고병원성 조류인플루엔자 바이러스의 생태 및 국내외 발생 현황 신유진 Page 1 / 10 BRIC View 2018-T04
고병원성 조류인플루엔자 바이러스의 생태 및
국내외 발생 현황
신 유 진
생물학·생명공학 프리랜서 컨설턴트 E-mail: yujin1023@hotmail.com 요약문 인플루엔자 A 바이러스는 지속적인 변이(mutation)와 재편성(reassortment)을 통해 인체감염을 증가시키는 위험 인자이다. 특히 고병원성 조류바이러스의 인체감염은 더욱 치명적이다. 고병 원성 조류인플루엔자 바이러스(avian flu)는 5번째 대유행(pandemic)을 일으킬 가능성이 가장 높은 인플루엔자 A 바이러스이다. 국내외 발생한 고병원성 조류인플루엔자 바이러스는 A/H5N1, A/H7N9, A/H5N8, A/H5N6이다. 국내에서는 인체 감염의 발생 사례는 없고 국외에서 는 확진 환자 A/H7N9(1.622명), A/H5N1(860명), A/H5N6(18명) 순서로 발생하였고 사망률은 A/H5N6(55.6%), A/H5N1(52.8%), A/H7N9(38.2%) 순서이다. A/H5N8은 국내 야생 조류에서 첫 발견 이후 미국, 유럽 등에서도 발견되고 현재까지 인체 감염 사례는 없다. 지속적인 주의, 관 리, 감독을 통해 대유행의 발생을 차단해야 할 것이다. 본 글에서는 고병원성 조류인플루엔자 바이러스의 생태 및 국내외 발생 현황에 대해 논의하고자 한다.Key Words: 고병원성 조류인플루엔자 바이러스, A/H5N1, A/H7N9, A/H5N8, A/H5N6
목 차
1. 서론 1.1 인플루엔자 A 바이러스 1.2 조류인플루엔자 바이러스 2. 본론 2.1 고병원성 조류인플루엔자 바이러스의 생태 2.2 고병원성 조류인플루엔자 바이러스의 국내외 발생 현황 BRIC View 동향리포트고병원성 조류인플루엔자 바이러스의 생태 및 국내외 발생 현황 신유진 Page 2 / 10 3. 결론 4. 참고문헌
1. 서론
1.1
인플루엔자 A 바이러스
인플루엔자 바이러스는 오소믹소비리대(Orthomyxoviridae)과에 속하는 단일 가닥 음성 RNA 유전자 바이러스로 항원형에 따라 A, B, C, D형으로 분류된다. 인플루엔자 A 바이러스는 대유행 독감의 원인이며 사람과 가금류를 감염시킨다. A형과 함께 B형은 주로 사람에게 감염을 일으키며 계절성 유행성(epidemic) 독감의 원인이다. 그에 비해, C형은 약한 호흡기 질환을 유발하여 계절성 유행성 독감을 유발하지 않는다. 2011년 첫 발견된 D형의 경우 소(cattle)에서 발견된 바이러스 형으로 인체 감염은 보고된 바 없다[1]. 인플루엔자 A 바이러스는 표면당단백질 헤마글루티닌(Hemagglutinin; HA1~HA18)과 뉴라미니데이즈(Neuraminidase; NA1~NA11)의 조합에 의해 다양한 아형(subtype)으로 구분된다. 또한, 각각의 아형은 사람, 가금류, 돼지 또는 박쥐 등 발견되는 종의 종류가 다르다[2](표 1). 표 1 헤마글루티닌과 뉴라미니데이즈 아형에 따른 다양한 숙주의 종 표면당단백질 종 아형 헤마글루티닌 (HA) 사람 HA1/2/3/5/6/7/9/10 가금류 HA1~16 돼지 HA1/2/3/4/5/9 박쥐 HA17/18 다른 동물 HA3/4/7 뉴라미니데이즈 (NA) 사람 NA1/2/6/7/8/9 가금류 NA1~9 돼지 NA1/2 박쥐 NA10/11 다른 동물 NA7/8 4번의 대유행 바이러스가 1918년부터 2009년 사이 발생하였다(그림 2). 가장 많은 사망자를 발생시킨 스페인 독감(A/H1N1) 이후 아시아 독감(A/H2N2), 홍콩 독감(A/H3N2)을 거쳐 2009년 돼지 독감(A/H1N1)pdm09은 우리나라에서도 260명의 사망자를 발생시켰다[3;4]. 5번째 대유행을 일으킬 가능성이 가장 높은 조류인플루엔자 바이러스의 발생은 지속적으로 보고되고 있다. 2003년부터 발견되기 시작한 조류인플루엔자 A/H5N1는 야생 조류와 가금류에서 발병하여 사람에게 전파되어 2017년 9월까지 총 16개국에서 454명의 사망자를 발생시켰다[5]. 2013년 중국에서 발생한 조류인플루엔자 A/H7N9의 경우는 2017년 현재까지 599명의 사망자를 발생시켰다[6].고병원성 조류인플루엔자 바이러스의 생태 및 국내외 발생 현황 신유진 Page 3 / 10
그림 1. 대유행 인플루엔자 A 바이러스의 역사
1.2 조류인플루엔자 바이러스
조류인플루엔자 바이러스는 헤마글루티닌 항원에 의해 병원성이 결정되며 정도에 따라 고병원성(Highly Pathogenic Avian Influenza Virus)과 저병원성(Low Pathogenic Avian Influenza Virus) 으로 분류된다. 고병원성 조류인플루엔자 바이러스는 주로 H5와 H7 아형을 가지며 이들 중 일부가 고병원성으로 발현된다. A/H5N1은 대표적인 고병원성 바이러스로 사이토카인 스톰(cytokine storm)을 유발하여 급성 호흡기 감염을 일으키는 치사율 50% 이상인 치명적인 바이러스다[7]. A/H7N9는 급성 폐질환과 급성 호흡곤란 증후군(acute respiratory distress syndrome; ARDS)을 유발하는 고병원성 조류인플루엔자 바이러스로 38%의 치사율을 보인다[8]. 고병원성 조류인플루엔자 바이러스는 높은 폐사율과 심각한 산란율 저하를 유발하여 제1종 가축전염병으로 분류하고 있다. 그에 비해 H5, H7을 제외한 나머지 아형의 조합인 저병원성 바이러스(예: A/H1N1, A/H1N2, A/H3N2, A/H9N2 등)는 가금류의 호흡기나 소화기에서 증식하여 무증상이거나 경증을 일으킨다. 따라서 제 2종 가축전염병으로 분류된다[9; 표 2]. A/H9N2는 1998년부터 현재까지 지속적으로 발병하고 있는 저병원성 조류인플루엔자 바이러스로 중국, 방글라데시, 이집트에서 발견되었다. 2000년도에 가장 높은 발병률을 보였고 확진 환자 31명 중 1명의 사망자가 발생한 사망률이 3.2%으로 낮은 치사율을 보인다.
고병원성 조류인플루엔자 바이러스의 생태 및 국내외 발생 현황 신유진 Page 4 / 10 표 2. 병원성에 따른 조류인플루엔자 바이러스의 특징 특성 고병원성 저병원성 아형 H5, H7 H1~H16 증식부위 전신 국소(호흡기, 소장) 임상증상 전신적, 중증 무증상 혹은 경증 폐사율 높음 낮음 인체감염 수 A/H5N1: 확진860명 사망 454 명 A/H7N9: 확진 1,568명 사망 599명 A/H9N2: 확진 31명 사망 1명 분류 제1종 가축전염병 제2종 가축전염병
2. 본론
2.1 고병원성 조류인플루엔자 바이러스의 생태
인플루엔자 A 바이러스가 숙주를 감염시켜 증식하여 배출하는 복제 과정은 수용체 인식, 바이러스의 침투, RNA 합성(synthesis), 복제, 스플라이싱(splicing), 단백질 합성, 바이러스 조립(assembly), 배출(budding)의 순서로 이루어진다(그림 2). 표면당단백질 헤마글루티닌은 바이러스가 숙주를 감염시킬 때 초기 숙주 수용체(sialic acid (SA) receptor)에 부착함으로써 바이러스가 숙주 내로 침투(receptor-mediated endocytosis)하는데 중요한 역할을 한다. 반면에 뉴라미니데이즈는 SA와 표면당단백질의 결합을 끊어 증식한 바이러스가 세포 밖으로 배출하는데 중요한 역할을 한다.고병원성 조류인플루엔자 바이러스의 생태 및 국내외 발생 현황 신유진 Page 5 / 10 HA의 숙주 SA수용체는 사람의 경우 상부 기관지 인플루엔자 바이러스 수용체 α-2.6-linked SA를 하부 기관지의 경우 α-2.3-linked SA수용체에 친화성이 있다. 고병원성 조류인플루엔자 바이러스 A/H5N1의 경우 하부 기관지 수용체에 친화성이 있어 상부 기관지보다는 감염은 어렵지만 하부 기관지에 감염이 되면 치명적 결과를 초래한다[10]. 흥미롭게도 A/H7N9의 경우 상부와 하부 기관지 모두 친화성이 있어 A/H5N1에 비해 상부 기관지를 통한 감염이 비교적 용의하다[11]. 조류인플루엔자가 사람을 감염시킨 경우는 고농도의 바이러스의 노출에 의해 바이러스가 하부 기관지까지 도달한 경우이다. 감염된 가금류와 직접적인 접촉이나 배설물 및 분비물, 공기 중의 전파를 통해 조류인플루엔자는 사람을 감염시킬 수 있다. 현재까지 명확한 조류인플루엔자의 사람 간 전파 경로는 알려지지 않았으나, 동물실험을 통해 공기 중으로 전파 가능성이 보고되었다[11,12]. 사람간의 전파는 사람과 같이 두 종류의 기관지 수용체를 가진 돼지에게 A/H5N1 바이러스가 전파된 후 사람에게 감염된 경우의 가능성이 있다[13]. H7 바이러스는 H5의 경우보다 조류에서 병원성이 낮아서 지속적인 조류와의 접촉으로 인체 감염을 증가시킨다. 따라서 지속적인 변이로 고병원성 바이러스가 발견된다. 예를 들면 2013년 중국에서 처음 발견된 A/H7N9은 저병원성 조류인플루엔자 바이러스였으나 2017년의 인체감염자의 경우 고병원성 조류인플루엔자로 밝혀졌다[14]. A/H7N9 바이러스는 오리의 헤마글루티닌 유전자, 야생조류의 뉴라미데이즈 유전자 그리고 닭의 H9N2 바이러스의 6개의 내부 유전자가 재편성된 바이러스로 밝혀졌다[15;16] (그림 3).
그림 3. 다수의 유전자 재편성을 거듭한 A/H7N9 바이러스의 구조[출처: CDC: Transmission of influenza virues from animals to people]
고병원성 조류인플루엔자 바이러스의 생태 및 국내외 발생 현황 신유진 Page 6 / 10
2.2 고병원성 조류인플루엔자 바이러스의 국내외 발생 현황
2.2.1 국내 발생 현황
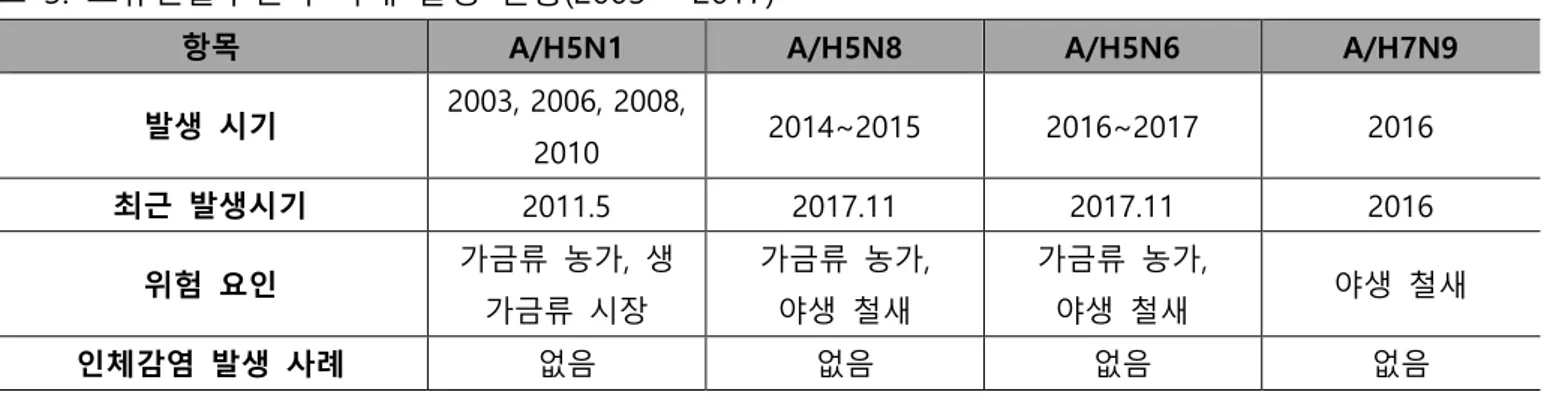
A/H5N1는 2003~2004년 국내에서 첫 발생한 이후 총 4번의 유행성 인플루엔자가 발생하였다. A/H5N1은 조류에서 고병원성을 보이며 높은 치사율의 원인이 되었다. A/H5N8은 국내 5번째 조류인플루엔자 유행성 인플루엔자로 2014년 한국에서 첫 발생하여 철새들에 의해서 2014~2015년 전 세계로 퍼졌다. 2013년 중국에서 가장 먼저 발견된 A/H5N6는 국내에서 2016년부터 2017년까지 양계장에서 발견 사례가 보고된다[17]. A/H7N9는 2016년 국내 야생 철새 분변에서 확인되었고 현재까지 가금 농가에서 발생한 적은 없다(표 3). 조류인플루엔자의 발병은 가금류 농가에 경제적으로 큰 손실을 가져와 2011년부터 작년까지 살 처분된 가금류는 모두 7,146만 마리에 달한다. 다행히 국내에서 발견된 조류인플루엔자 바이러스가 인체로 전파된 경우는 보고된 바 없으나 국외의 A/H5N1, A/H5N6, A/H7N9은 인체 감염의 잠재적 요인으로 여겨진다.표 3. 조류인플루엔자 국내 발생 현황(2003 ~ 2017)
항목 A/H5N1 A/H5N8 A/H5N6 A/H7N9
발생 시기 2003, 2006, 2008, 2010 2014~2015 2016~2017 2016 최근 발생시기 2011.5 2017.11 2017.11 2016 위험 요인 가금류 농가, 생 가금류 시장 가금류 농가, 야생 철새 가금류 농가, 야생 철새 야생 철새 인체감염 발생 사례 없음 없음 없음 없음
2.2.2 국외 발생 현황
국외에서 발생한 고병원성 조류인플루엔자 바이러스는 A/H5N1, A/H7N9, A/H5N6이다(표 4). A/H5N1은 2003년 이후 전 세계적으로 꾸준히 발생하고 있다. 국내의 경우와 달리 전 세계 16개국에서 인체감염 사례가 보고되었다(그림 4). 이집트와 인도네시아에서는 풍토병 양상으로 발생하였으나 대부분의 나라에서는 산발적으로 발생하고 있다. 환자 860명 중 454명이 사망함으로써 52.8%의 높은 사망률을 보인다[5]. 처음 A/H5N1이 발생 하였을 때 잠재적 대유행으로 이어질 것으로 우려되었으나 2014년 이후 인체 감염사례가 줄어들고 2017년에는 5건 이하의 인체감염 사례가 보고되었다. A/H7N9은 2013년 3월 이후 중국에서 발견된 이래 현재까지 산발적으로 일어나고 있지만 2013년 이후 지속적으로 일어나고 있다. 또한 홍콩, 대만, 말레이시아, 캐나다에서도 인체 감염이 보고되면서 잠재적 대유행의 위협이 되는 바이러스이다. 앞서 언급했듯이 첫 발견은 저병원성 조류인플루엔자 바이러스였으나 지속적인 변이로 고병원성 바이러스로 발견되었다. 전형적인 계절성 유행성 독감과 유사한 증상이 발생하고 3~7일 이내 급성호흡부전,
고병원성 조류인플루엔자 바이러스의 생태 및 국내외 발생 현황 신유진 Page 7 / 10 다발성장기부전, 중증 폐렴, 패혈증으로 진행되어 사망에 이르는 것으로 보고된다[18]. A/H5N6는 중국에서 가장 먼저 발견된 후 국내에서도 발견되었고 산발적으로 발생하고 있는 조류인플루엔자 바이러스이다. 2014년에서 2016년 사이 18명의 확진 환자 중 10명이 사망함에 따라 55.6%의 높은 사망률을 보인다. 표 4. 조류인플루엔자 국외 발생 현황(2003 ~ 2017)
항목 A/H5N1 A/H7N9 A/H5N6
발생국 16개국(그림 4) 중국, 홍콩, 대만, 말레이시 아, 캐나다 중국, 베트남, 라오스 최근 발생국 인도네시아 중국, 홍콩 중국 인체감염 발생 사례 2003~2017. 11 확진860명 사망 454 명 (52.8%) 2013~2017.9 확진 1,622명 사망 619명 (38.2%) 2014~2017.11 확진 18명 사망 10명 (55.6%) 사람 간 전파사례 가족, 병원 내 제한적 전파 가족, 병원 내 제한적 전파 없음 발생 양상 이집트, 인도네시아 풍토병 양상 그 외 산발적 발생 계절적 유행 (10~4월) 산발적 발생 위험 요인 가금류 농가, 생 가금류 시장 생 가금류 시장 가금류 농가, 생 가 금류 시장 그림 4. 전 세계 A/H5N1의 인체 감염 사례
고병원성 조류인플루엔자 바이러스의 생태 및 국내외 발생 현황 신유진 Page 8 / 10
3. 결론
고병원성 조류인플루엔자는 5번째 잠재적인 인플루엔자 대유행 바이러스로 예측되며 지속적인 위협이 되고 있다. 조류에서 고병원성인 A/H5N1은 인체 감염의 경우에도 50% 이상의 높은 사망률을 보이며 대유행으로 이어질 경우 치명적인 바이러스이다. 하지만 2016년 이집트, 2017년 인도네시아에서 최근 2년간 15건 이내의 감염 사례가 보고되면서 점차 발생의 빈도수가 줄어들고 있다. 한편, A/H7N9은 중국에서 2013년 첫 발견 이후 매년 증가하는 양상을 보이며 2017년까지 총 1,500명이 넘는 감염사례가 보고되면서 잠재적 대유행으로 이어질 것으로 우려되고 있다. A/H5N1보다 사망률은 낮지만 첫 발견 시 저병원성이 있던 A/H7N9는 최근 발생한 경우는 고병원성으로 드러나 지속적인 변이가 일어남으로써 주의가 필요하다. 중국의 경우 A/H7N9의 인체 감염은 2013년 이후 산발적이지만 지속적으로 일어나고 있어 중국 정부는 예방, 감시, 대유행을 대비한 지속적인 모니터링과 함께 A/H5와 A/H7의 bivalent 백신을 가금류에 접종하고 있다[19]. 다행히 국내에서 조류인플루엔자 바이러스 인체 감염사례는 보고된 바가 없지만 A/H5N1과 A/H7N9의 인체 감염이 우리와 지리적으로 근접한 중국에서 지속적으로 발생하고 있어 각별한 주의가 요구된다. 중국과 우리나라는 왕래가 빈번한 상황으로 무역, 관광 등 다양한 경로로 바이러스의 전파가 가능하다. 질병관리본부는 조류인플루엔자 감염 예방 및 관리지침을 개정하고 환자의 조기 발견을 위해 노력하고 있으며 가금류 농가, 철새 도래지 방문 자제, 개인위생 수칙 준수 당부하고 있고 야생 조류 및 사체 접촉 후 10일 이내 발열을 동반한 호흡기 증상이 발생하면 관할 지역 보건소 질병관리 본부에 신고를 권고한다. 마지막으로 A/H5N1, A/H7N9뿐 아니라 국내에서 처음 발생하여 미국, 유럽 등에서도 발견된 A/H5N8은 조류에서 고병원성의 바이러스이다[20;21]. 현재까지 인체 감염 사례는 알려지지 않았으나 지속적인 관찰이 필요하다.4. 참고문헌
[1] Hause BM, Ducatez M, Collin EA, Ran Z, Liu R, Sheng Z, Armien A, Kaplan B, Chakravarty S, Hoppe AD, Webby RJ, Simonson RR, Li F. Isolation of a novel swine influenza virus from Oklahoma in 2011 which is distantly related to human influenza C viruses. PLoS Pathog. 2013. 9(2):e1003176.
[2] Centers for Disease Control and Prevention, CDC. Transmission of Influenza Viruses from Animals to People; https://www.cdc.gov/flu/about/viruses/transmission.htm. Accessed Oct. 28, 2017.
[3] CDC. Past Pandemics;https://www.cdc.gov/flu/pandemic-resources/basics/past-pandemics.html. Accessed Oct. 29, 2017.
[4] 질병관리본부 (KCDC), 신종인플루엔자 A(H1N1) 유행경과; http://www.cdc.go.kr/CDC/contents/ CdcKrContentView.jsp?cid=26739&menuIds=HOME001-MNU1132-MNU1013-MNU1506-MNU1508. Accessed Oct. 29, 2017.
[5] Center for Health Protection. Avian Influenza; Latest Avian Influenza Updates – Global statistics and affected areas of avian influenza; http://www.chp.gov.hk/files/pdf/global_statistics_avian_influenza_e.pdf. Accessed Nov. 29, 2017.
고병원성 조류인플루엔자 바이러스의 생태 및 국내외 발생 현황 신유진 Page 9 / 10
mobile/26/68926_view.html. Accessed Nov. 29, 2017.
[7] Cheung, CY., Poon, LL., Lau, AS., et al. (2002). Induction of proinflammatory cytokines in human macrophages by influenza A (H5N1) viruses: a mechanism for the unusual severity of human disease? Lancet 360, 1831–7.
[8] Gao R, Cao B, Hu Y, Feng Z, Wang D, Hu W, Chen J, Jie Z, Qiu H, Xu K, Xu X, Lu H, Zhu W, Gao Z, Xiang N, Shen Y, He Z, Gu Y, Zhang Z, Yang Y, Zhao X, Zhou L, Li X, Zou S, Zhang Y, Li X, Yang L, Guo J, Dong J, Li Q, Dong L, Zhu Y, Bai T, Wang S, Hao P, Yang W, Zhang Y, Han J, Yu H, Li D, Gao GF, Wu G, Wang Y, Yuan Z, Shu Y. Human infection with a novel avian-origin influenza A (H7N9) virus. N Engl J Med. 2013 May 16;368(20):1888-97.
[9] 생명공학정책연구센터. 조류인플루엔자 발생 현황; http://www.bioin.or.kr/InnoDS/data/upload/issue/ b3b65b3c26984d06a4deb474c71e3b62.pdf. Accessed Nov. 28, 2017.
[10] Matrosovich, M., Zhou, N., Kawaoka, Y., and Webster, R. (1999). The surface glycoproteins of H5 influenza viruses isolated from humans, chickens, and wild aquatic birds have distinguishable properties. Journal of virology, 73(2):1146{55.
[11] Herfst S, Schrauwen EJ, Linster M, Chutinimitkul S, de Wit E, Munster VJ, Sorrell EM, Bestebroer TM, Burke DF, Smith DJ, Rimmelzwaan GF, Osterhaus AD, Fouchier RA. Airborne transmission of influenza A/H5N1 virus between ferrets. Science. 2012. 336(6088):1534-41.
[12] Belser JA, Gustin KM, Pearce MB, Maines TR, Zeng H, Pappas C, Sun X, Carney PJ, Villanueva JM, Stevens J, Katz JM, Tumpey TM. Pathogenesis and transmission of avian influenza A (H7N9) virus in ferrets and mice. Nature. 2013. 26;501(7468):556-9.
[13] Abente EJ, Kitikoon P, Lager KM, Gauger PC, Anderson TK, Vincent AL. A highly pathogenic avian-derived influenza virus H5N1 with 2009 pandemic H1N1 internal genes demonstrates increased replication and transmission in pigs. J Gen Virol. 2017. 98(1):18-30.
[14] Qi W, Jia W, Liu D, Li J, Bi Y, Xie S, Li B, Hu T, Du Y, Xing L, Zhang J, Zhang F, Wei X, Eden JS, Li H, Tian H, Li W, Su G, Lao G, Xu C, Xu B, Liu W, Zhang G, Ren T, Holmes EC, Cui J, Shi W, Gao GF, Liao M. Emergence and adaptation of a novel highly pathogenic H7N9 influenza virus in birds and humans from a 2013-human-infecting low pathogenic ancestor. J Virol. 2017. pii: JVI.00921-17.
[15] Liu D, Shi W, Shi Y, Wang D, Xiao H, Li W, Bi Y, Wu Y, Li X, Yan J, Liu W, Zhao G, Yang W, Wang Y, Ma J, Shu Y, Lei F, Gao GF. Origin and diversity of novel avian influenza A H7N9 viruses causing human infection: phylogenetic, structural, and coalescent analyses. Lancet. 2013. 381(9881):1926-32.
[16] Liu D1, Shi W2, Gao GF. Poultry carrying H9N2 act as incubators for novel human avian influenza viruses. Lancet. 2014. 8;383(9920):869.
[17] Kim HK, Jeong DG, Yoon SW. Recent outbreaks of highly pathogenic avian influenza viruses in South Korea. Clin Exp Vaccine Res. 2017. 6(2):95-103.
[18] 질병관리본부 (KCDC). ’16-‘17절기 중국 조류인플루엔자(A/H7N9) 인체감염 발생 동향;
http://cdc.go.kr/CDC/mobile/info/CdcKrInfo0301.jsp?menuIds=HOME001-MNU1154-MNU0005-MNU0037&cid=75127. Accessed Dec. 05, 2017.
[19] WHO. Influenza at the human-animal interface_Summary and assessment, 25 July 2017 to 27 September 2017;http://www.who.int/influenza/human_animal_interface/Influenza_Summary_IRA_HA_interface_09_27_2017.p df?ua=1. Accessed Oct. 28, 2017.
Shearn-고병원성 조류인플루엔자 바이러스의 생태 및 국내외 발생 현황 신유진 Page 10 / 10
Bochsler V, Killian ML, Pedersen JC, Hines N, Gidlewski T, DeLiberto T, Sleeman JM. Novel Eurasian highly pathogenic avian influenza A H5 viruses in wild birds, Washington, USA, 2014. Emerg Infect Dis. 2015. 21(5):886-90.
[21] European Food Safety Authority. Highly pathogenic avian influenza A subtype H5N8; http://onlinelibrary.wiley.com/doi/10.2903/j.efsa.2014.3941/epdf. Accessed Dec. 05, 2017.
※ 본 콘텐츠는 의 후원으로 작성되었습니다.
The views and opinions expressed by its writers do not necessarily reflect those of the Biological Research Information Center. 신유진(2018). 고병원성 조류인플루엔자 바이러스의 생태 및 국내외 발생 현황. BRIC View 2018-T04
Available from http://www.ibric.org/myboard/read.php?Board=report&id=2899 (Feb 06, 2018) Email: member@ibric.org

![그림 2. 인플루엔자 A 바이러스 복제 사이클 [출처: antiflu project]](https://thumb-ap.123doks.com/thumbv2/123dokinfo/5123563.86642/4.892.61.845.101.373/그림-인플루엔자-바이러스-복제-사이클-출처-antiflu-project.webp)
![그림 3. 다수의 유전자 재편성을 거듭한 A/H7N9 바이러스의 구조[출처: CDC: Transmission of influenza virues from animals to people]](https://thumb-ap.123doks.com/thumbv2/123dokinfo/5123563.86642/5.892.95.807.612.960/다수의-유전자-재편성을-거듭한-바이러스의-transmission-influenza-animals.webp)