INTRODUCTION
Nelumbo nucifera Gaertn. known as sacred lotus is a peren-nial aquatic crop grown and consumed over the world. Al-most all parts of the N. nucifera Gaertn. plant are eaten as a vegetable and used in indigenous systems of medicine in Eastern Asia, particularly in China, Japan, Korea and possi-bly India (Ling et al. 2005). Interest in N. nucifera Gaertn. as harmless medicine has increased because of its broad spectrum of biological properties. Previous research indi-cates that various parts of N. nucifera Gaertn. possess anti-diarrheal, psychopharmacological, diuretic, antipyretic (Su-jay et al. 2006) and antioxidant activities (Hu and Skibsted 2002; Jung and Kim 2003; Wu and Wang 2003).
Radiation is an important environmental risk factor for various cancers and also a major therapeutic agent in
can-cer treatment. Exposure of mammalian cells to irradiation induces several types of damage. Extensive progress has been made in understanding the mechanisms by which irra-diation rapidly produces a variety of changes that result in damage to critical cellular targets like DNA, lipids, proteins, and membranes (Cadet et al. 2004; Chung and Broaddus 2005). It has been shown to enhance the production of harm-ful reactive oxygen species (ROS) in a variety of cells, (Repine et al. 1981; Yamaguchi et al. 1994). Lipids are the most common biological targets of ROS (William and Ned 1990). Lipid oxidation gives rise to a number of secondary products. Lipid peroxidation-derived aldehydes (LPDAs) have the ability to exacerbate oxidative damage (Uchida 2000). Longevity and high reactivity allow these molecules to act inside and outside of cells, where they interact with biomolecules, such as nucleic acids and proteins (Rio et al. 2005). Because of this, there is an increasing interest in nat-ural radioprotector, namely flavonoids, present in medicinal and dietary plants, that may help prevent oxidative damages
─ ─ 259 ──
Protective Effects of Nelumbo nucifera against
γ
-
Irradiation
-induced Lipid Peroxidation in Mice Urine
Il Yun Jeong*, Hyo Jung Lee1, Yong Dae Park, Chang Hyun Jin and Dae Seong Choi Advanced Radiation Technology Institute, Korea Atomic Energy Research Institute,
Jeongeup 580-185, Korea
1Department of Life Science, Gwangju Institute of Science and Technology, Gwangju 500-712, Korea
Abstract -- The radioprotective effect of isoquercitrin-abundant fraction (IAF) of N. nucifera
Gaertn. leaf extract against γγ-irradiation-induced oxidative stress was evaluated by the lipid peroxi-dation-derived aldehydes (LPDAs) as a marker for oxidative risk in mice urine, and the DNA damage using comet assay in RAW 264.7 cells. Mice that were treated with IAF (50 mg/kg) and γγ
-irradiation showed considerably decreased LPDA levels relative to those that had received γγ- irra-diation alone. Furthermore, pretreatment with IAF resulted in a significant decrease in the amount of DNA damage in cells. It is demonstrated that pretreatment with IAF of N. nucifera Gaertn. gives protection against irradiation-induced cellular damage.
Key words : Nelumbo nucifera, Ionizing irradiation (IR), Reactive oxygen species (ROS), Isoquer-citrin
* Corresponding authors: Il Yun Jeong, Tel. +82-63-570-3150, Fax. +82-63-570-3159, E-mail. [email protected]
(Benkovic et al. 2008). In this study, the effects of the radio-protective effect of isoquercitrin-abundant fraction (IAF) of N. nucifera Gaertn. leaf extract against γ-irradiation-induced oxidative stress were examined. The efficiency of IAF was evaluated using the LPDAs as a marker for oxidative risk in mice urine, and the DNA damage using comet assay in RAW 264.7 cells.
MATERIAL AND METHODS
1. General experimental procedures
1H (500 MHz), 13C (125 MHz) and two-dimensional (2D)
NMR spectra were recorded with a JNM-ECA 500 spectro-meter (Jeol, Japan) using CD3OD, DMSO-d6, as a solvent.
Analytical and quantitative HPLC were performed using an Agilent Technologies Series 1100 system equipped with an automatic injector, a column oven and a UV detector. An RP-18 (150 mm×4.6 mm i.d., 3μm particle size,
Phenome-nex) was also used. The Diaion HP-20 resin used in this experiment was purchased from Mitsubishi (Tokyo, Japan).
2. Isolation and quantitative analysis of the isoquercitrin abundant fraction
N. nucifera leaves (1.3 kg) collected at Jeong-up, Korea, 2006, were extracted with 70% ethanol for 6 h, three times at room temperature. After filtration, the 70% ethanol extract was evaporated. The concentrated leaf ethanolic extract (124 g) was subjected to column chromatography with Dia-ion HP-20 resin, sequentially using 20% aqueous MeOH and MeOH as eluents. The isoquercitrin abundant fraction (IAF) was obtained from a MeOH eluate (Fraction 2, 31.1 g). The IAF was separated by column chromatography on a silica gel (230~400 mesh, Merch, Ø5×30 cm) using a
gra-dient of CHCl3-MeOH (90 : 10 to 0 : 100) to give seven frac-tions (Fraction 2-1~7). Fractions 2-3 (9.45 g) were subjected to silica gel column chromatography with CHCl3-MeOH to
yield nine fractions. Fractions 2-3~7 (317 mg) were further purified by semipreparative HPLC with H2O and MeOH
(3 : 7) to yield isoquercitrin (5 mg). Quantitative analysis was performed using an Agilent Technologies Series 1100 system. The column temperature was maintained at 23�C, with an injection volume of 10μl and a flow rate of 1 ml min-1. The IAF extract was separated using a reverse-phase
C18 column with a 0.2% acetic acid-acetonitrile mobile phase
and detected at 340 nm. An IAF was prepared in methanol at 10 mg ml-1. Isoquercitrin standard solution was prepared
by dilution of the stock solution with methanol to give solu-tions in the range of 12~500μg ml-1. The extract was
sub-jected to quantitative analysis, and the concentrations of the markers were determined by an external standard method.
3. Cell culture and animals
The female BALB/c mice were housed in polycarbonate cages and fed a standard animal diet and water. Animals were kept at 22~24�C with a 12 h light/dark cycle. Research was conducted according to the principles described in the ‘Animal Care Act’ prepared by the Ministry of Agriculture and Forestry, Republic of Korea. The BALB/c mice were purchased from Orient Inc. (Charles River Technology, Seoul, Korea). RAW 264.7 cells were routinely cultured in DMEM medium (Invitrogen, CA) supplemented with 10% fetal bovine serum, 100 U ml-1penicillin and 100 U ml-1
streptomycin at 37�C in a humidified atmosphere of 5% CO2.
Mice and cells were irradiated at room temperature with 137Cs γ-rays at a dose rate of 0.84 Gy min-1from a
biologi-cal irradiator, Gamma Cell-40 Exactor (MDS Nordion, Canada).
4. Urine collection and hydrazone derivatives Mice were randomly divided into three main groups. In the control group (n==8), the animals received a daily oral
injection of 0.9% NaCl/D.W. (v/v, 20/1) for three days. In the gamma-irradiated group (n==8), the animals were exposed to a single dose of 4 Gy whole-body irradiation. In the IAF and gamma-irradiated group (n==8), the animals were injected daily with freshly prepared IAF (0.5 ml, 50 mg kg-1body
weight) for three days before irradiation exposure. Animals were injected daily at 4:00 p.m. and placed in a specially designed well-ventilated acrylic container in which their entire bodies were exposed to 4 Gy. Animals were placed in metabolism cages to collect urine for 12 h urine collection following irradiation. During urine collection, the animals were allowed free access to tap water but received no food. The urine-collecting vessels were positioned over Styrofoam containers filled with dry ice, which permitted the collection of urine in the frozen state over 4-hour periods.
Phenylhydrazine (PH) was used as the derivatizing agent to identify urinary metabolites. Phenylhydrazine (22μl) was
dissolved in 5 ml of 2 M hydrochloric acid in order to make the derivatizing reagent. In 50 ml screw-capped PTFE-lined tubes, 2.0 ml aliquots of urine, 7.6 ml of water and 0.4 ml of PH reagent were mixed, followed by the addition of 10 ml of n-hexane. The tubes were then intermittently shaken for 10 min and centrifuged at 3000 rpm for 10 min. The sepa-rated organic phase was concentsepa-rated and dissolved in chlo-roform for GC-MS analysis. In order to make a standard solution, phenyhydrazine (16.5 mM) and aldedhydes in ethanol were added to acetic acid (1 mM) at room tempera-ture, and the mixture was concentrated in vacuo.
5. Gas chromatography-mass spectrometry GC-MS analysis was carried out on a 6890 gas chroma-tograph (Agilent) coupled to a 5975 mass spectrometer equipped with an HP-5 fused silica capillary column (25 m, 0.32 mm i.d., 0.25μm film thickness). GC conditions includ-ed helium flow at a rate of 0.7 ml min-1, and the injection
was carried out in the splitless mode. The temperature pro-gram consisted of a starting temperature of 70�C, which was increased to 175�C at a rate of 25�C/min, then to 200�C at a rate of 5�C/min, and finally to 300�C at a rate of 10�C min-1. The instrument was set on electron ionization mode.
The ion source temperature was 180�C and the ionization energy was 70 eV.
6. Comet assay
Single-cell gel electrophoresis (comet assay) was per-formed under alkaline conditions. The cell suspension (50 μl) was mixed with 80 μl of 0.5% low melting point agarose. Mixtures were spread onto microscope slides pre-coated with 0.6% normal melting point agarose and another 110μl of 0.5% low melting point agarose was used to cover the surface. The slides were kept in a 4�C refrigerator for 10 min to allow the agarose to solidify. The slides were then immersed in chilled lysis buffer (containing 2.5 M NaCl, 0.1 M EDTA, 10 mM Tris-base, pH 10, 10% DMSO, and 0.4% Triton X-100) at 4�C for 1 h. Slides were transferred into a horizontal electrophoresis tank filled with chilled electro-phoresis buffer (0.3 M NaOH, 1 mM EDTA) for 20 min to allow for DNA unwinding. Electrophoresis was performed at 25 V and 300 mA at 4�C for 20 min. After electrophoresis, the slides were neutralized in neutralization buffer, stained with ethidium bromide, kept in a humidified air-tight
con-tainer and examined using a fluorescence microscope. One hundred comets were observed for each sample. The inten-sity of the comet tail DNA relative to that of the head DNA, reflecting the number of DNA breaks, was determined with comet assay software.
7. Data analysis
All values are expressed as mean±standard deviation (SD). The statistical significance of differences between groups was evaluated for two parallel experiments using the student’s t-test.
RESULTS
Dried N. nucifera Gaertn leaves were finely sliced and extracted with 70% EtOH. This extract was submitted to chromatography on a Diaion HP-20 column to yield two eluted fractions, 20% aqueous MeOH and MeOH. Fraction 2 (MeOH eluate, IAF) was found to be the most suitable fraction for reproducible and effective partitioning of the active compounds. Fraction 2 was further separated by appli-cation to successive silica gel, sephadex LH-20 and semipre-parative HPLC, leading to isolation of the phenolic com-pound. Based on the 1H NMR, 13C NMR and MS spectra,
the isolated compound was easily identified as isoquercitrin by comparison of its spectral data with that reported previ-ously (Ohkoshi and Miyazaki 2007). The next step in our work was focused on determining the experimental condi-tions necessary to investigate the isoquercitrin content of IAF from N. nucifera Gaertn leaf extract by HPLC. The chromatograms of IAF and the isoquercitrin are shown in Fig. 1. An acetic acid (0.2% aqueous solution) and acetoni-trite (CH3CN) gradient was found to be an effective mobile
phase for HPLC separation. As can be seen, the peaks in the IAF chromatogram were identified as the isoquercitrin by comparison of HPLC retention times. The peaks correspon-ding to isoquercitrin eluted from the column in the same order and with the same retention time (19.8 min) as the authentic standard. For isolated isoquercitrin, the calibration curve was linear with R2==0.9996. The isoquercitrin
concen-tration was calculated to be about 39 mg g-1of IAF.
Analysis of the LPDAs present in urine was performed by conversion to the corresponding hydrazones using phenyl-hydrazine. Fig. 2 shows chromatograms of hydrazone
deri-vatives in mouse urine. γ-Irradiation (4 Gy) induced a signi-ficant increase in the levels of lipid peroxidation products in mouse urine compared to the control group. Pre-treatment with IAF ameliorated the effects of IR, and the concentration of lipid peroxidation products was significantly decreased compared to that in the irradiated group. Based on the above observations, we utilized GC co-elution methods as well as
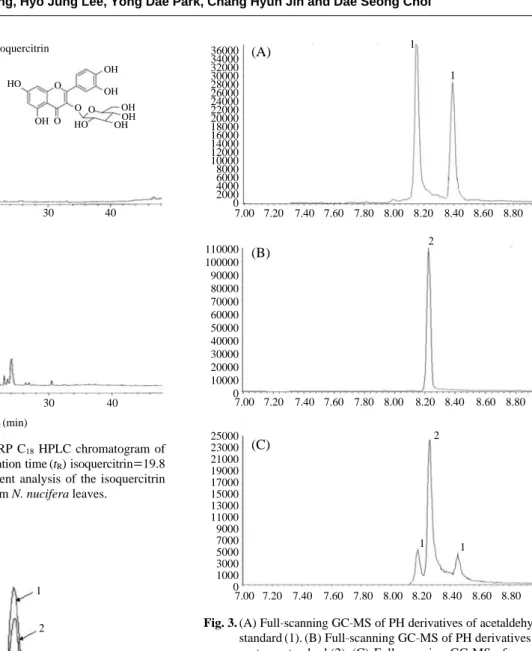
MS techniques to identify lipid degradation products. As shown in Fig. 3, the acetone derivative eluted at 8.26 min, while the acetaldehyde derivatives showed two peaks (8.18 and 8.44 min), which correspond to the E and Z isomers that were separated by GC-MS. The molecular ions 134 and 148 of the synthetic hydrazones were also identified and found to originate from acetaldehyde and acetone, respecti-vely.
We also examined the effects of γ-irradiation on DNA damage using comet assay. Pretreatment of RAW 264.7 cells with IAF in DMSO decreased DNA damage in irradi-ated RAW 264.7 cells compared with untreirradi-ated cells (Fig. 4). Fig. 1. HPLC chromatogram (A) RP C18HPLC chromatogram of
isolated isoquercitrin: Retention time (tR) isoquercitrin==19.8
min. (B) Isoquercitrin content analysis of the isoquercitrin abundant fraction (IAF) from N. nucifera leaves.
Fig. 2. Chromatogram of a mouse urine sample spiked with differ-ent amounts of four lipid degradation products. (1) 4 Gy
-irradiated group; (2) Non-irradiation group; (3) N. nucifera treated and irradiated group.
Fig. 3. (A) Full-scanning GC-MS of PH derivatives of acetaldehyde standard (1). (B) Full-scanning GC-MS of PH derivatives of acetone standard (2). (C) Full-scanning GC-MS of mouse urine (3). mAU 17.5 15 12.5 10 7.5 5 2.5 0 mAU 500 400 300 200 100 0 Isoquercitrin HO HO OH OH OH OHOH OH O O O O 0 10 20 30 40 0 10 20 30 40 Time (min) (A) (B) 300000 280000 260000 240000 220000 200000 180000 160000 140000 120000 100000 80000 60000 40000 20000 0 7.90 7.95 8.00 8.05 8.10 8.15 8.20 8.25 8.30 8.35 8.40 8.45 8.50 8.55 8.60 1 2 3 36000 34000 32000 30000 28000 26000 24000 22000 20000 18000 16000 14000 12000 10000 8000 6000 4000 2000 0 110000 100000 90000 80000 70000 60000 50000 40000 30000 20000 10000 0 25000 23000 21000 19000 17000 15000 13000 11000 9000 7000 5000 3000 1000 0 7.00 7.20 7.40 7.60 7.80 8.00 8.20 8.40 8.60 8.80 7.00 7.20 7.40 7.60 7.80 8.00 8.20 8.40 8.60 8.80 7.00 7.20 7.40 7.60 7.80 8.00 8.20 8.40 8.60 8.80 1 1 2 2 1 1 (A) (B) (C)
DISCUSSION
Evidence for the involvement of reactive oxygen species (ROS) in ionizing irradiation comes from previous works showing persistent oxidative stress in cells (Redpath and Gutierrez 2001; Rugo et al. 2002). Similar observations are reported on irradiation-induced oxidative damage in several organs (Sener et al. 2003; Bhatia and Manda 2004). Ionizing irradiation generates ROS as a result of water radiolysis. If not eliminated, these ROS can induce oxidative damage to vital cellular molecules and structures including DNA, lipids, proteins, and membranes (Cadet et al. 2004; Chung and Broaddus 2005). Because of this, there is an increasing interest in natural radioprotector, namely flavonoids, present in medicinal and dietary plants, that may help prevent oxida-tive damages (Benkovic et al. 2008). Flavonoids are the most common and widely distributed group of plant pheno-lics, and they act as potent metal chelators and free-radical scavengers. Our results suggest that the major flavonoid in N. nucifera Gaertn. is isoquercitrin, and it is abundant in
isoquercitrin-abundant fraction (IAF) prepared by using column chromatography with Diaion HP-20 resin. Several previous studies have been reported that isoquercitrin has strong radical scavenging activities (Gadow et al. 1997).
The major forms of cellular damage induced by irradiation are lipid peroxidation, and DNA damage. The present study demonstrates that concentration of LPDAs in mice urine indicating the level of oxidative stress was markedly enhan-ced with irradiation. However, pretreatment of IAF to mice resulted in decreased LPDAs (Fig. 2). A study of the mech-anisms of the adverse effects of lipid peroxidation in biolo-gical systems led to the hypothesis that toxic products origi-nate from lipid peroxidation of cellular membranes, which then diffuse and act at a distance. Some of these toxic pro-ducts have been isolated and characterized (Sim et al. 2003). Of these, 4-hydroxy-2-nonenal (HNE), malondialdehyde (MDA), acrolein, and crotonaldehyde have been intensely studied with respect to their chemical and biological intera-ctions with proteins and nucleic acid bases (Cighettiet et al., 1999). Our data revealed that urinary level of acetaldehyde and acetone was observed within 12 h after γ-irradiation. Fig. 4. Protective effects of N. nucifera on 137Cs γ-rays irradiation-induced DNA damage in RAW 264.7 macrophage cells. (A) Non-
irradi-ated cells, 4 Gy-irradiated cells, and N. nucifera treated and irradiated cells (B) Quantification of comet tails (p⁄0.05 compared to the control value; *p⁄0.05 compared to the value of 4 Gy irradiation alone).
Control 4 Gy 4 Gy++IAF Tail DNA (%) 50 40 30 20 10 0 Control 0 100μg/ml 4 Gy (A) (B)
Shara et al. have reported that malondialdehyde, acetalde-hyde and acetone were simultaneously identified in rat urine following chemically induced oxidative stress (Shara et al. 1992). These results indicate that LPADs such as acetalde-hyde and acetone are toxic products induced by oxidative stress.
Several techniques have been used to isolate and detect the small amounts of aldehydes that are formed in vivo. HPLC and gas chromatography (GC) are the most com-monly used techniques for the detection and quantification of aldehyde derivatives. In the field of GC, Cighetti and colleagues developed and fully validated a method based on derivatization of aldehydes with phenylhydrazine (PH) fol-lowed by mass spectrometric (MS) detection. This assay is characterized by mild reaction conditions to form a GC analyzable adduct (Esterbauer et al. 1991).
Moreover, our results also showed that γ-irradiation sig-nificantly affected the levels of primary DNA damage in RAW 264.7 cells, and that pretreatment with IAF decreased DNA damage in irradiated RAW 264.7 cells compared with untreated cells. An increase in DNA damage after irradiation has been observed in several different studies (Hosseinimehr et al. 2003; Mansour et al. 2008). In addition, DNA damage in cells exposed to γ-irradiation is an important marker of oxidative stress (Lee et al. 2007). IAF was found to protect against DNA damage induced by irradiation.
In conclusion, IAF from N. nucifera Gaertn. was shown to have radioprotective effect against γ-irradiation by pre-venting oxidative stress. This might be through antioxidant activities of isoquercitrin. Thus, supplementation with IAF of N. nucifera Gaertn. leaves may have a benefit for the protection against irradiation in medical and industrial fields.
ACKNOWLEDGMENT
This research was supported by Agricultural R&D Pro-motion Center of the Ministry for Food, Agriculture, Forestry and Fisheries, Korea.
REFERENCES
Benkovic V, Knezevic AH, Dikic D, Lisicic D, Orsoic N, Basic I, Kosalec I and Kopjar N. 2008. Radioprotective effects of
propolis and quercetin in γ-irradiated mice evaluated by alkaline comet assay. Phytomedicine. 15:851-858. Bhatia AL and Manda K. 2004. Study on pre-treatment of
melatonin against irradiation-induced oxidative stress in mice. Environ. Toxicol. Pharmacol. 18:13-20.
Cadet J, Bellon S, Douki T, Frelon S, Gasparutto D, Muller E, Pouget JP, Ravanat JL, Romieu A and Sauvaigo S. 2004. Irradiation-induced DNA damage: formation, measurement, and biochemical features. J. Environ. Pathol. Toxicol.
Oncol. 23:33-43.
Chung D and Broaddus C. 2005. Molecular targeting in radi-otherapy: epidermal growth factor receptor. Mol. Interv. 5:15-19.
Cighetti G, Debiasi S, Paroni R and Allevi P. 1999. Free and total malondialdehyde assessment in biological matrices by gas chromatography-mass spectrometry: what is needed for an accurate detection. Anal. Biochem. 266:222-229. Esterbauer H, Schaur J and Zollner H. 1991. Chemistry and
biochemistry of 4-hydroxynonenal, malonaldehyde and related aldehydes. Free Radic. Biol. Med. 11:81-128. Gadow AV, Joubert E and Hansmann CF. 1997. Comparison
of the antioxidant activity of aspalathin with that of other plant phenols of rooibos tea (Aspalathus linearis), α- Toco-pherol, BHT, and BHA. J. Agric. Food Chem. 45:632-638. Hosseinimehr J, Tavakoli H, Pourheidari R, Sobhani A and
Shafiee A. 2003. Radioprotective effects of citrus extract against gamma irirradiation in mouse bone marrow cells. J.
Radiat. Res. 44:237-241.
Hu M and Skibsted H. 2002. Antioxidative capacity of rhizome extract and rhizome knot extract of edible lotus (Nelumbo
nucifera). Food Chem. 76:327-333.
Jung A and Kim E. 2003 Antioxidant principles of Nelumbo
nucifera stamens. Arch. Pharm. Res. 26:279-285.
Lee JH, Kim SY, Kil IS and Park JW. 2007. Regulation of ionizing irradiation-induced apoptosis by mitochondrial NADP++
-dependent isocitrate dehydrogenase. J. Biol. Chem. 282:13385-13394.
Ling ZQ, Xie BJ and Yang EL. 2005. Isolation, Characteriza-tion, and determination of antioxidative activity of oligo-meric procyanidins from the seedpod of Nelumbo nucifera Gaertn. J. Agric. Food Chem. 53:2441-2445.
Mansour H, Hafez F and Fahmy M. 2008. Protective effect of
N-actylcysteine against irradiation induced DNA damage and hepatic toxicity in rats. Biochem. Pharmacol. 75:773 -780.
Ohkoshi E and Miyazaki H. 2007. Constituents from the leaves of Nelumbo nucifera Stimulate Lipolysis in the White Adi-pose Tissue of Mice. Planta Med. 73:1255-1259.
Repine E, Pfenninge W, Talmage W, Berger M and Pettijohn E. 1981. Dimethyl sulfoxide prevents DNA nicking
medi-ated by ionizing irradiation or iron/hydrogen peroxide- gen-erated hydroxyl radical. Proc. Natl. Acad. Sci. USA. 78: 1001-1003.
Redpath JL and Gutierrez M. 2001. Kinetics of induction of reactive oxygen species during the post-irradiation expres-sion of neoplastic transformation in vitro. Int. J. Radiat.
Biol. 77:1081-1085.
Rio D, Amanda S and Nicoletta P. 2005. A review of recent studies on malondialdehyde as toxic molecule and biologi-cal marker of oxidative stress. Nutr. Metab. Cardiovas. 15: 316-328.
Rugo RE, Secretan MB and Schiestl RH. 2002. X irradiation causes a persistent induction of reactive oxygen species and a delayed reintroduction of TP53 in normal human diploid fibroblasts. Radiat. Res. 158:210-219.
Sener G, Jahovic N, Tosun O, Atasoy BM and Yegen B. 2003. Melatonin meliorates ionizing irradiation-induced oxidative organ damage in rats. Life Sci. 74:563-572.
Shara MA, Dickson PH, Bagchi D and Stohs SJ. 1992. Excre-tion of formaldehyde, malondialdehyde, acetaldehyde and acetone in the urine of rats in response to 2,3,7,8- tetrachlo-rodibenzo-p-dioxin, paraquat, endrin and carbon tetrachlo-ride. J. Chromatogr. 576:221-233.
Sim S, Salonikas C, Naidoo D and Wilcken E. 2003. Improved
method for plasma malondialdehyde measurement by high -performance liquid chromatography using methyl malondi-aldehyde as an internal standard. J. Chromatogr. B Analyt.
Technol. Biomed. Life Sci. 785:337-344.
Sujay R, Atul W, Kakali M and Bishnu P. 2006. Antioxidant activity of Nelumbo nucifera (sacred lotus) seeds. J.
Ethno-pharmacol. 104:322-327.
Uchida K. 2000. Role of reactive aldehyde in cardiovascular diseases. Free Radic. Biol. Med. 28:1685-1696.
William P and Ned A. 1990. Suggested mechanisms for the production of 4-hydroxy-2-nonenal from the autoxidation of polyunsaturated fatty acids. Free Radic. Biol. Med. 8: 541-543.
Wu J and Wang L. 2003. Antioxidant activity of methanol extract of the lotus leaf (Nelumbo nucifera Gaertn). Am. J.
Chinese Med. 31:687-689.
Yamaguchi S, Sakurada S and Nagumo M. 1994. Role of intra-cellular SOD in protecting leukemic and cancer cells against superoxide and irradiation. Free Radic. Biol. Med. 17:389 -395.
Manuscript Received: October 26, 2009 Revision Accepted: November 10, 2009