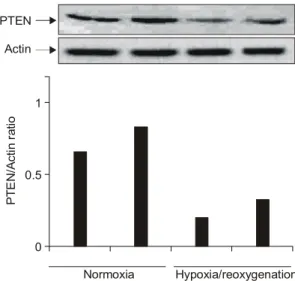
Fig. 2. PTEN expression after stimulation with hypoxia/
reoxygenation. The expression of PTEN in the synovial fibroblasts of patients with RA was examined by Western blotting. The expression of PTEN was reduced after hypoxia/reoxy- genation stimuli in rheumatoid synovial fibro- blasts. The density of PTEN electrophoretic band was performed with the image reader LAS-1000 lite (Fujifilm, Japan). The results shown above are representative of three sepa- rate experiments.
PTEN/Actin ratio
0 0.5 1 Actin PTEN
Normoxia Hypoxia/reoxygenation
리하여 배양한 활막 섬유모세포을 저산소 조건에서 24시간 노출하였으며 reoxygenation을 위하여 정상산 소 상태에서 10분간 배양하였다. 24시간 동안 저산
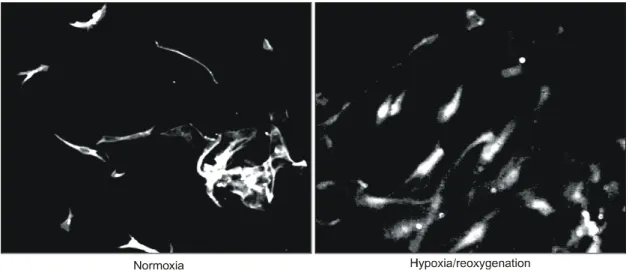
소증에서의 배양과 10분간 정상산소 상태에서의 배 양이 활막 섬유모세포의 생존율에 미치는 영향을 알 아보기 위하여 trypan blue exclusion 검사를 시행하 Fig. 3. PTEN expression after stimulation with hypoxia/reoxygenation. The expression of PTEN in the synovial
fibroblasts of patients with RA was examined by immunocytochemical stain. The expression of PTEN was decreased after hypoxia/reoxygenation stimuli in rheumatoid synovial fibroblasts. The analysis of PTEN was examined triplicates with Axiovert S100 fluorescence microscope (×100). The results shown above are representative of three separate experiments.
Normoxia
Fig. 4. PIP3 expression after stimulation with hypoxia/ reoxygenation. The expression of PIP3 in the rheumatoid synovial fibroblasts was examined by immunocytochemical stain. The expression of PIP3 was increased after hypoxia/reoxygenation stimuli in rheumatoid synovial fibroblasts. The analysis of PIP3 was examined triplicates with Axiovert S100 fluorescence microscope (×100). The results shown above are representative of three separate experiments.
Normoxia
였으며, 배양 후 활막 섬유모세포의 생존율은 98%
이상을 유지하였다(data not shown). 정상산소 상태에 서 배양한 활막 섬유모세포와 hypoxia/reoxygenation 자극을 준 후의 활막 섬유모세포에서 PTEN 발현의 변화를 비교하였을 때, hypoxia/reoxygenation 자극을 받은 류마티스 관절염 환자의 활막 섬유모세포에서 PTEN 발현이 현저하게 감소된 것을 관찰할 수 있었 다(그림 2). 면역 세포화학 검사에서도 정상산소 상 태에서 배양한 활막 섬유모세포에 비해 hypoxia/reoxy- genation 자극을 준 후의 활막 섬유모세포에서 PTEN 발현이 현저하게 감소된 것이 관찰되었다(그림 3).
3. Hypoxia/reoxygenation이 활막 섬유모세포에서 PIP3의 발현에 미치는 영향
류마티스 관절염 환자의 활막 섬유모세포에서 hy- poxia/reoxygenation 자극에 의해 PTEN 발현이 감소 함을 관찰할 수 있었으며, 이러한 결과를 뒷받침하 기 위하여 PTEN에 의해서 발현을 조절 받는 PIP3의 발현에 hypoxia/reoxygenation 자극이 어떠한 영향을 주는지 조사하였다. 세포막에서 발현되는 PIP3는 면 역 세포화학 검사를 이용하여 관찰하였으며, 정상산 소 상태에서 배양한 류마티스 관절염 환자의 활막 섬유모세포에 비해 hypoxia/reoxygenation 자극을 준 후의 활막 섬유모세포에서 PIP3의 발현이 현저하게 증가된 것을 관찰할 수 있었다(그림 4).
4. Hypoxia/reoxygenation이 활막 섬유모세포에서 p-AKT의 발현에 미치는 영향
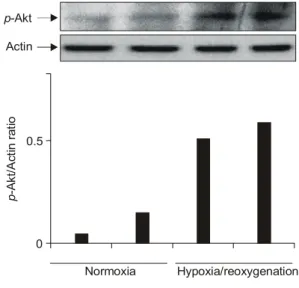
류마티스 관절염 환자의 활막 섬유모세포에서 hy- poxia/reoxygenation 자극에 의해 PTEN 발현이 조절 되는 것을 뒷받침하기 위하여 PTEN의 하위단계의 신호에 해당하는 p-Akt를 정상산소 상태에서 배양한 활막 섬유모세포와 hypoxia/reoxygenation 자극을 준 후의 활막 섬유모세포에서 Western blotting을 이용하 여 관찰하였다. 류마티스 관절염 환자의 활막 섬유 모세포에서 정상산소 상태에서 배양한 활막 섬유모 세포에 비해 hypoxia/reoxygenation 자극을 준 후의 활막 섬유모세포에서 p-Akt의 발현이 현저하게 증가 하는 것을 관찰할 수 있었다(그림 5).
고 찰
류마티스 관절염은 만성 염증성 질환으로 염증성 활막염과 활막의 비후 및 점진적인 침범 관절의 대 칭성 파괴를 특징으로 한다. 염증성 활막 조직에는 T 림프구, 내피세포, 활막 섬유모세포 등의 다양한 세포군이 존재하여 류마티스 관절염의 병인에 관련 되며, 특히 활막 섬유모세포는 염증과 무관하게 관 절의 파괴에 중요한 역할을 하는 것으로 알려져 있 다. 그렇지만 활막 섬유모세포의 활성화 기전과 어 떻게 관절 연골을 파괴시킬 수 있는지에 대해서는 잘 알려져 있지 않다. 특히 활성화된 활막 섬유모세 포는 종양 세포와 유사한 특성을 갖고 있어 관절연 골에 유착하고 침습성 성장을 하는 특징을 갖는 것 으로 알려져 있지만3), 그 원인은 알려져 있지 않다.
Fig. 5. p-Akt expression after stimulation with hypoxia/
reoxygenation. The expression of p-Akt in the synovial fibroblasts of patients with RA was examined by Western blotting. The expression of p-Akt was increased after hypoxia/reoxy- genation stimuli in rheumatoid synovial fibro- blasts. Actin was used as a house-keeping pro- tein. The density of p-Akt electrophoretic band was performed with the image reader LAS- 1000 lite (Fujifilm, Japan). The results shown above are representative of three separate ex- periments.
p-Akt/Actin ratio
0 0.5 Actin p-Akt
Normoxia Hypoxia/reoxygenation
이러한 류마티스 관절염에서 활막 섬유모세포의 특징적인 소견과 관련하여 종양 억제 유전자인 p53 의 역할에 관한 연구가 보고되었으며, 류마티스 관 절염의 활막과 활성화된 활막 섬유모세포에 p53의 변이가 있음을 보고하였다20). 그렇지만 류마티스 관 절염에서 p53의 변이에 대한 결과가 일관되지 않고 매우 다양한 결과를 보이는 점은 p53의 변이가 류마 티스 관절염의 특징적인 염증성 변화에 이차적으로 발생하는 것이지 그 자체가 병인의 근본적인 원인은 아니며, 활막 섬유모세포의 종양 세포와 유사한 특 성의 원인으로는 보고 있지 않다.
PTEN은 종양 억제 유전자로서 tyrosine phosphatase activity를 갖고 있으며 세포골격 단백질인 tensin과 auxilin에 상동성을 갖는 물질로서4,5), 여러 가지 악성 종양에서 PTEN의 변이가 발견되었으며4-6), 류마티스 관절염에서는 염증성 활막에 특징적으로 PTEN이 감 소되어 있는 것으로 알려졌다7). 일부 보고에서는 류 마티스 관절염의 활막 섬유모세포에 tyrosine kinase 의 활성도가 증가해 있음을 보여 주었는데, 이는 류 마티스 관절염에서 PTEN의 감소는 tyrosine kinases 와 phosphatases의 불균형을 초래하여 관절의 파괴에 중요한 역할을 하는 활막조직의 활막 섬유모세포의 침습성과 파괴성의 특징을 갖게 하는 것으로 보고하 였다21). 본 연구에서도 정상인의 활막 섬유모세포에 비해 류마티스 관절염 환자의 활막 섬유모세포에서 PTEN의 발현이 현저하게 감소된 것을 관찰할 수 있 었으며, 이는 이전의 보고와 같이 류마티스 관절염 의 병인에 PTEN이 관련되어 있음을 뒷받침 해주는 증거가 될 수 있음을 보여준다. 그러나 이러한 PTEN의 감소에 대한 기전에 대해서는 정확히 알려 져 있지 않다. 류마티스 관절염의 활막에 증가해 있 는 TGF-β가 PTEN 감소의 원인으로 생각하고 있지 만 활막에 전반적으로 TGF-β의 발현이 증가되어 있 는 것은 활막의 일부에만 PTEN의 발현이 감소하는 것을 적절히 설명해 주지 못하고 있다22).
류마티스 관절염에서 활막의 저산소증은 ischemia- reperfusion에 의한 reactive oxygen radical의 생성을 통해서 endogenous biomolecules인 콜라겐, proteogly- cans, ceruloplasmin, immunoglobulin23), protease inhibi- tor24), 결체조직, 단백질과 glycosaminoglycans25) 등에 산화적 손상을 일으키는 것으로 알려져 있다. 또한,
NF-κB 등과 같은 전사인자의 oxidative induction을 일으켜 cytokine, MHC 항원과 VEGF의 발현을 증가
시키고14,15), 염증반응에서 조직의 파괴에 중요한 ma-
trix metalloproteinase의 발현과 관련된 Activation Protein-1 (AP-1)의 oxidative induction에 관련되는 것 으로 알려져 있다16). 또한 최근에 저자들은 hypoxia /reoxygenation에 의한 자극이 류마티스 관절염 환자 의 활막 섬유모세포에서 유착분자인 ICAM-1의 발현 을 증가시키고 염증세포와의 유착성 반응이 증가하 여 류마티스 관절염에 반응할 수 있음을 보고한 바 있다13). 이와 같이 류마티스 관절염에서의 hypoxia/
reoxygenation은 여러 가지 면역반응에 영향을 주어 류마티스 관절염의 병인에 중요한 역할을 할 수 있 으며 활막 섬유모세포의 PTEN 발현에 영향을 줄 수 있을 것으로 추정되지만 이에 대한 연구는 없는 실 정이다.
본 연구에서는 류마티스 관절염 환자의 활막 섬유 모세포에 24시간 동안 hypoxia/reoxygenation의 자극 을 주었을 때 PTEN의 발현이 감소하는 것을 관찰할 수 있었으며, 이는 류마티스 관절염에서 지속적인 hypoxia/reoxygenation에 의한 자극이 PTEN의 감소의 원인이 될 수 있으며, 또한 관절 파괴에 중요한 역 할을 하는 활막 섬유모세포의 관절의 연골에 유착하 고 침습성 성장을 하는 특징을 갖게 하는 원인 인자 로 생각할 수 있게 해주는 근거가 될 수 있는 결과 이다. 더 나아가 본 연구에서는 류마티스 관절염의 활막 섬유모세포에서 PTEN에 의해 조절을 받는 것 으로 알려진 PIP3와 p-Akt의 발현을 hypoxia/reoxy- genation의 자극 전후에 조사하여 hypoxia/reoxygena- tion의 자극 후에 PTEN의 감소와 함께 PIP3와 p-Akt 의 발현이 현저하게 증가하는 것을 관찰할 수 있었 다. 류마티스 관절염의 활막 섬유모세포에서 hypoxia/
reoxygenation의 자극 후에 나타나는 PIP3와 p-Akt의 증가는 hypoxia/reoxygenation에 의한 자극이 PTEN의 감소의 중요한 원인이 될 수 있음을 보여주는 간접 적인 증거라 할 수 있다.
결론적으로, 류마티스 관절염에서 활막 섬유모세 포는 정상인의 비해 종양 억제인자인 PTEN의 발현 이 현저하게 감소해 있으며, hypoxia/reoxygenation에 의한 자극은 활막 섬유모세포의 PTEN 발현의 감소 를 일으키는 중요한 원인이 되며, hypoxia/reoxy-
genation의 자극에 의한 PIP3와 p-Akt의 발현의 증가 는 이를 간접적으로 증명해 주고 있다. 따라서 류마 티스 관절염의 관절에 존재하는 hypoxia/reoxygena- tion에 의한 자극은 관절 파괴에 중요한 역할을 하는 활막 섬유모세포가 연골에 유착하고 침습성 성장을 하는 특징을 갖게 하는 중요한 원인이라고 생각한 다. 따라서 저산소증 상태의 과정을 조절할 수 있는 방법은 류마티스 관절염의 치료의 한 방법으로써 도 움을 줄 수 있을 것으로 추정되며, PTEN이 류마티 스 관절염의 병인에 관여하는 구체적인 기전에 대해 서는 보다 많은 연구가 필요할 것으로 보인다.
결 론
류마티스 관절염에서 활막 섬유모세포는 관절의 파괴에 중요한 역할을 하며, 특징적인 관절 연골에 유착하고 침습성 성장을 특징으로 하지만 이에 대한 구체적인 기전은 알려져 있지 않다. 종양억제 유전 자인 PTEN은 여러 가지 암에서 변이를 보여 암의 전이와 주변 장기의 침습에 관련되며, 류마티스 관 절염에서도 병인에 중요한 역할을 하는 것으로 알려 져 있다. 저자의 연구에서는 정상인에 비해 류마티 스 관절염 환자의 활막 섬유모세포에서 현저하게 PTEN의 발현이 감소했고, hypoxia/reoxygenation에 의 한 자극은 활막 섬유모세포에서 PTEN 발현을 감소 시키며, down-stream signals인 PIP3와 p-Akt의 발현을 현저히 증가시켰다. 이는 류마티스 관절염에서 hy- poxia/reoxygenation은 관절 파괴에 중요한 역할을 하 는 활막 섬유모세포가 연골에 유착하고 침습성 성장 을 하는 특징을 갖게 하는 중요한 원인이기 때문이 라고 추측되며, PTEN이 류마티스 관절염의 병인에 어떻게 관여하는지에 대한 추가적인 연구가 필요할 것으로 보인다.
REFERENCES
1) Keyszer GM, Heer AH, Gay S. Cytokines and on- cogenes in cellular interactions of rheumatoid arthritis.
Stem Cells 1994;12:75-86.
2) Gay S, Gay RE, Koopman WJ. Molecular and cel- lular mechanisms of joint destruction in rheumatoid arthritis: two cellular mechanisms explain joint des- truction? Ann Rheum Dis 1993;52:S39-47.
3) Muller-Ladner U, Gay RE, Gay S. Cellular pathways of joint destruction. Curr Opin Rheumatol 1997;9:
213-20.
4) Li J, Yen C, Liaw D, Podsypanina K, Bose S, Wang SI, et al. PTEN, a putative protein tyrosine phos- phatase gene mutated in human brain, breast, and prostate cancer. Science 1997;275:1943-7.
5) Steck PA, Pershouse MA, Jasser SA, Yung WK, Lin H, Ligon AH, et al. Identification of a candidate tumour suppressor gene, MMAC1, at chromosome 10q23.3 that is mutated in multiple advanced cancers.
Nature Genet 1997;15:356-62.
6) Teng DH, Hu R, Lin H, Davis T, Iliev D, Frye C, et al. MMAC1/PTEN mutations in primary tumor specimens and tumor cell lines. Cancer Res 1997;
57:5221-5.
7) Pap T, Franz JK, Hummel KM, Jeisy E, Gay R, Gay S. Activation of synovial fibroblasts in rheumatoid arthritis: lack of Expression of the tumour suppressor PTEN at sites of invasive growth and destruction.
Arthritis Res 2000;2:59-64.
8) Paleolog EM, Young S, Stark AC, McCloskey RV, Feldmann M, Maini RN. Modulation of angiogenic vascular endothelial growth factor by tumor necrosis factor alpha and interleukin-1 in rheumatoid arthritis.
Arthritis Rheum 1998;41:1258-65.
9) Allen RE, Blake DR, Nazhat NB, Jones P. Super- oxide radical generation by inflamed human synovium after hypoxia. Lancet 1989;29:282-3.
10) Unsworth J, Outhwaite J, Blake DR, Morris CJ, Free- man J. Dynamic studies of the relationship between intraarticular, synovial fluid oxygen tension, and lipid peroxidation in the inflamed knee: an example of reperfusion injury. Ann Clin Biochem 1988;25:
8S-11S.
11) Ghezzi P, Dinarello CA, Bianchi M, Rosandich ME, Repine JE, White CW. Hypoxia increases production of interleukin-1 and tumor necrosis factor by human mononuclear cells. Cytokine 1991;3:189-94.
12) Hempel SL, Monick MM, Hunninghake GW. Effect of hypoxia on release of IL-1 and TNF by human alveolar macrophages. Am J Respir Cell Mol Biol 1996;14:170-6.
13) Han MK, Kim JS, Park BH, Kim JR, Hwang BY, Lee HY, et al. NF-kappaB-dependent lymphocyte hyper- adhesiveness to synovial fibroblasts by hypoxia and reoxygenation: potential role in rheumatoid arthritis.
J Leukoc Biol 2003;73:525-9.
14) Hitchon C, Wong K, Ma G, Reed J, Lyttle D, El-
Gabalawy H. Hypoxia-induced production of stromal cell-derived factor 1 (CXCL12) and vascular endo- thelial growth factor by synovial fibroblasts. Arthritis Rheum 2002;46:2587-97.
15) Berse B, Hunt JA, Diegel RJ, Morganelli P, Yeo K, Brown F, et al. Hypoxia augments cytokine (trans- forming growth factor-beta (TGF-beta) and IL-1)- induced vascular endothelial growth factor secretion by human synovial fibroblasts. Clin Exp Immunol 1999;115:176-82.
16) Cha HS, Ahn KS, Jeon CH, Kim J, Song YW, Koh EM. Influence of hypoxia on the expression of matrix metalloproteinase-1, -3 and tissue inhibitor of metal- loproteinase-1 in rheumatoid synovial fibroblasts. Clin Exp Rheumatol 2003;21:593-8.
17) Arnett FC, Edworthy SM, Bloch DA, McShane DJ, Fries JF, Cooper NS, et al. The American Rheu- matism Association 1987 revised criteria for the classification of rheumatoid arthritis. Arthritis Rheum 1988;31:315-24.
18) Matsuoka N, Eguchi K, Kawakami A, Ida H, Na- kashima M, Sakai M, et al. Phenotypic characteristics of T cells interacted with synovial cells. J Rheumatol 1991;18:1137-42.
19) Rosengard BR, Cochrane DE. Complementmediated cytolysis: a quick, simple method for determining levels of immunoglobulin E bound to mast cells. J
Histochem Cytochem 1983;131:441-4.
20) Firestein GS, Echeverri F, Yeo M, Zvaifler NJ, Green DR. Somatic mutations in the p53 tumor suppressor gene in rheumatoid arthritis synovium. Proc Natl Acad Sci USA 1997;94:10895-900.
21) Williams WV, VonFeldt JM, Ramanujam T, Weiner DB. Tyrosine kinase signal transduction in rheu- matoid synovitis. Semin Arthritis Rheum 1992;
21:317-29.
22) Lafyatis R, Thompson NL, Remmers EF, Flanders KC, Roche NS, Kim SJ, et al. Transforming growth factor-beta production by synovial tissues from rheumatoid patients and streptococcal cell wall arthritic rats. Studies on secretion by synovial fibroblast-like cells and immunohistologic localiza- tion. J Immunol 1989;143:1142-8.
23) Lunec J, Blake DR, McCleary SJ, Brailsford S, Bacon PA. Self-perpetuating mechanisms of immunoglo- bulin G aggregation in rheumatoid inflammation. J Clin Invest 1985;76:2084-90.
24) Desrochers PE, Weiss SJ. Proteolytic inactivation of alpha-1-proteinase inhibitor by a neutrophil me- talloproteinase. J Clin Invest 1988;81:1646-50.
25) McNeil JD, Wiebkin OW, Betts WH, Cleland LG.
Depolymerisation products of hyaluronic acid after exposure to oxygen-derived free radicals. Ann Rheum Dis 1985;44:780-9.