Corallococcus와 Myxococcus 속 점액세균 균주들에 의한 항균 물질의 생산
신혜진, 윤진권, 안동주, 조경연*
호서대학교생명공학과점액세균은행
Received : October 29, 2012 / Accepted : January 3, 2013
서 론
그람음성토양세균인점액세균은다양한생리활성물질을 생산하여이들로부터 100종류의기본구조를갖는 500여물 질들이탐색되었다[5, 13, 19]. 현재 3아목, 7과, 22속 47종이 알려져있는데[2, 15], Corallococcus, Myxococcus, Nanno- cystis, Sorangium 속에속한균주들이다른종류들에비해 상대적으로많은수가분리되었다[5]. 점액세균에서발견된 대부분의생리활성물질은 Sorangium과 Myxococcus 속균 주들에서분리되었으며, Corallococcus와 Nannocystis 속균 주들에서는극소수의물질들만이발견되었다[5, 20]. 이러한 이유로인해이들은다른점액세균종류에비해생리활성물 질생산능력이좋지않은것으로알려져왔다. 하지만최근 의유전체분석결과에의하면 Corallococcus 속균주도최 소한 13개의폴리케타이드또는폴리케타이드와비리보솜 펩티드의 혼성체 생합성효소 유전자군을 가지고 있어
Myxococcus와 Sorangium 속균주들에필적하는많은수의 생리활성물질을생산할수있는능력을가지고있는것으로 추정된다[8, 21]. 실제 Corallococcus 속균주들은다양한 생리활성물질을 생산하는 Myxococcus 속 균주들과 16S
rRNA 유전자염기서열이 3% 이상차이가나지않아같은
속으로도분류할수있을만큼계통적으로매우가깝다[3].
Myxococcus 속균주들이 생산하는생리활성물질은 16종
이상이알려져있다[16]. Corallococcus 속균주들이생산 하는 것으로알려진생리활성물질들로는 corallopyronin, pyrrolnitrin, myxothiazol, myxovalargin이있는데[16], 이 중 myxothiazol[4]과 myxovalargin[11]은 Myxococcus 속에 서최초발견된물질이며, Pseudomonas pyrrocinia에서최
초로 발견된 물질인 pyrrolnitrin도 점액세균 중에서는
Myxococcus 속에서처음발견되었다[6]. Corallopyronin만이 Corallococcus 균주에서최초로발견된물질이다[10]. 본연 구에서는국내에서분리한 Corallococcus와 Myxococcus 속 균주들을대상으로배양추출물을제조하여항균활성을비교 하였고, Staphylococcus aureus와 Pseudomonas aeroginosa 에항세균활성을보인 C. coralloides 균주들의항세균물질 생산조건을조사하였다.
Production of Antimicrobial Substances by Strains of Myxobacteria Corallococcus and Myxococcus. Shin, Hyejin, Jinkwon Youn, Dongju An, and Kyungyun Cho*. Myxobacteria Bank, Department of Biotechnology, Hoseo University, Asan 336-795, Korea
We prepared culture extracts of 174 Corallococcus and 207 Myxococcus strains isolated in Korea, and compared their antimi- crobial activity against Candida albicans, Pseudomonas aeruginosa, and Staphylococcus aureus. The percentage of strains showing antifungal activity was lower in Corallococcus (7.5% [13 of the 174 strains]) than in Myxococcus (51.7% [107 of the 207 strains]). However, the percentage of strains exhibiting antibacterial activity was higher in Corallococcus (12.1% [21 strains]) than in Myxococcus (1% [2 strains]). The culture extracts of 6 Corallococcus strains inhibited both P. aeruginosa and S. aureus and displayed similar high-performance liquid chromatography chromatograms, although the shapes of their fruiting bodies were dissimilar. The rate of production of antibacterial substances was the highest when the strains were cultured in CYS medium for more than 6 days.
Keywords: Myxobacteria, antimicrobial activity, Corallococcus, Myxococcus
*Corresponding author
Tel: +82-41-540-5627, Fax: +82-41-548-6231 E-mail: [email protected]
실험재료 및 방법
사용 균주
Corallococcus와 Myxococcus 속균주들은모두호서대학 교점액세균은행에서직접분리하여보유하고있는균주들 을 사용하였다. P. aeruginosa ATCC 10145, S. aureus ATCC 25923, Candida albicans ATCC18804는 한국생명 공학연구원생물자원센터에서구입하였다.
배지 및 배양조건
C. coralloides의일반적인배양에는 VY/3 배지를사용하 였으며, 필요에따라서 CY[17] 또는 CYE 배지[7]를사용하 였다. 배양추출물제조를위해서는 CYS 배지를사용하였 다. VY/3 배지는 VY/2 배지[17]를 변형시킨 배지로 0.5%
baker's yeast, 0.1% CaCl2·2H2O, 10 mM 3-[N-morpholino]
propanesulfonic acid (MOPS) (pH 7.6), 0.5µg/ml cyanocobala- mine을함유하고 있다. CYS 배지는 0.5% casitone, 0.1%
yeast extract, 0.3% soluble starch, 0.1% MgSO4·7H2O, 0.05%
CaCl2, 50 mM 4-(2-hydroxyethyl)-1-piperazineethanesulfonic acid (HEPES) (pH 7.6), 0.1% 미량원소용액, 0.5µg/ml cya- nocobalamine을 첨가하였으며, 미량원소용액은 1리터당 100 mg MnCl2·4H2O, 20 mg CoCl2, 10 mg CuSO4, 10 mg Na2MoO4·2H2O, 20 mg ZnCl2, 5 mg LiCl, 5 mg SnCl2· 2H2O, 10 mg H3BO3, 20 mg KBr, 20 mg KI, 8 g EDTA Na-Fe3+ salt (trihydrate)를첨가하였다. 자실체형성유도를
위해서는 WCE 배지를 사용하였다. WCE 배지는 10 mM
MOPS, 0.1% CaCl2·2H2O, 1.5% 한천으로구성된고체평판 배지표면에살아있는대장균농축액 20µl를올려놓은배지 이다. P. aeruginosa와 S. aureus는 NA 배지에서배양하였 으며, C. albicans는 YM 배지에서배양하였다. NA 배지는 0.3% beef extract와 0.5% peptone을첨가하였으며, YM 배 지는 0.3% yeast extract, 0.3% malt extract, 0.5% peptone, 1% glucose로구성되어있었다.
균주들의 형태적 특성 조사
점액세균단일세포의형태관찰을위해서는 VY/3 배지에
서 3일동안 배양한영양세포들을 Nikon ECLIPSE E600 위상차현미경을사용하여관찰하였으며, 점액세균의자실 체 형태는 WCE 배지에서 7일간 배양한자실체를 Nikon SMZ1000 입체현미경을사용하여관찰하였다. 사진은 Nikon
DS-Fi1 디지털카메라를사용하여촬영하였다.
배양추출물의 제조
C. coralloides 균주들을 Amberlite XAD16 (Sigma, USA) 2 g이들어있는 50 ml CYS에서 5일간진탕배양한후,
레진과세포를회수하여증류수로 2회세척하였다. 50% 아 세톤으로 2회추출하고, 다시 100% 아세톤으로 2회추출하 여추출액을모았다. 추출물을건조시킨후, 1 ml의 DMSO 에녹여보관하였다. HPLC 분석을위해서는레진만을증류
수로세척하고 50% 아세톤과 100% 아세톤으로추출하였
다. 회전증발기를사용하여아세톤만을제거한다음, 남은 수용액을에틸아세테이트로추출하였고, 건조시킨후, 80%
메탄올 1 ml에용해하였다.
항균 활성 조사
C. albicans, P. aeruginosa 또는 S. aureus가도말된평 판배지위에배양추출액 10µl를함유한 6 mm 종이디스크 (Advantec MFS, Japan)를올려놓았다. 대조구로는 DMSO 를함유한종이디스크를사용하였다. 32oC에서 2일간배양한 후, 생장저지환의지름을측정하여항균활성을측정하였다. HPLC 분석
배양추출물의 HPLC 분석은 Zorbax SB-C18 컬럼(4.6 mm
×150 mm, 5µm)을 장착한 Agilent 1260 VL Infinity Series HPLC 시스템을 사용하였다(Agilent Technologies, USA). 이동상 A와 B는각각 0.1% formic acid를함유한증 류수와 acetonitrile이었으며, 유속은 0.2 ml/min이었고, 처 음 30분간 5~100% B 용매기울기, 30~40분에는 100% B 용 매, 그리고 40~50분에는 5% B 용매로용리하였다. 용리액 은 254 nm에서검출하였다.
결과 및 고찰
Corallococcus 및 Myxococcus 속 균주들의 배양추출 물 제조와 항균 활성 측정
호서대학교점액세균은행은전국 2,015지점에서채취한토 양시료로부터분리한 2,672 균주의야생점액세균을보유하 고있다[9, 14]. 16S rRNA 유전자염기서열및자실체형태 를기반으로한균주동정에의하면, 보유균주들의 40.9%가 Myxococcus 속 균주들이며, 28.0%가 Corallococcus 속, 23.5%가 Sorangium 속, 그리고나머지 7.7%가다른속에속 하는점액세균균주인것으로분석되었다.
Gerth 등은점액세균에서탐색된생리활성물질과생산균
주에대한정보를바탕으로 Myxococcus 속과 Sorangium 속 균주들이다양한생리활성물질을생산하는반면, Corallococcus 속균주들은생리활성물질을거의생산하지않음을보고하 였다[5]. 국내에서분리한 Corallococcus 속균주들도동일한 양상을보이는지알아보기위하여, 국내토양시료에서분리한 Corallococcus 174속균주와 Myxococcus 207속균주들을배 양하여배양추출물을제조하고, C. albicans, S. aureus, P.
aeruginosa에대한 생장저지활성을측정하였다. 그결과, 174 Corallococcus 속균주중 7.5%에해당하는 13균주가 C.
albicans의성장을억제하는항진균활성을보였으며, 12.1%
에해당하는 21균주가그람음성세균인 S. aureus에, 3.4%
에해당하는 6균주가그람양성세균인 P. aeruginosa에항 세균활성을보였다(Table 1). S. aureus에활성을보인 6균 주들은모두 P. aeruginosa에도활성을보였지만, C. albicans 에활성을보인 13균주들은 S. aureus과 P. aeruginosa에활 성을보이지않았다. 따라서 174균주중 19.5%에해당하는
34균주가항진균또는항세균활성을보였다.
Myxococcus 속균주들의경우에는 207 균주중 51.7%에 해당하는 107균주가 C. albicans의성장을억제하는항진균 활성을보였으며, 1%에해당하는 2균주가그람음성세균인 S. aureus에항세균활성을보였다. P. aeruginosa에항세균 활성을보인균주들은없었다(Table 1). S. aureus에활성을 보인 2균주중 1균주가 C. albicans에도활성을보였으므로 207균주중 52.2%에해당하는 108균주가항진균또는항세 균활성을보였다. 이는 Corallococcus 속균주들에비해 2.7 배 높은 비율이다. 이러한 결과는 국내 토양에서 분리된
Corallococcus 속균주들도국외분리균주들과마찬가지로
Myxococcus 속균주들에비해활성물질생산비율이낮음을
보여준다. 이러한특성으로인해그동안 Corallococcus에서 의신규물질발견이 Myxococcus에비해상대적으로낮은것 으로사료된다. 하지만최근의유전체분석결과에의하면 Corallococcus 균주들도 Myxococcus 속균주들과유사한숫 자의이차대사산물생합성유전자를가지고있어많은수의 생리활성물질을생산할수있는능력을가지고있는것으로 나타났다[8]. 따라서지금까지 Corallococcus 균주들에서발
견된물질이극소수였던이유는물질탐색을위해사용된배 양조건에서물질이극소량생산되었거나아예생산이되지 않아활성조사에서검출되지않았기때문인것으로추정된다. 개별활성에서는다른양상을보였는데, 항진균활성의경 우에는 Corallococcus 속균주들(7.5%)이 Myxococcus 균주 들(51.7%)에비해낮은비율을보였지만, 항세균활성의경 우에는 Corallococcus 속 균주들(12.1%)이 Myxococcus 속 균주들(1%)에비해높은비율을보였다. 특히 Myxococcus 속 균주들에서는 P. aeruginosa에대한저해활성을전혀 보 이지 않았지만, Corallococcus의 경우에는 6균주가 P.
aeruginosa에대한성장저해활성을보였다(Table 1).
이후의실험에서는 Corallococcus 속균주들에의한물질 생산특성을조사하기위하여 S. aureus와 P. aeruginosa에 활성을 보인 6균주 KYC4006, KYC4010, KYC4012, KYC4021, KYC4023, KYC4046를대상으로자실체 형성, 배지및배양시간에따른물질생산을조사하였으며, 배양추
출물들을 HPLC로비교분석하였다.
S. aureus와 P. aeruginosa에 성장저지 활성을 보인 C.
coralloides 균주들의 특성
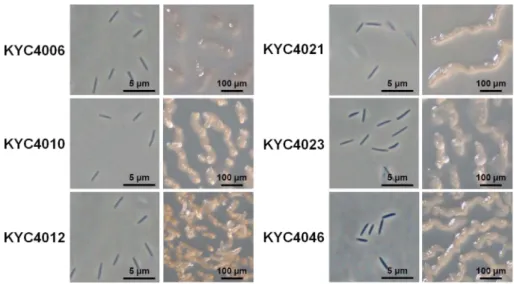
S. aureus와 P. aeruginosa에활성을보인 6균주들을자실 체유도배지인 WCE 배지에서 7일간배양하였을때, 6균주 들은 모두 구불구불한 산등성이 형상의 전형적인 C.
coralloides 자실체를형성하였으며, 일부균주들은산등성이 위에 뿔과같은 구조물을형성하기도 하였다(Fig. 1). 16S rRNA 유전자서열도 C. coralloides의표준균주인 DSM2259 (Accession number DQ768120)와 99.7% 이상동일하였다. 따 라서 6균주는모두 C. coralloides인것으로확인되었다. 하 Table 1. Antimicrobial activity of the culture extracts of Corallococcus and Myxococcus strains.
Classification Number of strains
Number of strains showing growth inhibitory activity against:
Candida albicans Staphylococcus aureus Pseudomonas aeruginosa
Corallococcus coralloides 154 13 (8.4) 19 (12.3) 6 (3.8)
Corallococcus exiguus 20 0 2 (10.0) 0
Subtotal 174 13 (7.5) 21 (12.1) 6 (3.4)
Myxococcus fulvus 58 31 (53.4) 2 (3.4) 0
Myxococcus macrosporus 5 2 (40.0) 0 0
Myxococcus stipitatus 46 27 (58.6) 0 0
Myxococcus virescens 28 18 (64.2) 0 0
Myxococcus xanthus 70 29 (41.4) 0 0
Subtotal 207 107 (51.7) 2 (1.0) 0
Total 381 120 (31.5) 23 (6.0) 6 (1.6)
(): Percentage of strains showing antimicrobial activity.
Culture extracts of 174 Corallococcus and 207 Myxococcus strains were prepared, and their antimicrobial activity against Pseudomo- nas aeruginosa ATCC 10145, Staphylococcus aureus ATCC 25923, and Candida albicans ATCC 18804 was examined.
지만균주들간에는자실체의형태와색에서차이를보이는
데, KYC4010은끝이산등성이위에동그란뿔을형성한반
면, KYC4012는끝이뾰족한뿔을형성하였으며, KYC4006
은 자실체 색이 다른 균주와 차이를 보였다(Fig. 1). 16S
rRNA 유전자서열에서도차이를보였다. 6균주들은모두지
역적으로 다른 장소에서 분리한 균주들로, KYC4006,
KYC4012는강원도에서채취한토양에서분리하였으며, 다
른 4균주는각각경남, 인천, 전북, 충북에서채취한토양에 서분리하였다(Table 2). 이러한결과들을바탕으로 6균주는 모두다른균주들인것으로사료되었다.
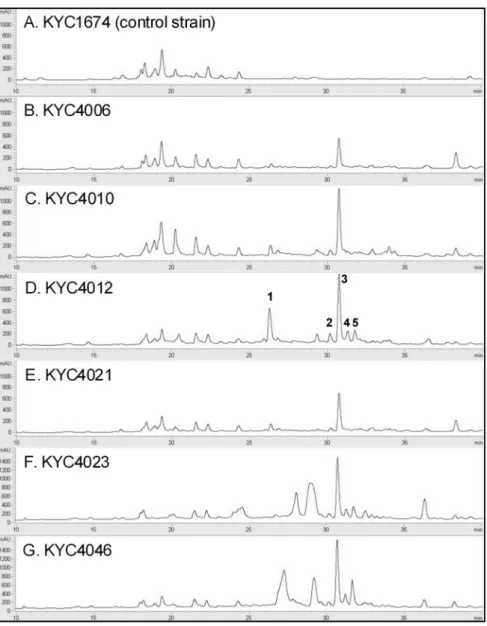
C. coralloides가 생산하는 항세균 물질의 HPLC 분석 S. aureus와 P. aeruginosa에활성을보인 6균주들이활성 을전혀보이지않은 C. coralloides 균주 KYC1674와물질 생성에서어떤차이를보이는지조사하기위하여이들의배
양추출물을 Zorbax SB-C18 컬럼을장착한 HPLC를사용하 여비교분석하였다. 7개균주들의배양추출물을 HPLC로 전개하였을때, 25분이전에서는대체로유사한크로마토그 램을 보였다(Fig. 2). 하지만 25분 이후에는 활성이 없는
KYC1674와활성이있는 6균주들의배양추출물들사이에눈
에 띄는 차이가 관찰되었다. 특히, 26.62, 30.14, 30.71,
31.23, 31.71분에서검출된피크들은대부분의활성이있는
균주들에서검출되었지만활성이없는 KYC1674의추출물 에서는검출되지않았다. 이후실험을위해차이를보이는 피크각각에번호를매겨구분하였다(Fig. 2).
분석된피크중어떤피크가항세균활성을보이는지조 사하기위하여 6균주의하나인 KYC4012의배양추출물을대
상으로 HPLC 전개를실시하여 5개피크의분획을받아농
축한다음, S. aureus와 P. aeruginosa를대상으로활성을측 정하였다. 그결과, 각각 30.14, 30.71, 31.23, 31.71분의머 Fig. 1. Vegetative cells and fruiting bodies of Corallococcus coralloides strains.
Table 2. C. coralloides strains producing antimicrobial substances that inhibit the growth of S. aureus and P. aeruginosa.
Strains Geographical origin Size of growth inhibition zone (mm)
C. albicans S. aureus P. aeruginosa
KYC1674 Gurye, Jeonnam - - -
KYC4006 Gangneung, Gangwon - 18 10
KYC4010 Jecheon, Chungbuk - 19 11
KYC4012 Jeongseon, Gangwon - 20 10
KYC4021 Sancheong, Gyeongnam - 20 11
KYC4023 Imhak, Incheon - 20 11
KYC4046 Jeonju, Jeonbuk - 20 10
The culture extract from each strain was absorbed on a 6-mm paper disc, and the disc was placed on agar plates poured with cul- tures of P. aeruginosa ATCC 10145, S. aureus ATCC 25923, and C. albicans ATCC 18804 cultures. The plates were incubated at 32oC for 2 days. After incubation, the clear zones indicating growth inhibition were measured.
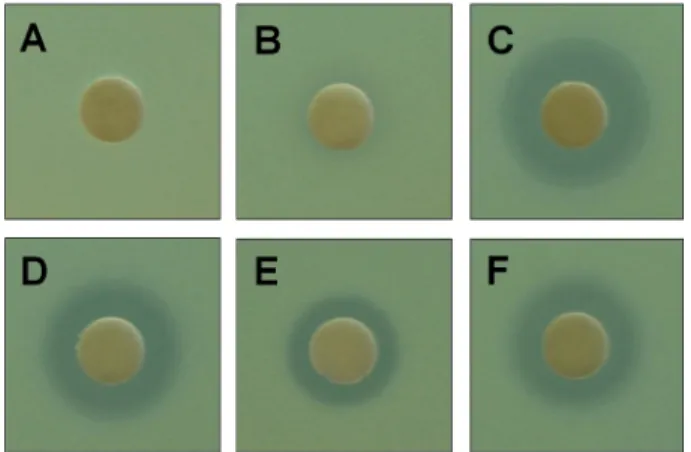
무름시간을갖는피크-2, 3, 4, 5 분획이 S. aureus에성장
저해활성을보였으며, 26.62분의머무름시간을갖는피크
-1 분획은활성을보이지않았다(Fig. 3). 한편, 다섯피크모 두는 P. aeruginosa에저해활성을보이지않았다.
C. coralloides에서발견된생리활성물질로는 corallopyronin, pyrrolnitrin, myxothiazol, myxovalargin 등이 있는데, myxothiazol과 pyrrolnitrin은전자전달을저해하는항진균 물질이며[6, 18], corallopyronin은 RNA 중합효소의작용을
저해하는항세균물질이고[1], myxovalargin은단백질합
성을 저해하는 항세균 물질로 알려져 있다[12]. 그런데
myxovalargin이 S. aureus와 P. aeruginosa에 모두활성 을보이는반면, corallopyronin은 S. aureus에만활성을보
이고 P. aeruginosa에는 활성을보이지않는 것으로알려
져있다. 하지만현재의결과로는 KYC4012의배양추출물
에서활성을보이는물질이 corallopyronin인지는확인되지 않았다.
Fig. 2. Chromatograms showing the high-performance liquid chromatography (HPLC) results for the culture extracts of the selected C. coralloides strains.
HPLC was performed using the Agilent 1260 VL Infinity Series HPLC System with a Zorbax SB-C18 column (4.6 mm × 150 mm; 5µm).
The mobile phases A and B were water and acetonitrile with 0.1% formic acid, respectively, and the gradient elution at a flow rate of 0.2 ml/min was performed as follows: 0-30 min for 5-100% B (linear gradient), 30-40 min for 100% B (isocratic gradient), and 40-50 min for 5% B (isocratic gradient). The elutes were analyzed using a variable wavelength detector at 254 nm. The peaks com- mon to the chromatograms of the extracts from all the 6 selected strains were numbered.
배지에 따른 항세균 물질의 생산
점액세균의배양에서많이사용하는배지로는 CYE, CY, VY/2 등이있다. CYE는 1%의 casitone과 0.5%의효모추출 물을함유한배지로고농도영양분에적응된용균성점액세 균의배양에사용하는데[7], 대부분의균주들은이배지에서 다세포자실체를생성하지않는다. CY는 0.3%의 casitone과 0.1%의 yeast extract를함유한배지[17]로 Corallococcus를 포함한용균성야생점액세균의배양에사용하며, 일정기간 성장후일부균주들은자실체를생성하기도한다. VY/2는 효모를먹이로일반적인야생점액세균의분리및배양에사 용하는배지[17]로대부분의점액세균들은 VY/2 배지에서
일정기간성장한후, 다세포자실체를형성한다. VY/3 배지
는 VY/2에 10 mM MOPS를첨가한배지이다.
C. coralloides 균주들에의한항세균물질생산이최적인 배지를선별하기위하여 CYE, CY, VY/3 배지에서 KYC4012 을배양하고, 배양추출물을제조한다음, HPLC 분석을통 해물질피크의크기를비교하였다. 그결과, VY/3에서는 1~5 번피크물질생성이모두관찰되었으나, CYE와 CY에서는 이들피크들이전혀검출되지않았다(Table 3). KYC4023과 KYC4046을대상으로배양과, S. aureus를대상으로하는생 리활성조사에서도동일한결과를얻었다. 이러한결과의원 인에대한설명중하나는 Corallococcus속균주들이 CYE 과같이영양생장만을위한배지에서는항세균물질을생산
하지않지만, VY/3와같이영양생장을지원하다가후기에자
실체를형성하게하는배지에서는항세균물질을생산한다 는것이다.
이러한설명과이전에생리활성물질생산에사용되었던배
지조성을기반으로 CYS 배지를디자인하고 KYC4012에의
한항세균물질의생산을조사하였다. 그결과, CYS 배지에 서는 VY/3 배지에서보다피크에따라 2~8배많은물질을생 산하였다(Table 3). CYS 배지는 0.5% casitone과 0.1%
yeast extract 외에도 0.3% soluble starch, 0.1% MgSO4· 7H2O, 0.05% CaCl2, 50 mM HEPES, 0.1% 미량원소용액, 0.5µg/ml cyanocobalamine을함유하고있다. Corallococcus 와 Myxococcus속균주들은 CYS 배지에서배양초기에왕성 히영양생장하며, 후기에는자실체를형성하였다.
배양시간에 따른 항세균 물질의 생산
Corallococcus속균주에의한항세균활성물질의생산이
배양시간에따라어떻게달라지는지조사하기위하여 CYS 배지에 KYC4012 균주를접종하고, 2일, 4일, 6일, 8일, 10일
동안배양한다음, XAD-16 레진을회수하여배양추출물을
제조하였다. 그리고 HPLC 분석을통하여배양시간에따른 3번피크(30.71분)의변화를조사하였다. 3번피크물질의양 은 6일째까지계속증가하였으며, 이후에는정체되는양상 Fig. 3. Antimicrobial activity of the HPLC peak fractions.
The culture extract from C. coralloides KYC4012 was separated by performing HPLC, and the antimicrobial activity of the peak fractions shown in Fig. 2 against S. aureus ATCC 25923 was assayed. (A) Negative control, (B) Peak-1, (C) Peak-2, (D) Peak-3, (E) Peak-4, (F) Peak-5.
Table 3. Effect of culture medium on the production of antimicrobial substances in C. coralloides KYC4012.
Peak Retention time (min)
Relative peak area (%)
CYE CY VY/31 CYS
Peak-1 26.62 0.0 0.0 100.0 589.9
Peak-2 30.14 0.0 0.0 100.0 222.4
Peak-3 30.71 0.0 0.0 100.0 388.9
Peak-4 31.23 0.0 0.0 100.0 331.1
Peak-5 31.71 0.0 0.0 100.0 823.6
1: Control
C. coralloides KYC4012 was cultured in CYE, CY, VY/3, and CYS broths and the culture extracts prepared from each cul- ture were analyzed using HPLC.
Fig. 4. Production of the peak-3 substance by C. coralloides KYC4012.
Production of the peak-3 substance was expressed as relative HPLC peak area of peak-3.
을보였다. 따라서최대수율을얻기위해서는 6일간배양하 는것이가장적절한것으로나타났다. Corallococcus 속균 주들은물질생산능력에비해탐색된생리활성물질이극소 수이다. 본연구에서얻은결과들은향후 Corallococcus 속 균주들로부터신규생리활성물질을탐색하는데기반이될 것으로기대한다.
요 약
국내토양시료에서분리한 174 Corallococcus 속균주와 207 Myxococcus 속균주들의배양추출물을제조하고 Can- dida albicans, Pseudomonas aeruginosa, Staphylococcus
aureus에대한항균활성을비교하였다. 항진균활성을보이
는균주의비율은 Corallococcus가 7.5%(174균주중 13균 주)로 51.7%(207균주중 107균주)를보인 Myxococcus에비 해 낮았다. 하지만, 항세균 활성을 보인 균주의 비율은 Corallococcus가 12.1%(21균주)로 1%(2균주)를보인 Myxo- coccus에비해상대적으로높았다. Corallococcus 균주중 P.
aeruginosa, S. aureus에 모두 활성을 보이는 6 C. coral-
loides 균주들은자실체형성에서는차이를보였지만항세균
활성을나타내는배양추출물의 HPLC 크로마토그램은유사
하였는데, 이들이생산하는항세균물질의생산은 CYS 배지
에서 6일이상배양할때가장높았다.
Acknowledgments
This study was supported by the Basic Science Research Pro- gram through the National Research Foundation of Korea (NRF), funded by the Ministry of Education, Science and Technology (2010-0022869).
References
1. Belogurov, G. A., M. N. Vassylyeva, A. Sevostyanova, J. R.
Appleman, A. X. Xiang, R. Lira, S. E. Webber, S. Klyuyev, E.
Nudler, I. Artsimovitch, and D. G. Vassylyev. 2009. Transcription inactivation through local refolding of the RNA polymerase structure. Nature 457: 332-335.
2. Dawind, W. 2000. Biology and global distribution of myxobac- teria in soil. FEMS Microbiol. Rev. 24: 403-427.
3. Garcia, R., K. Gerth, M. Stadler, I. J. Jr. Dogma, and R. Müller.
2010. Expanded phylogeny of myxobacteria and evidence for cultivation of the 'unculturables'. Mol. Phylogenet. Evol. 57:
878-887.
4. Gerth, K., H. Irschik, H. Reichenbach, and W. Trowitzsch. 1980.
Myxothiazol, an antibiotic from Myxococcus fulvus (Myxo- bacterales). I. Cultivation, isolation, physico-chemical and bio- logical properties. J. Antibiot. 33: 1474-1479.
5. Gerth, K., S. Pradella, O. Perlova, S. Beyer, and R. Müller. 2003.
Myxobacteria: proficient producers of novel natural products with carious biological activities-past and future biotechnologi- cal aspects with the focus on the genus Sorangium. J. Bio- technol. 106: 233-253.
6. Gerth, K., W. Trowitzsch, V. Wray, G. Höfle, H. Irschik, and H.
Reichenbach. 1982. Pyrrolnitrin from Myxococcus fulvus (My- xobacterales). J. Antibiot. 35: 1101-1103.
7. Hagen, D. C., A. P. Bretscher, and D. Kaiser. 1978. Synergism between morphogenetic mutants of Myxococcus xanthus.
Dev. Biol. 64: 284-296.
8. Huntley, S., Y. Zhang, A. Treuner-Lange, S. Kneip, C. W. Sensen, and L. Søgaard-Andersen. 2012. Complete genome sequence of the fruiting myxobacterium Corallococcus coralloides DSM 2259. J. Bacteriol. 194: 3012-3013.
9. Hyun, H., J. Chung, H. Lee, J. Youn, C. Lee, D. Kim, and K.
Cho. 2009. Isolation of cellulose-degrading myxobacteria Sor- angium cellulosum. Korean J. Microbiol. 45: 48-53.
10. Irschik, H. and H. Reichenbach. 1985. The mechanism of action of myxovalargin A, a peptide antibiotic from Myxococ- cus fulvus. J. Antibiot. 38: 1237-1245.
11. Irschik, H., K. Gerth, T. Kemmer, H. Steinmetz, and H. Reichen- bach. 1983. The myxovalargins, new peptide antibiotics from Myxococcus fulvus (Myxobacterales). I. Cultivation, isolation, and some chemical and biological properties. J. Antibiot. 36: 6- 12.
12. Irschik, H., R. Jansen, G. Höfle, K. Gerth, and H. Reichen- bach. 1985. The corallopyronins, new inhibitors of bacterial RNA synthesis from myxobacteria. J. Antibiot. 38: 145-152.
13. Kim, Y. S., W. C. Bae, and S. J. Back. 2003. Bioactive sub- stances from myxobacteria. Korean J. Microbiol. Biotechnol.
31: 1-12.
14. Park, S., B. Lee, J. Kim, C. Lee, E. Jang, and K. Cho. 2004.
Isolation and characterization of bacteriolytic wild myxobacte- ria. Korean J. Microbiol. Biotechnol. 32: 218-223.
15. Reichenbach, H. 2005. Myxococcales. pp. 1059-1144. In Brenner, D. J., N. R. Krieg, J. T. Staley, and G. M. Garrity (ed.), Bergey's Manual of Systematic Bacteriology, 2nd ed. Bergey's Manual Trust, East Lansing, MI, USA.
16. Reichenbach, H. and G. Hofle. 1999. Myxobacteria as produc- ers of secondary metabolites. pp. 149-179. In Grabley S. and R. Thiericke (ed.), Drug Discovery from Nature, Springer Ver- lag, Berlin, Germany.
17. Reichenbach, H. and M. Dworkin. 1992. The myxobacteria, pp. 3416-3487. In Balows, A., H. G. Trper, M. Dworkin, W.
Harder, and K.-H. Schleifer (ed.), The Prokaryotes, 2nd ed., vol. IV, Springer Verlag, New York, USA.
18. Thierbach, G. and H. Reichenbach. 1981. Myxothiazol, a new antibiotic interfering with respiration. Antimicrob. Agents Chemother. 19: 504-507.
19. Weissman, K. J. and R. Müller. 2009. A brief tour of myxobac- terial secondary metabolism. Bioorg. Med. Chem. 17: 2121- 2136.
20. Weissman, K. J. and R. Müller. 2010. Myxobacterial second- ary metabolites: bioactivities and modes-of-action. Nat. Prod.
Rep. 27: 1276-1295.
21. Wenzel, S. C. and R. Müller. 2009. The impact of genomics on the exploitation of the myxobacterial secondary metabolome.
Nat. Prod. Rep. 26: 1385-1407.