Protective effect of TM-25659, known as TAZ
activator on high-fat diet-induced insulin
resistance in C57BL/6J mice
by
Jong Gab Jung
Major in Molecular Medicine
Department of Biomedical Sciences
Protective effect of TM-25659, known as TAZ
activator on high-fat diet-induced insulin
resistance in C57BL/6J mice
by
Jong Gab Jung
A Dissertation Submitted to The Graduate School of
Ajou University in Partial Fulfillment of the Requirements
for the Degree of Ph.D. in Biomedical Sciences
Supervised by
Kwan Woo Lee, M.D., Ph.D.
- Abstract -
Protective effect of TM-25659, known as TAZ activator, on high-fat
diet-induced insulin resistance in C57BL/6J mice
2-butyl-5-methyl-6-(pyridine-3-yl)-3-[2'-(1H-tetrazole-5-yl)-biphenyl-4-ylmethyl]-3H-imidazo[4,5-b]pyridine] (TM-25659), known as the TAZ activator, inhibits adipocyte differentiation by interacting with peroxisome proliferator-activated receptor gamma (PPAR-γ). TM-25659 was previously shown to decrease weight gain in a high fat (HF) diet induced obesity (DIO) mouse model. However, the fundamental mechanisms underlying the effects of TM-25659 remain unknown. Therefore, we investigated the molecular mechanisms underlying the contribution of TM-25659 on palmitate (PA)-induced insulin resistance in both C2 myotubes and HepG2 cells. TM-25659 improved the PA-induced insulin resistance in C2 myotubes. TM-25659 induced expression of FGF21 mRNA and protein and secretion of FGF21 in C2 myotubes via activation of GCN2 pathways (GCN2-phosphor-eIF2α-ATF4 and FGF21). This effect of TM-25659 was diminished by FGF21 siRNA. Furthermore, TM-25659 improved the PA-induced insulin resistance in HepG2 cells. TM-25659 induced activation of AMPK in HepG2 cells. This beneficial effect of TM-25659
homeostasis, insulin sensitivity, insulin resistance, and inflammation, but food intake was not affected. TM-25659 increased expressions of both FGF21 mRNA and protein on HF-diet induced insulin resistance in C57BL/6J mice muscle. Also, TM-25659 induced activation of AMPK protein on HF-diet induced insulin resistance in C57BL/6J mice liver. These data indicate TM-25659 may be beneficial for treating insulin resistance by inducing FGF21 and AMPK activation in models of PA-induced insulin resistance and HF-diet induced insulin resistance.
Key words: TAZ, TM-25659, FGF21, HF, PA, insulin resistance, AMPK
CONTENTS
ABSTRACT ··· ⅰ CONTENTS ··· ⅲ LIST OF FIGURES (PART 1) ··· ⅶ LIST OF FIGURES (PART 2) ··· ⅹ
PART 1. TM-25659-induced activation of FGF21 level decreases insulin resistance and inflammation in skeletal muscle via GCN2 pathways
Ⅰ. INTRODUCTION ··· 2
Ⅱ. MATERIALS AND METHODS ··· 8
A. MATERIALS ··· 9
B. METHODS ··· 10
1. Preparation of PA ··· 10
2. Cell culture ··· 10
3. Immunoblot analysis ··· 11
9. IL-1β assay ··· 15 10. Statistical analysis ··· 15
Ⅲ. RESULTS ··· 16 A. TM-25659 restored PA-induced insulin resistance and inflammation
in skeletal muscle cells ··· 17 B. TM-25659 increased FGF21 levels by GCN2 pathways in skeletal muscle cells ···
··· 22 C. Effects of TM-25659 reduced insulin resistance and inflammation
in skeletal muscle cells treated with or without FGF21 siRNA ··· 27 D. TM-25659 prevented HF-diet induced obesity ··· 31 E. TM-25659 restored the insulin signaling pathways in DIO mice skeletal muscle ···
··· 33 F. TM-25659 lowered the muscle pro-inflammatory cytokine levels in DIO mice ····
··· 35 G. TM-25659 regulates expression of both FGF21 mRNA and protein
in DIO mice skeletal muscles ··· 37
PART 2. Protective effects of TM-25659 on high-fat diet-induced insulin resistance in liver via AMPK pathways
Ⅰ. INTRODUCTION ··· 45
Ⅱ. MATERIALS AND METHODS ··· 52
A. MATERIALS ··· 53
B. METHODS ··· 54
1. Preparation of PA ··· 54
2. Cell culture ··· 54
3. Immunoblot analysis ··· 54
4. RNA isolation and quantitative real-time polymerase chain reaction (PCR) · 55 5. Animal experiments ··· 56
6. Insulin tolerance tests ··· 57
7. Glucose tolerance tests ··· 57
8. Statistical analysis ··· 57
Ⅲ. RESULTS ··· 58 A. TM-25659 restored PA-induced reduction of insulin signaling pathway
D. TM-25659 reduced PA-induced reduction of inflammation in HepG2 cells ··· 67
E. TM-25659 improved glucose homeostasis and insulin sensitivity in DIO mice ··· ··· 69
F. TM-25659 restored the insulin signaling pathways in DIO mice liver ··· 71
G. TM-25659 lowered the serum and liver pro-inflammatory cytokine levels in DIO mice ··· 73
H. TM-25659 activated AMPK pathways in DIO mice liver ··· 77
I. TM-25659 repressed serum lipid levels in HF-diet fed mice ··· 79
J. TM-25659 prevented HF-diet induced hepatic steatosis ··· 81
K. TM-25659 reduced expression of lipogenic genes but increased expression of fatty acid oxidation genes in HF diet-fed mice ··· 83
Ⅳ. DISCUSSION ··· 85
Ⅴ. CONCLUSION ··· 88
REFERENCES ··· 90
LIST OF FIGURES (PART 1)
Fig. 1. Currently proposed signaling pathways for insulin- and contraction-stimulated
glucose transporter 4 (GLUT4) translocation in muscle ··· 3
Fig. 2. TAZ, a transcriptional modulator of enhances osteogenic differentiation and suppresses adipogenic differentiation ··· 4
Fig. 3. FGF21 and KLB are maybe actually good for a lot of things ··· 5
Fig. 4. The GCN2/eIF2α/ATF4/FGF21 signaling pathway ··· 6
Fig. 5. TM-25659 inhibited the PA-induced reduction of ISGU ··· 18
Fig. 6. TM-25659 inhibited the PA-induced reduction in insulin-stimulated phosphor-AKT ··· 19
Fig. 9. Effects of TM-25659 on FGF21 expressions in skeletal muscle cells ··· 23
Fig. 10. TM-25659 induced GCN2-phosphor-eIF2α-ATF4 and FGF21 pathways ··· 24
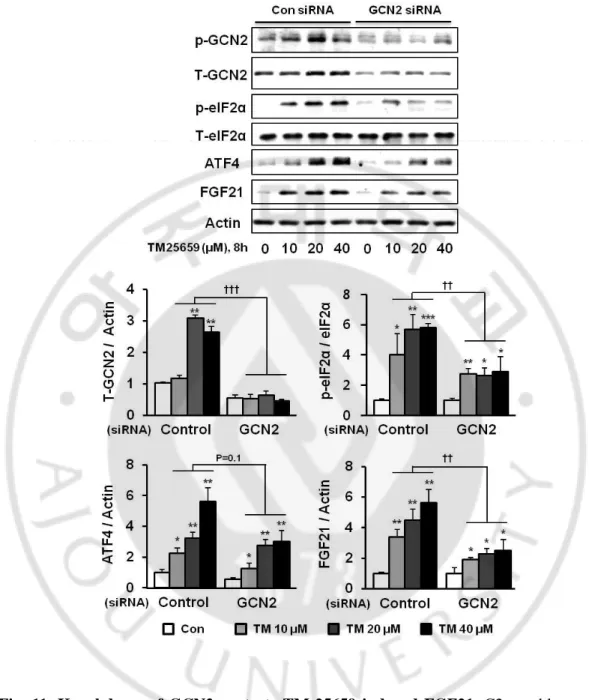
Fig. 11. Knockdown of GCN2 protects TM-25659 induced FGF21 ··· 25
Fig. 12. Effects of TM-25659 on FGF21 expression in PKR and PERK pathways ···· 26
Fig. 13. Knockdown of FGF21 reduced FGF21 levels ··· 28
Fig. 14. TM-25659 prevented PA-induced reduction in insulin stimulated AKT
phosphorylation in C2 myotubes treated with or without FGF21 siRNA ··· 29
Fig. 15. TM-25659 prevented PA-induced pro-inflammatory cytokine
upregulation in C2 myotubes treated with or without FGF21 siRNA ··· 30
Fig. 16. Effects of TM-25659 administration on body weight in DIO mice ··· 32
LIST OF FIGURES (PART 2)
Fig. 1. Obesity update 2012 (from OECD 2012/09/07) ··· 46
Fig. 2. Medical complications of obesity ··· 47
Fig. 3. Diabetes in numbers ··· 48
Fig. 4. TAZ, a transcriptional modulator of enhances osteogenic differentiation
and suppresses adipogenic differentiation ··· 49
Fig. 5. Conditions leading to AMPK activation in higher eukaryotes and some of
its consequences ··· 50
Fig. 6. TM-25659 prevented the PA-induced reduction of insulin signaling pathways ···· ··· 60
Fig. 10. TM-25659 prohibited PA-induced reduction of insulin signaling
in HepG2 cells ··· 66
Fig. 11. TM-25659 prevented PA-induced upregulation of pro-inflammatory cytokine expression in HepG2 cells ··· 68
Fig. 12. TM-25659 improved glucose tolerance and insulin resistance
in HF-diet induced obese mice ··· 70
Fig. 13. TM-25659 prevented reduction of insulin-stimulated AKT phosphorylation In HF diet-fed mice liver ··· 72
Fig. 14. TM-25659 reduced inflammatory cytokine levels in HF diet-fed mice ··· 74
Fig. 15. TM-25659 prevented HF-diet induced liver injury ··· 75
Fig. 16. TM-25659 prevented the HF-diet induced upregulation of pro-inflammatory cytokines in HF diet-fed mice liver ··· 76
Fig. 19. TM-25659 ameliorated hepatic steatosis in HF diet-fed mice ··· 82
Fig. 20. TM-25659 reduced expression of lipogenic genes and increased expression of fatty acid oxidation genes in HF diet-fed mice liver ··· 84
PART 1.
TM-25659-induced activation of FGF21 level
decreases insulin resistance and inflammation
Obesity is attracting attention as a social problem (OECD Obesity Update, 2012). The World Health Organization reported the rate of global obesity has doubled since the 1980s (WHO, 2014). Obesity is closely associated with insulin resistance and other complications, including type 2 diabetes, osteoarthritis, cardiovascular disease, high blood pressure, and cancer (Chobanian, et al., 2003; Hursting, et al., 2007; Poirier, et al., 2006; Pottie , et al., 2006; Seidell, 2000; Visscher and Seidell., 2001). Insulin induces glucose uptake under physiological conditions by stimulating PI3K-AKT pathways and by phosphorylating and reducing the activity of AKT substrate 160, which inhibits glucose transporter 4 translocation to the membrane (Fig. 1) (Sakamoto and Holman., 2008). Insulin resistance is a common physiological state in which higher than normal insulin levels are required to exert physiological effects in peripheral tissues such as the liver, adipose, and skeletal muscle tissues (Woods, et al., 2009).
Transcriptional co-activator with PDZ-binding motif (TAZ) functions as a co-activator, and its activity is regulated via interactions with 14-3-3 protein and PDZ domain proteins (Kanai,
et al., 2000). Increased TAZ activity in the nucleus suppresses adipocyte development and
increases osteoblast differentiation by interacting with peroxisome proliferator-activated receptor gamma (PPAR-γ) and runt-related transcription factor 2 (RUNX2), respectively (Fig. 2) (Park, et al., 2004; Murakami, et al., 2005; Hong, et al., 2005; Lei, et al., 2008; Wang, et al., 2009). The TAZ activator, 2-butyl-5-methyl-6-(pyridine-3-yl)-3-[2'-(1H-tetrazole-5-yl)-biphenyl-4-ylmethyl]-3H-imidazo[4,5-b]pyridine] (TM-25659), inhibits adipocyte development by interacting with PPAR-γ and decreases weight gain in obese mice models (Jang, et al., 2012). However, the fundamental mechanisms underlying the effects of TM-25659 are currently unknown.
Fibroblast growth factor 21 (FGF21) is a member of the FGF family with multiple metabolic functions (Kharitonenkov, et al., 2005; Potthoff, et al., 2012). FGF21 functions as an endocrine hormone with anti-diabetic and anti-obesity effects and is produced in peripheral tissues (e.g., the liver, white and brown adipose tissues, skeletal muscles, and pancreas) (Fig. 3) (Benhamed, et al., 2012; Fisher, et al., 2010; Gimeno and Moller., 2014). Specifically, the administration of recombinant FGF21 or overexpression of FGF21 ameliorates obesity, insulin sensitivity, insulin secretion, inflammation, and glucose clearance in obese or diabetic animals (Kharitonenkov, et al., 2005; Xu, et al., 2009; Adams and Kharitonenkov., 2012; Wang, et al., 2009; Wente, et al., 2006). In contrast, FGF21 deficiency leads to body weight gain, impaired glucose tolerance, elevated blood insulin, and fatty liver development (Badman, et al., 2007; Inagaki, et al., 2007; Kharitonenkov and Shanafelt., 2009).
General control nonderepressible 2 (GCN2) is a serine/threonine protein kinase used to detect amino acid deficiency. Decreased consumption of dietary protein leads to a reduction in the amount of amino acids delivered to the liver, activation of GCN2, and an eventual increase in eIF2-α phosphorylation and activation of ATF4 (Anthony, et al., 2004; Laeger, et
al., 2014). ATF4 binds amino acid response elements (AAREs) in the FGF21 promoter, which leads to an increase of FGF21 in the liver and blood circulation (Fig. 4). FGF21 increases energy expenditure (EE) and decreases body weight gain in mice. However, GCN2 knockout mice fail to exhibit increased eIF2α phosphorylation during both dietary and pharmacological amino acid deprivation, ultimately resulting in hepatic steatosis and liver injury(Anthony, et al., 2004; Guo, et al., 2007; Wilson, et al., 2013).
We found TM-25659 improved insulin signaling and inflammation in PA-induced insulin-resistant skeletal muscle cells. TM-25659 also increased FGF21 mRNA, protein, and secretion levels in association with increased GCN2 in C2 myotubes. TM-25659 increased FGF21 protein levels were reduced by GCN2 siRNA. Additionally, administration of TM-25659 lowered fasting glucose levels, improved insulin resistance, ameliorated inflammation, and increased FGF21 protein levels in our HF-diet induced obesity (DIO) mice models. In this study, we evaluated the reciprocal relationship between TM-25659 and FGF21 in both C2myotubes and mice, and examined the hypothesis that FGF21 mediates the metabolic benefits of TM-25659 by comparing the effects of TM-25659 treated skeletal muscle cells subjected to small interfering RNA (siRNA)-mediated knockdown of FGF21 (FGF21 siRNA) using PA-induced insulin-resistant skeletal muscle cells as controls.
A. MATERIALS
TM-25659 was obtained from the Korea Research Institute of Chemical Technology (Daejeon, South Korea). Bovine serum albumin (BSA), insulin, PA, and methyl cellulose were purchased from Sigma-Aldrich (St. Louis, MO, USA). Anti-AKT, anti-phosphor-AKT (ser473), anti-eIF2α, anti-phosphor-eIF2α (ser51), anti-PERK, anti-phosphor-PERK (Thr980), and anti-ATF4 antibodies were acquired from Cell Signaling Technology (Beverly, MA, USA). Anti-GCN2, anti-phosphor-GCN2 (Thr898) antibodies were acquired from Biorbyt (Cambridge, Cambridgeshire CD4, UK). Anti-FGF21 antibodies were purchased from Abcam (Cambridge, MA, USA). Anti-actin antibody was obtained from Santa Cruz Biotechnology (Santa Cruz, CA, USA). Culture media, culture supplements, horse serum, and fetal bovine serum (FBS) were purchased from Gibco-BRL (Grand Island, NY, USA). Mouse FGF21 and IL-1β ELISA kits were purchased from R&D systems (R&D systems, Minneapolis, MN, USA). TM-25659 was dissolved in 0.5% methyl cellulose in water for in
B. METHODS
1. Preparation of PA
PA/BSA conjugates were prepared by soaping PA with sodium hydroxide (NaOH) and mixing with BSA. Briefly, 20 mM PA in 10 mM NaOH was incubated at 70°C for 30 min, and the fatty acid soaps were combined with 5% fatty acid-free BSA in phosphate-buffered saline (PBS) at a 1:3 volume ratio. The combined fatty acids consisted of 5 mM PA and 3.75% BSA. The PA/BSA conjugates were diluted in 5% horse serum medium (~0.4% BSA) and administered to cultured cells at a final concentration of 500 μM PA. The molar ratio of PA to BSA was ~3.5:1, and the concentration of BSA in the 500 μM PA medium was ~0.7%.
2. Cell culture
C2 mice skeletal myoblasts were obtained from Dr. Hey-Sun Kim (Ajou University, Suwon, South Korea) and grown in high-glucose (4.5 g/L glucose) Dulbecco’s modified Eagle’s medium (DMEM), supplemented with 10% FBS and antibiotics (10 μg/mL streptomycin and 100 IU/mL penicillin), at 37°C in a humidified atmosphere of 95% air and 5% CO2. After
washing the 70–80% confluent myoblasts with PBS, differentiation to myotubes was initiated by transferring the cells to low-serum differentiation medium [DMEM with low
3. Immunoblot analysis
Cells and mice soleus muscles were suspended in RIPA buffer [150 mM sodium chloride (NaCl), 1% NP-40, 0.5% deoxycholate, 0.1% sodium dodecyl sulfate (SDS), 50 mM Tris– HCl, pH 7.5, and a protease inhibitor cocktail (Roche Applied Science, Mannheim, Germany)] and incubated on ice for 20 min. Whole proteins were extracted by differential centrifugation (13,000 × g, 10 min) and the protein concentrations in the lysates were determined using a protein assay kit (Bio-Rad, Hercules, CA, USA). An equal volume of 2× SDS sample buffer (125 mM Tris–HCl, pH 6.8, 4% SDS, 4% 2-mercaptoethanol, and 20% glycerol) was added to the cell lysates and equivalent amounts of protein (20 μg) were loaded onto 8–12% polyacrylamide gels, electrophoresed, and transferred electrophoretically to polyvinylidene fluoride membranes (Millipore, Billerica, MA, USA). After blocking the membranes with 5% skim milk for 30 min, the target antigens were reacted with primary antibodies, followed by secondary antibodies (horseradish peroxidase-conjugated anti-goat IgG, or anti-rabbit IgG). Immunoreactive bands were visualized by enhanced chemiluminescence (Amersham Pharmacia Biotech, Arlington Heights, IL, USA).
4. RNA isolation and quantitative real-time polymerase chain reaction (PCR)
Total RNA was isolated from cell or muscle tissues and prepared using RNAiso Plus reagent (TaKaRa Bio Inc., Otsu, Japan) according to the manufacturer’s instructions. Briefly, C2 myotube cDNA was synthesized with avian myeloblastosis virus reverse transcriptase using random 9-mers. The cDNA was amplified by PCR with primer sets for FGF21 [CCC CAA ACC AGG ATG CAA CA (forward, F) and ATG CTG CTT GGT CTT GGG GC (reverse, R)], tumor necrosis factor (TNF)-α [GGC ACT CCC CCA AAA GAT GG (F) and CCC TGC CAC AAG CAG GAA TG (R)], interleukin (IL)-6 [CCA TCC AGT TGC CTT CTT GGG (F) and GCC GTG GTT GTC ACC AGC AT (R)], IL-1β [TCT CGC AGC AGC ACA TCA ACA (F) and CCT GGA AGG TCC ACG GGA AA (R)], monocyte chemoattractant protein (MCP)-1 [CAG CCA GAT GCA GTT AAC GC (F) and GCC TAC TCA TTG GGA TCA TCT (R)], and RPL32 [AAG CGA AAC TGG CGG AAA CC (F) and CCC ATA ACC GAT GTT GGG CA (R)]. Quantitative real-time PCR was performed using SYBR Green (TaKaRa Bio Inc.) with a TaKaRa TP-815 instrument. All expression values were normalized to RPL32 mRNA levels.
5. siRNAs
Twenty-one nucleotide siRNA duplexes were designed and created by Bioneer (Daejeon, South Korea). The siRNA sequences were as follows: green fluorescent protein, 5′-GUU CAG CGU GUC CGG CGA GTT-3′, FGF21, 5′-CUG AUG GAA UGG AUG AGA U-3′, GCN2, 5′-GUG GAA GCU AAA UUC UGU A-3′, PERK, 5′-GGG ACA AGT AGG GAC CAA G-3′ and PKR, 5′-TGG AAA ATC CCG AAC AAG-3′. C2 myotubes were transfected with siRNA oligonucleotides using a pipette-type electroporator (Microporator-Mini; Digital Biotechnology, Seoul, South Korea) according to the manufacturer’s instructions. Briefly, 1 × 106 C2 myotubes were transfected with 1 nM siRNA (1 μg in 10 μL of R buffer) by microporation at a pulse voltage of 1005 V, pulse width of 35 ms, and pulse number of 2. After transfection, the C2 myotubes were differentiated by switching the medium to differentiation medium for 3 days. Following differentiation, the cells were seeded in 12-well plates at a concentration of 1 × 105 cells/well and treated with or without TM-25659 for 16 h.
6. Uptake of 2-N-(7-nitrobenz-2-oxa-1,3-diazol-4-yl) amino-2-deoxyglucose (2-NBDG) C2 myotubes were pretreated with PA (500 μM) and TM-25659 (25 or 50 μM) for 16 h. Next, the cells were starved for 4 h and then preincubated in Krebs-Ringer bicarbonate buffer (pH 7.4) containing 2% BSA at 37°C for 30 min. They were then treated with 500 μM 2-NBDG (cat. no. N13195; Invitrogen, Carlsbad, CA, USA) with or without 100 nM insulin at 37°C for 2 h. The cells were washed with ice-cold PBS three times and lysed with cell lysis buffer. The lysates were centrifuged at 13,000 rpm at 4°C for 20 min. The supernatants were measured for fluorescence (excitation 475 nm, emission 550 nm) using a SpectraMax Gemini EM microplate reader (Molecular Devices, Sunnyvale, CA, USA). Protein concentrations were determined by Bradford assay.
7. Animal experiments
Six-week-old male C57BL/6J mice were purchased from Japan SLC Inc. (Hamamatsu, Japan). All animal experiments were approved by the Animal Ethics Committee of the Laboratory Animal Research Center, Ajou University Medical Center (Suwon, South Korea). The mice were housed in a temperature-controlled room (22 ± 2°C) with a 12/12 h light/dark cycle and fed ad libitum. Following 2 weeks of adaptation, the mice were fed a HF diet (n = 12) for 6 weeks. The mice were provided a HF diet with 60% kcal from fat (Research Diets, New Brunswick, NJ, USA) ad libitum. The HF group was randomly assigned during the
8. FGF21 secretion
The concentration of FGF21 in the culture medium of C2 myotubes was determined using a mouse/rat FGF21 enzyme-linked immunosorbent assay (ELISA) (R&D Systems, Minneapolis, MN, USA).
9. IL-1β assay
The concentration of 1β in the cultured C2 myotubes was determined using a mouse IL-1β enzyme-linked immunosorbent assay (ELISA) (R&D Systems, Minneapolis, MN, USA).
10. Statistical analysis
Data are presented as the mean ± standard error of at least three independent experiments. Statistical differences between the groups were determined using Student’s t-test and Fisher’s exact test. P-values < 0.05 were considered significant.
A. TM-25659 restored PA-induced insulin resistance and inflammation in skeletal muscle cells
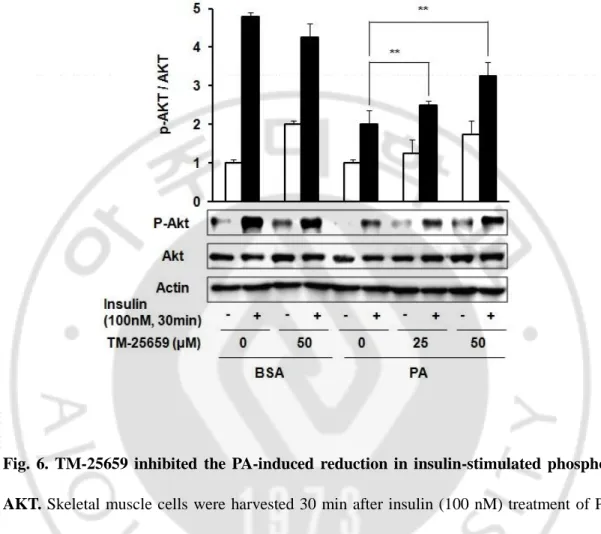
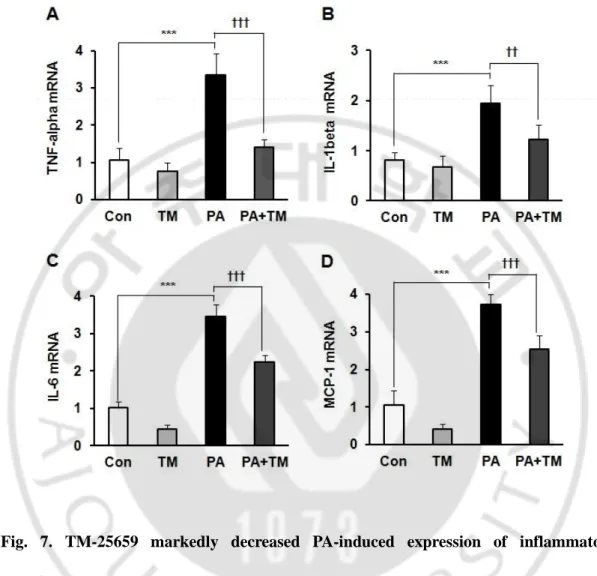
In previous experiments, we found 500 µM PA decreased insulin-stimulated glucose uptake (ISGU) in C2 myotubes. (Jung, et al., 2011). In this study, we investigated the effects of TM-25659 on the PA-induced ISGU in C2 myotubes. TM-TM-25659 prevented the decrease in ISGU in C2 myotubes (Fig. 5). Additionally, we investigated the effects of TM-25659 on the reduction of insulin-stimulated AKT phosphorylation by PA in C2 myotubes. TM-25659 significantly prevented the PA-induced reduction in insulin-stimulated AKT phosphorylation (Fig. 6). These data suggest TM-25659 restored the PA-induced decrease in insulin sensitivity. PA increased pro-inflammatory cytokine gene expressions in C2 myotubes, while TM-25659 markedly prevented the PA-induced expressions of TNF-α, IL-1β, IL-6, and MCP-1 in C2 myotubes (Fig. 7A-D). Additionally, the PA-induced IL-1β maturation levels decreased in C2 myotubes in the PA+25659 group (Fig. 8). These data demonstrate TM-25659 prevented the PA-induced expression of pro-inflammatory cytokines in C2 myotubes.
Fig. 5. TM-25659 inhibited the PA-induced reduction of ISGU. Skeletal muscle cells were harvested 30 min after insulin (100 nM) treatment of PA, TM-25659, or vehicle-treated cells for 16 h. TM-25659 prevented PA-induced impaired glucose uptake by 2-NBDG in C2 myotubes. The data are given as means ± standard error. *P < 0.05, **P < 0.01, †††P < 0.001.
Fig. 6. TM-25659 inhibited the PA-induced reduction in insulin-stimulated phosphor-AKT. Skeletal muscle cells were harvested 30 min after insulin (100 nM) treatment of PA, TM-25659, or vehicle-treated cells for 16 h. Representative immunoblots showing phosphor (Ser 473) and total AKT in skeletal muscle cells. The data are given as means ± standard error. **P < 0.01.
Fig. 7. TM-25659 markedly decreased PA-induced expression of inflammatory cytokines. Skeletal muscle cells were harvested of PA, TM-25659, or vehicle-treated cells for 16 h. (A–D) TNF-α, IL-1β, IL-6, and MCP-1 mRNA expression as quantified by real-time PCR. [Con, BSA; TM, 25 or 50 µM TM-25659; PA, 500 µM Palmitate]. The data are given as means ± standard error. ***P < 0.001, ††P < 0.01, †††P < 0.001.
Fig. 8. TM-25659 inhibited the PA-induced upregulation of IL-1β levels in skeletal muscle cells. C2 myoblasts were differentiated into C2 myotubes and treated with PA, TM-25659, or vehicle. Levels of IL-1β as quantified by enzyme-linked immunosorbent assays. The data are given as means ± standard error. *P < 0.05, **P < 0.01.
B. TM-25659 increased FGF21 levels by GCN2 pathways in skeletal muscle cells
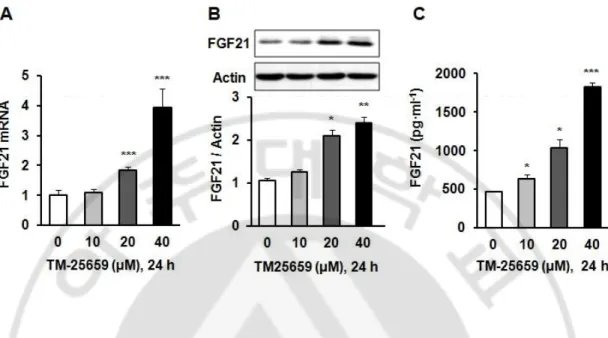
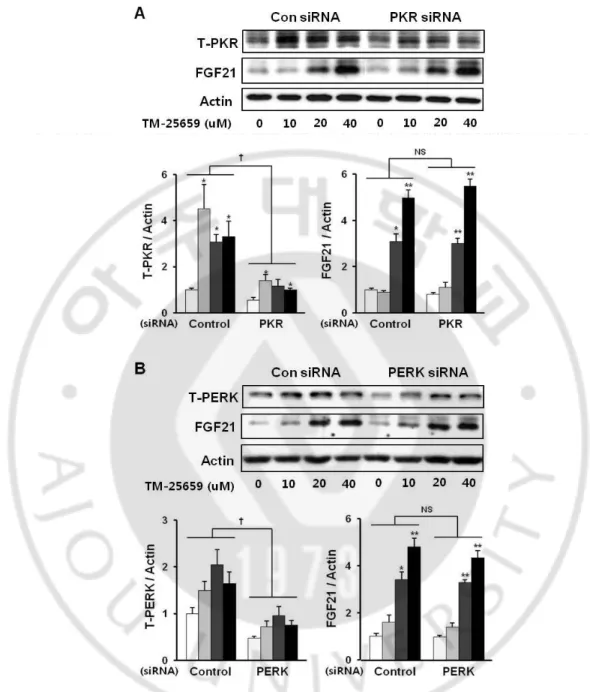
We found TM-25659 increased FGF21 mRNA, protein, and secretion levels in C2 myotubes. Importantly, TM-25659 significantly increased FGF21 mRNA expression in C2 myotubes in a dose-dependent manner (Fig. 9A). We also observed increased FGF21 protein levels in C2 myotubes treated with TM-25659 (Fig. 9B). Moreover, TM-25659 increased FGF21 secretion in C2 myotubes (Fig. 9C). These data show TM-25659 induced FGF21 mRNA and protein expression and FGF21 secretion in C2 myotubes. TM-25659 induced increased FGF21 protein levels through GCN2 pathways (GCN2-phosphor-eIF2α-ATF4 and FGF21) (Fig. 10). Therefore, we next investigated whether TM-25659 increased GCN2 pathways in C2 myotubes that were or were not treated with GCN2 siRNA. We found TM-25659 induced GCN2 pathways, but this induction was diminished by GCN2 siRNA (Fig. 11). We also investigated the involvement of other pathways such as PERK and PKR in TM-25659-induced FGF21 expressions in C2 myotubes. We found TM-25659 induced the induction of PERK and PKR, while FGF21 induction was not reduced by PERK of PKR siRNA treatment (Fig. 12A and 12B). These data indicate TM-25659 induced increases in FGF21 levels through GCN2 pathways in C2 myotubes.
Fig. 9. Effects of TM-25659 on FGF21 expressions in skeletal muscle cells. C2 myoblasts were differentiated into C2 myotubes and treated with 10–40 µM TM-25659 or vehicle. (A) FGF21 mRNA expression levels as quantified by real-time PCR. (B) FGF21 protein levels by immunoblotting. (C) Secretion of FGF21 as quantified by enzyme-linked immunosorbent assays. The data are given as means ± standard error. *P < 0.05, **P < 0.01, ***P < 0.001.
Fig. 10. TM-25659 induced GCN2-phosphor-eIF2α-ATF4 and FGF21 pathways. C2 myoblasts were differentiated into C2 myotubes and treated with 40 µM TM-25659 for 0, 2, 4, 8, 12 h. GCN2 pathway protein levels by immunoblotting.
Fig. 12. Effects of TM-25659 on FGF21 expression in PKR and PERK pathways. C2 myoblasts were differentiated into C2 myotubes and treated with 10, 20, or 40 µM of
TM-C. Effects of TM-25659 reduced insulin resistance and inflammation in skeletal muscle cells treated with or without FGF21 siRNA
We previously showed TM-25659 markedly increased FGF21 mRNA and protein expressions in TM-25659 treated skeletal muscle cells (Fig. 9). We investigated the effects of TM-25659 on PA-induced insulin resistance and inflammation in C2 myotubes with or without FGF21 siRNA treatment. FGF21 mRNA and protein expressions decreased following FGF21 siRNA treatment (Fig. 13A and 13B). TM-25659 prevented PA-induced decrease in insulin-stimulated AKT phosphorylation, and the beneficial effects of TM-25659 decreased following FGF21 siRNA treatment (Fig. 14). We next investigated whether TM-25659 prevented the PA increased levels of pro-inflammatory cytokines in C2 myotubes treated with or without FGF21 siRNA. We found TM-25659 prevented PA increased pro-inflammatory cytokine expression (MCP-1, TNF-α, IL-1β and IL-6), but the TM-25659-induced decrease in induction was lowered by FGF21 siRNA (Fig. 15A-D). Consistent with our observations in C2 myotubes, TM-25659 significantly decreased the PA induced expression of pro-inflammatory cytokines in C2 myotubes, whereas these effects were abolished by FGF21 siRNA.
Fig. 13. Knockdown of FGF21 reduced FGF21 levels. C2 myoblasts were differentiated into C2 myotubes and treated with or without FGF21 siRNA. (A) FGF21 mRNA expression with or without FGF21 siRNA as quantified by real-time PCR. (B) Representative immunoblots showing FGF21 in skeletal muscle cells with or without FGF21 siRNA. The data are given as means ± standard error. †††P < 0.001.
Fig. 14. TM-25659 prevented PA-induced reduction in insulin stimulated AKT phosphorylation in C2 myotubes treated with or without FGF21 siRNA. Skeletal muscle cells were harvested 30 min after insulin (100 nM) treatment of PA, TM-25659, or vehicle-treated cells for 16 h with or without FGF21 siRNA. Representative immunoblots showing phosphor (Ser 473) and total AKT in skeletal muscle cells of the control (Con), TM-25659 (TM), palmitate (PA), and PA+TM groups with or without FGF21. The data are given as means ± standard error. **P < 0.01, ††P < 0.01.
Fig. 15. TM-25659 prevented PA-induced pro-inflammatory cytokine upregulation in C2 myotubes treated with or without FGF21 siRNA. TNF-α, IL-1β, IL-6, and MCP-1 mRNA expression in C2 myotubes treated with or without FGF21 siRNA as quantified by real-time PCR. Skeletal muscle cells were harvested 30 min after insulin (100 nM) treatment of PA treated cells with TM-25659 or vehicle for 16 h. The data are given as means ±
D. TM-25659 prevented HF-diet induced obesity
The mice were fed a HF diet for 6 weeks to examine the metabolic effects of TM-25659 treatment before any effects occurred on body weight. Mice in the HF group were randomly divided into two groups for the next 8 weeks: the HF and HF+TM-25659 groups. Body weight gain was increased by the high-fat diet and this increase was decreased in the TM-25659 treated HF+TM group compared to a vehicle treated HF group which can be recognized with the naked eye (Fig. 16A and B). We investigated the food intake in order to understand the body weight gain loss mechanisms of TM-25659. Daily food intake showed no significant differences between HF and HF+TM mice on a high-fat diet (Fig. 16C). These results suggested TM-25659 attenuates weight gain, regardless of food intake.
Fig. 16. Effects of TM-25659 administration on body weight in DIO mice. C57BL/6J mice were fed HF for 6 weeks, separated into two groups (n = 8 for each group) and then orally administrated every other day with either vehicle or TM-25659 (50 mg/kg) for 8 weeks. (A) Body weight measured biweekly. HF+TM group is less than HF group, increased body weight. (B) A representative photograph of decreased weight gain in HF+TM group mice. (C) Food intake was similar between the HF and HF+TM group. (ND – Normal chow
E. TM-25659 restored the insulin signaling pathways in DIO mice skeletal muscle DIO mice were administered TM-25659 to test its beneficial effects on HF-diet induced insulin signaling in skeletal muscles. The insulin-stimulated ND group had markedly increased levels of phosphorylated AKT in skeletal muscles, whereas those in the HF group decreased markedly. The HF+TM group showed increased levels of insulin-stimulated AKT phosphorylation in skeletal muscles, comparable to those in the HF group (Fig. 17). These results suggest TM-25659 prevented the HF-diet induced reduction in insulin signaling.
Fig. 17. TM-25659 prevented HF-diet induced reduction of insulin-stimulated AKT phosphorylation in mice skeletal muscle. C57BL/6J mice were fed HF for 6 weeks, separated into two groups and then orally administrated every other day with either vehicle or TM-25659 (50 mg/kg) for 8 weeks. Representative immunoblots showing phosphor (Ser 473) and total AKT in mice skeletal muscles of the ND, HF, and HF+TM group mice. (ND – Normal-chow diet, HF – High-fat diet, HF+TM – High-fat diet+TM-25659) The data are given as means ± standard error. *P<0.05, **P<0.01, †††P < 0.001.
F. TM-25659 lowered the muscle pro-inflammatory cytokine levels in DIO mice
TM-25659 protected against pro-inflammatory cytokine gene expression in HF-diet fed mice. The HF group showed increased levels of TNF-α, IL-1β, MCP-1, and IL-6 expressions in skeletal muscles. The HF+TM group showed decreased levels of TNF-α, IL-1β, MCP-1, and IL-6 mRNA expressions in skeletal muscles, comparable to those in the HF group. (Fig. 18A-D). These data suggest TM-25659 improved the HF-diet induced pro-inflammatory cytokine activity in C57BL/6J mice skeletal muscle.
Fig. 18. TM-25659 prevented HF diet-induced upregulation of pro-inflammatory cytokines. C57BL/6J mice were fed HF for 6 weeks, separated into two groups and then orally administrated every other day with either vehicle or TM-25659 (50 mg/kg) for 8 weeks. (A-D) TNF-α, IL-6, MCP-1, and IL-1β mRNA expressions in skeletal muscles are measured by way of quantification using real-time PCR (ND – Normal chow diet, HF – High-fat diet, HF+TM – High-fat diet+TM-25659). The data shown are means ± standard error. *P < 0.05, **P<0.01, ***P < 0.001 vs. vehicle in ND group mice. †P < 0.05, ††P <
G. TM-25659 regulates expression of both FGF21 mRNA and protein in DIO mice skeletal muscles
Because FGF21 was markedly expressed in skeletal muscle cells, we investigated the effects of TM-25659 on FGF21 expression in mice skeletal muscles. After TM-25659 treatment, the FGF21 mRNA expression significantly increased, and the FGF21 protein levels significantly increased (Fig. 19A and 19B). Therefore, TM-25659 is able to induce FGF21 expression.
Fig. 19. TM-25659 increases FGF21 levels. C57BL/6J mice were fed HF for 8 weeks, separated into two groups and then orally administrated every other day with either vehicle
In this study, treating skeletal insulin resistant muscle cells with the TM-25659, known as TAZ activator protected against PA-induced insulin resistance and inflammation. TM-25659 increased the FGF21 mRNA and protein levels in HF-diet induced mice skeletal muscles and C2 myotubes. Additionally, TM-25659 increased FGF21 mRNA expressions and stimulated FGF21 secretion in C2 myotubes. But, TM-25659 did not induced expression of FGF21 mRNA and protein in liver. When skeletal muscle cells were treated with FGF21 siRNA, TM-25659 did not protect against PA induced insulin resistance and inflammation. Additionally, we demonstrated that TM-25659 administration to HF-diet induced obese mice improved fasting glucose levels and insulin sensitivity. Thus, the beneficial effects of TM-25659 must be associated with increased FGF21 in skeletal muscles. Improved body weight following treatment with TM-25659 has been reported (Jang, et al., 2012), but neither muscle specific insulin sensitivity nor the relationships between FGF21 and TM-25659 have not been examined. FGF21 is a metabolic hormone produced mainly by the liver, but it is also expressed in muscle and adipose tissue where it regulates glucose and lipid homeostasis via pleiotropic actions (Woo, et al., 2013). Here, we demonstrated TM-25659 increased FGF21 RNA and protein levels directly as dose- and time-dependent variables, and that FGF21 was secreted at significantly increased levels into mice skeletal muscle cell medium. To test these effects in mice, TM-25659 was injected into C57BL6J mice every other day for
activating transcription factor 4 (ATF4) from the cytoplasm to the nucleus in autophagy-deficient skeletal muscles. In addition, FGF21 was found to be induced in the skeletal muscles under leucine or methionine deprivation through the GCN2-phosphor-eIF2-α and ATF4 pathways (De Sousa-Coelho, et al., 2013; Lees, et al., 2014; Wan, et al., 2014). We investigated TM-25659-induced changes in amino acid levels. We found that TM-25659 resulted in GCN2 activation by leucine or methionine deprivation (data not shown). TM-25659 induced FGF21 does react in the same way in the GCN2-phosphor-eIF2-α and ATF4 pathways. But the relationship of TAZ and GCN2 is unknown. TM-25659 improved insulin resistance in HF-diet fed mice muscles. Mounting evidence suggests macrophage infiltration and accumulation of chronic inflammatory cytokines such as TNF-α, IL-1β, IL-6, or MCP-1 in adipose tissues are positively correlated with insulin resistance (Weisberg, et al., 2006; Xu,
et al., 2003; Gao, et al., 2013; Ota, et al., 2013). Because TM-25659 reduced inflammatory
cytokines such as TNF-α, IL-1β, IL-6, and MCP-1, TM-25659 is protective against insulin resistance in mice fed a HF-diet. Another cause of reduced insulin resistance was FGF21 expression following TM-25659 treatment of muscles. Xu, et al. (2009) reported a FGF21 infusion given to DIO mice protected against insulin resistance by enhancing EE and lipid oxidation. Furthermore, transgenic mice overexpressing hepatic FGF21 were protected from HF-diet induced insulin resistance (Kharitonenkov, et al., 2005; Inagaki, et al., 2007). However, FGF21 knockdown mice had reduced adiponectin levels and aggravated chronic
the accumulation of FGF21 in skeletal muscle and protected against insulin resistance and chronic inflammation in muscles.
Our results suggest TM-25659 administered to HF-diet induced obese mice prevented insulin resistance and inflammation. Also, TM-25659 prevented PA-induced insulin resistance and inflammation in C2 myotubes. These beneficial effects were mediated via the induction of FGF21 expression.
PART 2.
Protective effects of TM-25659 on high-fat
diet-induced insulin resistance in liver via
According to the World Health Organization, as of 2012, the obese population is currently 1 billion people. The worldwide obese population is increasing continuously. Also, obesity update 2012 (from OECD 2012/09/07) reported at least one out of every two people are currently obese or overweight in over half of the OECD countries (Fig. 1).
Fig. 1. Obesity update 2012 (from OECD 2012/09/07). ( http://www.oecd.org/els/health-systems/49716427.pdf)
In addition, obesity is known to cause a variety of chronic degenerative diseases such as hypertension, inflammation, insulin resistance, type 2 diabetes, fatty liver, and other health problems. (Rahmouni, et al., 2005; Kotsis, et al., 2010; Wellen and Hotamisligil., 2003; Shoelson, et al., 2007; Hotamisligil, et al., 1996; Barbare and Jeffrey., 2000; Mokdad, et al., 2003; Eckel, et al., 2011; Wanless and Lentz., 1990; Fabbrini, et al., 2010). The orient has also seen an increase in obesity due mainly in adopting a westernized lifestyle (Couch, et al., 2000; Hu., 2011; Kim, et al., 2011; Egger and Dixon., 2014). Obesity is important because these serious complications are a result of obesity rather than obesity itself (Fig. 2).
Diabetes is one of the diseases caused by obesity and diabetes in the population is also increasing continuously. According to data from Nature (2012), the diabetes population has reached 346 million people. The incidence of diabetes in high income countries has reached 8% and the prevalence of diabetes in the upper middle or middle income countries has reached 10% (Fig. 3).
Nature 2012, 485, S2-S3 (2012/05/17) Fig. 3. Diabetes in numbers. (Tony Scully., 2012)
Obesity therapeutic agents are currently commercially available, as well as diabetes therapeutic agents. The current agents have negative side effects. The development of new therapeutic agents are needed. This study, along with a co-investigator from the Research Institute of Chemical Technology (PhD. Ann), facilitates the movement of the screening of low-molecular-weight substances that regulate the movement to the nucleus of the TAZ protein from a library of low-molecular-weight compounds, TAZ gene expression and protein activity within the nucleus substance that controls were uncovered 2-butyl-5-methyl-
6-(pyridine-3-yl)-3-[2'-(1H-tetrazole-5-yl)-biphenyl-4-ylmethyl]-3H-imidazo[4,5-b]pyridine] (TM-25659). In the previous study, TM-25659, a low molecular weight compound, does not affect the expression or promotion of TAZ proteins, but increases migration cytoplasm to the nucleus of TAZ proteins. Also, TM-25659 inhibits 3T3-L1 adipocyte differentiation and decreases weight gain in obese mice models (Harada and Rodan., 2003; Hong, et al., 2005; Jang, et al., 2012) (Fig. 4).
AMP-activated protein kinase (AMPK) is a serine/threonine kinase that controls cellular energy homeostasis. It stimulates ATP-producing pathways and diminishes energy consumption (Fig. 5). AMPK inhibits ER and oxidative stress, inflammation, gluconeogenesis, lipogenesis, and activates cellular glucose uptake, and beta-oxidation of fatty acid, all of which appear to be involved in the pathogenesis of insulin resistance (Terai,
et al., 2005; Shin, et al., 2009; O’Neill, 2013; O’Neill, et al., 2013; Kim, et al., 2008; Park, et al., 2008; Hardie, et al., 2002). Recently, AMPK has been implicated in a regulator of
inflammatory responses that treatment with AMPK activators inhibits palmitate-induced expression of pro-inflammatory cytokines in most cell types (Su, et al., 2007; Katerelos, et
Our results show beneficial effects of TM-25659 on DIO mice and PA-induced insulin-resistant HepG2 cells. TM-25659 increased AMPK and decreased PA-induced insulin resistance through AMPK-mediated pathways in HepG2 cells. Additionally, administration of TM-25659 increased the AMPK and ACC phosphorylation levels in DIO mice. In this study, we investigated the administration of a TM-25659 ameliorated weight gain, suppressed insulin resistance, inflammation, and hepatic steatosis in HF DIO mice. Also, we investigated TM-25659 improved insulin resistance and underlying molecular mechanism via AMPK mediated pathways in PA-induced insulin-resistant HepG2 cells.
A. MATERIALS
TM-25659 was obtained from the Korea Research Institute of Chemical Technology (Daejeon, South Korea). Bovine serum albumin (BSA), insulin, PA, and methyl cellulose were purchased from Sigma-Aldrich (St. Louis, MO, USA). Anti-IRS, anti-phosphor-IRS (Tyr612) antibodies, anti-AKT AKT (Ser473), anti-AMPK, anti-phosphor-AMPK (Thr172), anti-ACC, anti-phosphor-ACC (Ser79) antibodies were acquired from Cell Signaling Technology (Beverly, MA, USA). Anti-actin and anti-tubulin antibodies were obtained from Santa Cruz Biotechnology (Santa Cruz, CA, USA). Culture media, culture supplements, horse serum, and fetal bovine serum (FBS) were purchased from Gibco-BRL (Grand Island, NY, USA). Mouse IL-1β ELISA kit was purchased from R&D systems (R&D systems, Minneapolis, MN, USA). TM-25659 was dissolved in 0.5% methyl cellulose in sterilized water for in vivo administration.
B. METHODS
1. Preparation of PA
PA/BSA conjugates were prepared by soaping PA with NaOH and mixing with BSA. Briefly, 20 mM PA in 10 mM NaOH was incubated at 70°C for 30 min, and the fatty acid soaps were combined with 5% fatty acid-free BSA in phosphate-buffered saline (PBS) at a 1:3 volume ratio. The combined fatty acids consisted of 5 mM PA and 3.75% BSA. The PA/BSA conjugates were diluted in Dulbecco’s modified Eagle’s (DMEM) medium containing 10% FBS (approximately 0.4% BSA) and administered to cultured cells at a final concentration of 250 or 500 μM PA. The molar ratio of PA to BSA was approximately 3.5:1, and the concentration of BSA in the 250 or 500 μM PA medium was approximately 0.7%.
2. Cell culture
HepG2 cells were obtained from ATCC and grown in Dulbecco’s modified Eagle’s medium (DMEM), supplemented with 10% FBS and antibiotics (10 μg/mL streptomycin and 100 IU/mL penicillin), at 37°C in a humidified atmosphere of 95% air and 5% CO2.
3. Immunoblot analysis
Rad, Hercules, CA, USA). An equal volume of 2× SDS sample buffer (125 mM Tris-HCl, pH 6.8, 4% SDS, 4% 2-mercaptoethanol, and 20% glycerol) was added to the cell lysates, and equivalent amounts of protein (20 μg) were loaded onto 8–12% polyacrylamide gels, electrophoresed, and transferred electrophoretically to polyvinylidene fluoride membranes (Millipore). After blocking the membranes with 5% skim milk for 30 min, the target antigens were reacted with primary antibodies, followed by secondary antibodies (horseradish peroxidase-conjugated anti-goat IgG, or anti-rabbit IgG). Immunoreactive bands were visualized by enhanced chemiluminescence (Amersham Pharmacia Biotech, Arlington Heights, IL, USA).
4. RNA isolation and quantitative real-time polymerase chain reaction (PCR)
Total RNA was isolated from cell or liver tissues and prepared using RNAiso Plus reagent (Takara Bio Inc., Otsu, Japan) according to the manufacturer’s instructions. Briefly, HepG2 cells cDNA were synthesized with avian myeloblastosis virus reverse transcriptase using random 9-mers. The cDNA was amplified by PCR with primer sets for mousetumor necrosis factor (TNF)-α [GGC ACT CCC CCA AAA GAT GG (forward, F) and CCC TGC CAC AAG CAG GAA TG (reverse, R)], mouse interleukin (IL)-6 [CCA TCC AGT TGC CTT CTT GGG (forward, F) and GCC GTG GTT GTC ACC AGC AT (reverse, R)], mouse IL-1β
GGA CG (forward, F) and TGA GGA ACA AGC ACC GCC TG (reverse, R)], human interleukin (IL)-6 [TGT GTG GGG CGG CTA CAT CT (forward, F) and GCC TTC GGT CCA GTT GCC TT (reverse, R)], human IL-1β [CCT TTG GTC CCT CCC AGG AA (forward, F) and TGA GTC TGC CCA GTT CCC CA (reverse, R)], human monocyte chemoattractant protein (MCP)-1 [TGA AAG TCT CTG CCG CCC TTC (forward, F) and ATC TGG CTG AGC GAG CCC TT (reverse, R)], and human RPL32 [TGG CAT TGG GGT TGG TGA CT (forward, F) and TGC CGA GAT CGC TCA CAA TG (reverse, R)].
Quantitative real-time PCR was performed using SYBR Green (Takara Bio Inc.) with a Takara TP-815 instrument. All expression values were normalized to RPL32 mRNA levels.
5. Animal experiments
Six-week-old male C57BL/6J mice were purchased from Japan SLC Inc. (Hamamatsu, Japan). All animal experiments were approved by the Animal Ethics Committee of the Laboratory Animal Research Center, Ajou University Medical Center (Suwon, South Korea). The mice were housed in a temperature-controlled room (22 ± 2°C), with a light/dark cycle of 12 h and fed ad libitum. Following 2 weeks of adaptation, the mice were fed a HF diet (n = 12) for 6 weeks. The mice were provided a HF diet with 60% kcal from fat (Research Diets, New Brunswick, NJ, USA) ad libitum. The HF group was randomly assigned during the additional 8 weeks to two groups: HF and HF+50 mg/kg TM-25659. The mice were
6. Insulin tolerance tests
The intraperitoneal insulin tolerance tests (ITT) were performed by intraperitoneally injecting regular human insulin (0.5 U/kg) after a 6-h fast. Blood samples were taken at different time points (0, 30, 60, and 120 min after insulin loading) from a tail vein. Plasma glucose was measured using Accu-chek (Korea Roche Diagnostics, Seoul, Korea).
7. Glucose tolerance tests
The oral Glucose tolerance tests (OGTT) were performed by oral glucose administration (2 g/kg) after a 6-h fast. Blood samples were taken at different time points (0, 30, 60, and 120 min after glucose loading) from a tail vein. Plasma glucose was measured using Accu-chek (Korea Roche Diagnostics, Seoul, Korea).
8. Statistical analysis
Data are presented as the mean ± standard error of at least three independent experiments. Statistical differences between the groups were determined using Student’s t-test and Fisher’s exact test. P-values < 0.05 were considered significant.
A. TM-25659 restored PA-induced reduction of insulin signaling pathway and induction of inflammation in HepG2 cells
In previous experiments, we found 500 µM PA decreased insulin-stimulated insulin signaling pathways in HepG2 cells. We investigated the effects of TM-25659 on the reduction of insulin-stimulated IRS and AKT phosphorylation by PA in HepG2 cells. HepG2 cells were treated with TM-25659 in the presence of PA and then stimulated with insulin. TM-25659 inhibited the PA-induced reduction in insulin-stimulated IRS and AKT phosphorylation significantly (Fig. 6). These data suggest TM-25659 restored the PA-induced decrease of insulin signaling pathways in HepG2 cells. In addition, TM-25659 protected against pro-inflammatory cytokine gene expressions in PA-induced upregulation of inflammation in HepG2 cells. The PA group showed increased levels of TNF-α, IL-1β, MCP-1, and IL-6 expressions in HepG2 cells. The PA+TM group showed decreased levels of TNF-α, IL-1β, MCP-1, and IL-6 mRNA expressions in HepG2 cells, comparable to those in the PA group. (Fig. 7A-D). These data suggest TM-25659 improved the PA-induced pro-inflammatory cytokine activity in HepG2 cells.
Fig. 6. TM-25659 prevented the PA-induced reduction of insulin signaling pathways. HepG2 cells were harvested 30 min after insulin (100 nM) treatment of PA, TM-25659, or vehicle-treated cells for 12 h. Representative immunoblots showing phosphor (Tyr 612), total IRS, phosphor (Ser 473), and total AKT in HepG2 cells. The data are given as means ± standard error. *P < 0.05, **P < 0.01, ††P < 0.01.
Fig. 7. TM-25659 reduced the PA-induced expressions of inflammatory cytokines. HepG2 cells were harvested of PA, TM-25659, or vehicle-treated cells for 12 h. (A–D) TNF-α, IL-1β, MCP-1, and IL-6 mRNA expressions as quantified by real-time PCR. [Con, BSA; TM, 50 µM TM-25659; PA, 500 µM Palmitate]. The data are given as means ± standard
B. TM-25659 increased AMPK and ACC phosphorylation in HepG2 cells
We next examined whether TM-25659 was related to AMPK and ACC phosphorylation. Expression of AMPK and ACC in HepG2 cells increased with TM-25659 treatment (Fig. 8). The PA group showed decreased AMPK and ACC phosphorylation in HepG2 cells. The PA+TM group showed increased levels of phosphorylated AMPK and ACC in HepG2 cells, comparable to those in the PA group (Fig. 9). TM-25659 significantly induced AMPK and ACC phosphorylation, however these effects did not occur in AMPK inhibitor compound C treated cells (Fig. 9). These data suggest TM-25659 induced AMPK phosphorylation in HepG2 cells.
Fig. 8.TM-25659 increased phosphorylation of AMPK and ACC in HepG2 cells. HepG2 cells were harvested from TM-25659 treated cells for 24 h. Representative immunoblots showing phosphor-ACC, total ACC, phosphor-AMPK, and total AMPK in HepG2 cells of the control and TM treated cells, respectively. The data are given as means ± standard error. *P < 0.05, **P < 0.01.
Fig. 9. TM-25659 prohibited PA-induced reduction of AMPK and ACC phosphorylation in HepG2 cells. HepG2 cells were harvested 12 h after compound C (10 uM) and PA treatment of TM-25659-treated cells for 24 h. Representative immunoblots
C. TM-25659 reduced PA-induced reduction of insulin signaling pathways in HepG2 cells
TM-25659 significantly inhibited PA-induced reduction of insulin signaling pathways in HepG2 cells. TM-25659 mediated inhibition of PA-induced reduction of IRS and AKT phosphorylation in HepG2 cells and was significantly reduced by compound C, an AMPK inhibitor (Fig. 10). These data showed TM-25659 inhibited PA-induced insulin resistance via the AMPK signaling in HepG2 cells.
Fig. 10.TM-25659 prohibited PA-induced reduction of insulin signaling in HepG2 cells. HepG2 cells were harvested 30 m after insulin (100 nM) treatment of PA, compound C,
TM-D. TM-25659 reduced PA-induced reduction of inflammation in HepG2 cells
TM-25659 significantly inhibited PA-induced upregulation of pro-inflammatory cytokines in HepG2 cells. TM-25659 mediated inhibition of PA-induced upregulation of TNF-α, IL-1β, IL-6, and MCP-1 in HepG2 cells and was significantly induced by compound C, an AMPK inhibitor (Fig. 11). These data suggest TM-25659 prevented PA-induced inflammation via AMPK signaling in HepG2 cells.
Fig. 11. TM-25659 prevented PA-induced upregulation of pro-inflammatory cytokine expression in HepG2 cells.HepG2 cells were harvested of PA, TM-25659, Compound C, or vehicle-treated cells for 8 h. TNF-α, IL-1β, IL-6, and MCP-1 mRNA expressions as quantified by real-time PCR. The data are given as means ± standard error. *P < 0.05, **P <
E. TM-25659 improved glucose homeostasis and insulin sensitivity in DIO mice
Glucose and insulin homeostasis decrease in obesity. The weight loss effects of TM-25659 were confirmed previously. Thus, experiments were carried out to verify if TM-25659 improves insulin resistance. The glucose levels of the HF group at fasting were 291 mg/dL, whereas that in the ND group was 196 mg/dL. The glucose levels in the HF+TM group decreased from 291 to 257 mg/dL at fasting (Fig. 12A). An OGTT showed TM-25659 repressed the elevation in glucose caused by oral glucose administration, demonstrating ameliorated glucose tolerance in the HF+TM group (Fig. 12B). The area under the curve (AUC) provided the same outcome, confirming TM-25659 repressed the AUC of the OGTT (Fig. 12C). The blood insulin levels in the HF group at fasting were 7.83 ng/mL, whereas that in the ND group was 1.72 ng/mL. The glucose levels in the HF+TM group decreased from 7.83 to 3.72 ng/mL at fasting (Fig. 12D). Mice administered TM-25659 orally showed reduced glucose profiles in reaction to an insulin injection compared to those in the HF group, indicating ameliorated insulin sensitivity in the HF+TM group (Fig. 12E). The AUC offered the same outcome, confirming TM-25659 repressed the AUC of the IPITT (Fig. 12F). These blood data suggest TM-25659 improved glucose metabolism and insulin sensitivity in DIO mice.
Fig. 12. TM-25659 improved glucose tolerance and insulin resistance in HF-diet induced obese mice. (A) Fasting blood glucose levels. (B) An oral glucose tolerance test (OGTT) was performed 8 weeks after TM-25659 treatment. (C) Area under the curve (AUC) of the OGTT. (D) Fasting blood insulin levels. (E) An intraperitoneal insulin tolerance test (IPITT) was performed 8 weeks after TM-25659 treatment. (F) AUC of the IPITT (ND – Normal chow diet, HF – High fat diet, HF+TM – High fat diet+TM-25659). The data shown are means ± standard error. *P < 0.05, ***P < 0.001 vs. vehicle in normal diet (ND) group
F. TM-25659 restored the insulin signaling pathways in DIO mice liver
In previous studies, we found the treatment of DIO mice with TM-25659 suppressed body weight gain and improved skeletal muscle insulin resistance (PART 1). So, we next examined whether TM-25659 mediated insulin resistance in DIO mice liver. DIO mice were administered TM-25659 to test its beneficial effects on HF-diet induced insulin signaling in the liver. The insulin-stimulated ND group had markedly increased levels of phosphorylated AKT in liver, whereas those in the HF group decreased markedly. The HF+TM group showed increased levels of insulin-stimulated AKT phosphorylation in liver, comparable to those in the HF group (Fig. 13). These results suggest TM-25659 prevented the HF-diet induced reduction in insulin signaling.
Fig. 13. TM-25659 prevented reduction of insulin-stimulated AKT phosphorylation in HF diet-fed mice liver. C57BL/6J mice were fed HF for 6 weeks, separated into two groups and then orally administrated every other day with either vehicle or TM-25659 (50 mg/kg) for 8 weeks. Representative immunoblots showing phosphor (Ser 473) and total AKT in mice liver of the ND, HF, and HF+TM group mice. (ND – Normal chow diet, HF – High-fat diet, HF+TM – High-fat diet+TM-25659) The data are given as means ± standard error. **P<0.01, ***P<0.001, †††P < 0.001; n=6/group.
G.
TM-25659 lowered the serum and liver pro-inflammatory cytokine levels in
DIO mice
The HF group increased serum pro-inflammatory cytokine levels on fasting. The serum fasting TNF-α levels in the ND group was 3.05 pg/mL, while that in the HF group was 5.21 pg/mL. The HF+TM group had significantly decreased levels of TNF-α, from 5.21 to 3.78 pg/mL at fasting (Fig. 14A). The serum IL-6 levels in the HF group at fasting was 4.51 pg/mL, whereas that in the ND group was 2.89 pg/mL. The HF+TM group showed a decreased (P = 0.06) IL-6 level from 4.51 to 3.37 pg/mL at fasting (Fig. 14B). The serum MCP-1 levels in the HF group at fasting was 54.76 pg/mL, whereas that in the ND group was 13.51 pg/mL. The HF+TM group had a significantly decreased MCP-1 level from 54.76 to 17.38 pg/mL at fasting (Fig. 14C). These data suggest TM-25659 prevented the HF-diet induced upregulation of serum pro-inflammatory cytokines in C57BL/6J mice. Increased liver inflammation levels are observed in diet induced obesity. The effects of TM-25659 in blood concentrations of alanine aminotransferase (ALT) and aspartate aminotransferase (AST) were also examined. As shown in Fig. 15, TM-25659 significantly lowered ALT and AST levels. Taken together, these results suggests TM-25659 treatment of HF-diet fed mice attenuates liver injury levels. We next examined whether TM-25659 protected against pro-inflammatory cytokine gene expressions in HF-diet fed mice. The HF group showed
Fig. 14. TM-25659 reduced inflammatory cytokine levels in HF diet-fed mice. (A) Serum levels of tumor necrosis factor (TNF)-α. (B) Serum levels of interleukin (IL)-6. (C) Serum levels of monocyte chemoattractant protein-1 (MCP-1). (ND – Normal chow diet, HF – High-fat diet, HF+TM – High-fat diet+TM-25659) The data are given as means ± standard error. *P < 0.05, **P < 0.01, ***P < 0.001 vs. vehicle in ND group mice. ††P < 0.01, †††P < 0.001 vs. vehicle in HF group mice; n = 8/group.
Fig. 15. TM-25659 prevented HF-diet induced liver injury.C57BL/6J mice were fed HF for 6 weeks, separated into two groups and then orally administrated every other day with either vehicle or TM-25659 (50 mg/kg) for 8 weeks. (A) Serum levels of ALT (B) Serum levels of AST. (ND – Normal chow diet, HF – High-fat diet, HF+TM – High-fat diet+TM-25659) The data shown are means ± standard error. ***P < 0.001 vs. vehicle in normal diet (ND) group mice. †P < 0.05, ††P < 0.01 vs. vehicle in HF group mice; n = 8/group.
Fig. 16. TM-25659 prevented HF-diet induced upregulation of pro-inflammatory cytokines in HF diet-fed mice liver. C57BL/6J mice were fed HF for 6 weeks, separated into two groups and then orally administrated every other day with either vehicle or TM-25659 (50 mg/kg) for 8 weeks. (A-D) TNF-α, IL-6, MCP-1, and IL-1β mRNA expressions in liver were measured by way of quantified by real-time PCR (ND – Normal chow diet, HF – High-fat diet, HF+TM – High-fat diet+TM-25659). The data shown are means ± standard error. **P < 0.01, ***P < 0.001 vs. vehicle in normal diet (ND) group mice. †P < 0.05, ††P <
H. TM-25659 activated AMPK pathways in DIO mice liver
In previous experiments, TM-25659 prevented the HF-diet induced upregulation of liver inflammation levels. AMPK is a key regulator of inflammatory responses that treatment with AMPK activators prevents palmitate-induced pro-inflammatory cytokines (Mihaylova and Shaw., 2011; Jäger, et al., 2007; Salminen, et al., 2011). So, we next examined whether TM-25659 mediated AMPK in HF-diet fed mice. The HF group showed decreased ACC and AMPK phosphorylation in the liver. The HF+TM group showed increased levels of phosphorylated ACC and AMPK in the liver, comparable to those in the HF group (Fig. 17). These data suggest TM-25659 prevented the HF-diet reduced ACC, AMPK phosphorylation in C57BL/6J mice liver.
Fig. 17.TM-25659 prevented reduction on AMPK phosphorylation in HF diet-fed mice liver. C57BL/6J mice were fed HF for 6 weeks, separated into two groups and then orally administrated every other day with either vehicle or TM-25659 (50 mg/kg) for 8 weeks. Representative immunoblots showing phosphor-ACC, total ACC, phosphor-AMPK, and total AMPK in mice liver of the ND, HF and HF+TM group mice (ND – Normal-chow diet, HF – High-fat diet, HF+TM – High fat diet+TM-25659). The data shown are means ±
I. TM-25659 improved serum lipid levels in HF-diet fed mice
The HF group showed increased fasting serum lipid levels. The serum fasting triglyceride levels in the ND group was 98 mg/dL, while that in the HF group was 123 mg/dL. The HF+TM group had significantly decreased levels of triglyceride from 123 to 88 mg/dL at fasting (Fig. 18A). The serum total cholesterol levels in the HF group at fasting was 244 mg/dL, whereas that in the ND group was 141 mg/dL. The HF+TM group showed a decreased total cholesterol level from 244 to 206 mg/dL at fasting (Fig. 18B). The serum free fatty acid levels in the HF group at fasting was 1511 uEq/L, whereas that in the ND group was 1223 uEq/L. The HF+TM group had a significantly decreased free fatty acid level from 1511 to 1172 uEq/L at fasting (Fig. 18C). These data suggest TM-25659 prevented the HF-diet induced upregulation of serum lipid levels in C57BL/6J mice.
Fig. 18. TM-25659 improved the serum lipid profiles in HF diet-fed mice. C57BL/6J mice were fed HF for 6 weeks, separated into two groups and then orally administrated every other day with either vehicle or TM-25659 (50 mg/kg) for 8 weeks. (A) Serum levels of triglyceride. (B) Serum levels of total cholesterol. (C) Serum levels of free fatty acid. (ND – Normal chow diet, HF – High-fat diet, HF+TM – High-fat diet+TM-25659) The data shown are means ± standard error. **P < 0.01, ***P < 0.001 vs. vehicle in ND group mice. ††P < 0.01, †††P < 0.001 vs. vehicle in HF group mice; n = 8/group.
J. TM-25659 prevented HF-diet induced hepatic steatosis
An increase in hepatic steatosis is also generally observed in diet-induced obesity (Perlemuter et al., 2007). Results in Fig. 11 show HF-diet induced significant lipid accumulation in the liver, as evidenced by the large cell vacuolation in H&E stained mice liver sections and Oil Red O-stained large lipid droplets. However, TM-25659-treated mice displayed significantly less fat accumulation in the liver, to levels similar to mice fed Normal chow diet (Fig. 19).
Fig. 19. TM-25659 ameliorated hepatic steatosis in HF diet-fed mice. C57BL/6J mice were fed HF for 6 weeks, separated into two groups and then orally administrated every other day with either vehicle or TM-25659 (50 mg/kg) for 8 weeks. Images of liver sections
K. TM-25659 reduced expression of lipogenic genes but increased expression of fatty acid oxidation genes in HF diet-fed mice.
We next examined whether TM-25659 protected against lipogenesis gene expressions in HF-diet fed mice. The HF group showed increased levels of PPAR-γ, Mogat1, and VLDL receptor mRNA expressions in the liver. The HF+TM group showed decreased levels of PPAR-γ, Mogat1, and VLDL receptor mRNA expressions in the liver, comparable to those in the HF group (Fig. 20A). Also, The HF group showed decreased levels of Acadl, Mcad, PPAR-β/δ, and UCP2 mRNA expressions in the liver. The HF+TM group showed increased levels of Acadl, Mcad, PPAR-β/δ, and UCP2 mRNA expressions in the liver, comparable to those in the HF group (Fig. 20B). These data suggest TM-25659 improved the HF-diet induced upregulation of lipogenesis and reduction of fatty acid oxidation in C57BL/6J mice liver.