의
의
의학
학
학 석
석
석사
사학
사
학
학위
위
위 논
논
논문
문
문
Shh induces Motor Neuron
differentiation in P19 Embryonic
Carcinoma Cells
아
아
아 주
주
주 대
대
대 학
학
학 교
교
교 대
대
대 학
학
학 원
원
원
의
의
의 학
학
학 과
과
과
박
박
박 래
래
래 희
희
희
Shh induces Motor Neuron Differentiation in
P19 Embryonic Carcinoma Cells
by
Rae Hee Park
A Dissertation Submitted to The Graduate School of Ajou University
in Partial Fulfillment of the Requirements for the Degree of
MASTER OF MEDICAL SCIENCES
Supervised by
Haeyoung Suh-Kim, Ph.D.
Department of Medical Sciences
The Graduate School, Ajou University
박
박
박래
래
래희
희
희의
의
의 의
의
의학
학
학 석
석
석사
사
사학
학
학위
위
위 논
논
논문
문
문을
을 인
을
인
인준
준
준함
함
함.
.
.
심
심
심사
사
사위
위
위원
원
원장
장
장
서
서
서 해
해
해 영
영
영
인
인
인
심
심
심 사
사
사 위
위
위 원
원
원
이
이
이 영
영
영 돈
돈
돈
인
인
인
심
심
심 사
사
사 위
위
위 원
원
원
조
조
조 은
은
은 혜
혜
혜
인
인
인
아
아
아 주
주
주 대
대
대 학
학
학 교
교
교 대
대
대 학
학
학 원
원
원
2
2
20
0
00
0
05
5
5년
년
년 6
6
6월
월
월 2
2
22
2
2일
일
일
- ABSRACT-
Sonic hedgehog induces Motor Neuron Differentiation in P19
Embryonic Carcinoma Cells
Sonic hedgehog (Shh) is a member of the hedgehog family of signalling molecules and secreted from two signalling centers, the notochord and the floor
plate, where it functions as a morphogen to induce early dorso-ventral patterning of the cenral nervous system (CNS). More recently, multiple actions of Shh
during CNS development have been discovered in additional sites, where it specifies the fates of oligodendrocytes and motor neurons as well as
proliferation of neural precursors and control of axon outgrowth.
To explore the roles of Shh in neuronal differentiation, we utilized
embryonal carcinomal P19 cells as an in vitro model system. We overexpressed Shh in P19 cells and investigated it’s effects on proliferation and differentiation of
P19 cells., P19/Shh, P19 cells overexpressing Shh, proliferated at higher rates than normal P19 cells even when normal P19 cells stopped growth and underwent
differentiation. Ironically, P19/Shh also differentiated into neurons at higher rates than normal P19 cells. Upregulation of both proliferation and neuronal
differentiation P19/Shh suggests that Shh may induce neuronal fates from uncommitted P19 cells and concomitantly promote the proliferation of the
ChAT) as well as a dopaminergic neuron marker (TH) were upregulated in neuronal cells derived from P19/Shh. Collectively, these results suggest that Shh
may have multiple functions in neural induction of uncommitted stem cells, proliferation of neural precursors, and dorso-ventral patterning of the CNS.
Key words : Sonic hedgehog, Proliferation, Differentiation, Motor neuron, P19 cells
TABLE OF CONTENTS
ABSTACT --- i
TABLE OF CONTENTS --- ii
LIST OF FIGURES --- vi
LIST OF TABLE --- viii
LIST OF ABBREVIATIONS --- ix
I. INTRODUCTION --- 1
A. Sonic hedgehog --- 1
B. Transcriptional regulation of development by Shh --- 4
1. Midbrain dopaminergic neuron --- 4
2. Spinal cord motor neuron --- 5
C. P19 embryonic carcinoma cells --- 6
II. MATERIALS AND METHODS --- 8
A. Materials --- 8
B. Mathods --- 9
1. P19 cell culture --- 9
2. Purification & titeration of 5E1 antibody --- 9
2.1 Affinity chromatography --- 9
2.2 Enzyme-Linked Immunosorbent (ELISA) Assay --- 10
3. Preparation of Shh conditioned medium --- 10
5. Growth kinetics --- 11
6. 3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyltetrazolium bromide (MTT) assay --- 12
7. Differentiation of P19 cell --- 12
7.1 Conventional neuronal differentiation --- 12
7.2 Current neuronal differentiation --- 13
8. Reverse Transriptase-Polymerase Chain Reaction (RT-PCR) --- 14
9. Western blotting --- 15
10. Immunocytochemistry --- 16
III. RESULTS --- 17
1. Expression of hShhN in P19 cells --- 17
2. Preparation of 5E1, a neutralizing antibody against Shh --- 19
3. The concentration of secreted Shh --- 19
4. Shh induces proliferation of P19 cells --- 22
5. Shh induces neuronal differentiation of P19 cells --- 25
6. Shh promotes neuronal precursor cell/ motor neurons survival or proliferation --- 28
7. Shh induces motor neuron differentiation in P19 cells --- 30
8. Shh induces dopaminergic and motor neuron differentiation in P19 cells --- 34
9. Shh promotes neuronal differentiation in P19 cells without treatment of RA or aggregation culture --- 37
IV. DISCUSSION --- 39 V. CONCLUSIONS --- 43 REFERENCES --- 44 국문요약 국문요약국문요약 국문요약 --- 54
LIST OF FIGURES
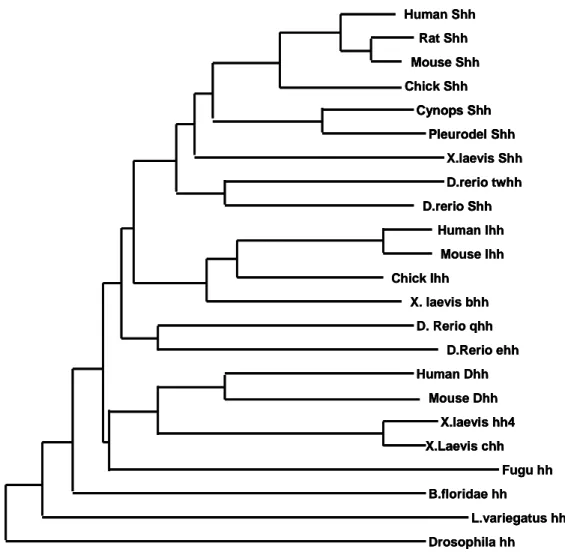
Fig. 1. Phylogentic relationship of numbers of the Hh protein family --- 2
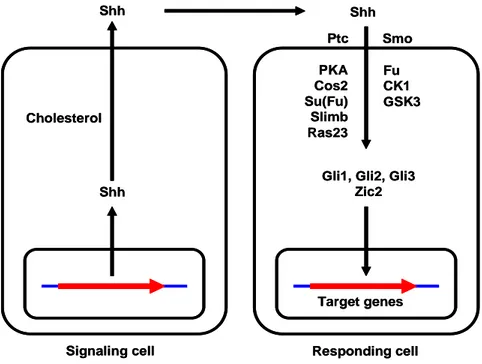
Fig. 2. The Shh signaling pathway --- 3
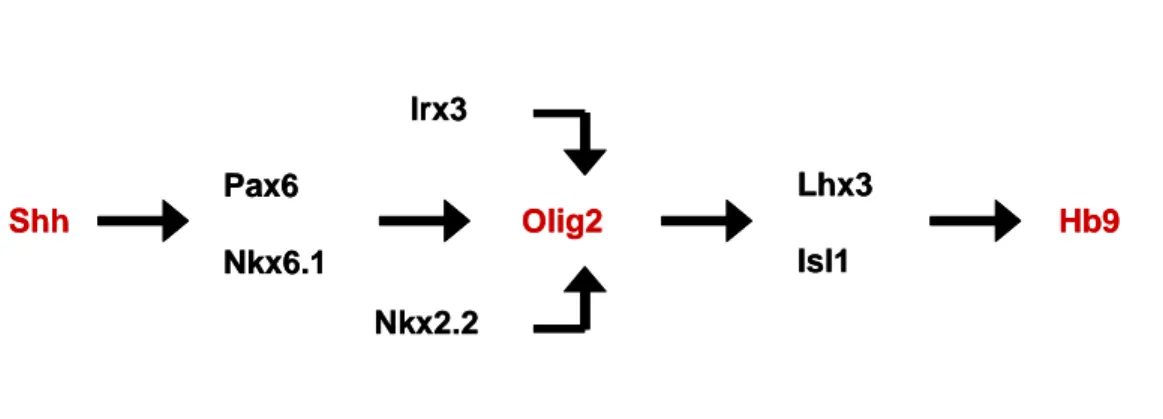
Fig. 3. Shh-activated transcriptional pathway of spinal motor neuron generation--- 6
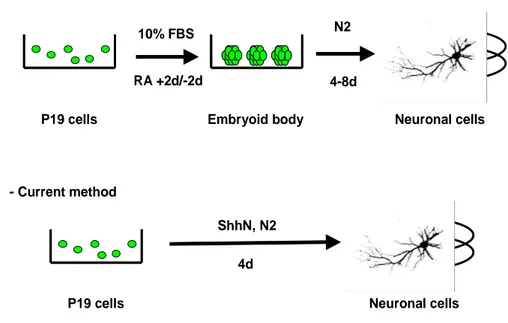
Fig. 4. The method for neural induction of P19 cells --- 13
Fig. 5. Generation of ShhN-Producing P19 Cell Lines --- 18
Fig. 6. Preparation of 5E1, a neutralizing antibody against Shh --- 20
Fig. 7. Soluble Shh activity is inhibited by Shh neutralizing antibody, 5E1 --- 21
Fig. 8. Shh induces proliferation of P19 cells --- 23
Fig. 9. Cell viability of P19, P19/hShhN cells was measured by MTT (3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyltetrazolium bromide) assay --- 24
Fig. 10. Neuronal differentiation of P19, P19/hShhN cells --- 27
Fig. 11. eGFP-labeled neuronal precursors in P19/pNPEeGFP --- 29
Fig. 12. RT-PCR of the expression of HD, bHLH and motor neuron specific markers, Nkx6.1, Olig2, Ngn2, NeuroD, Hb9 and ChAT in P19 and P19/hShhN --- 31
Fig. 13. Shh expression induces motor neuron differentiation of P19 cells (1) -- 32
Fig. 14. Shh expression induces motor neuron differentiation of P19 cells (2) -- 33
--- 35 Fig. 16. Shh expression induces dopamine neurons and motor neurons
differentiation of P19 cells --- 36 Fig. 17. Shh promotes neuronal differentiation in P19 cells without treatment
LIST OF TABLES
LIST OF ABBREVIATION
bHLH b helix-loop-helix
bp base pair
ChAT Choline acetyltransferase
CNPase 2', 3'-cyclic nucleotide 3'-phosphodiesterase
CNS Central nerve system DA Dopaminergic neuron
Dhh Desert hedgehog
DMEM Dulbecco's modified Eagle's medium
ECL Enhanced chemiluminescence
ELISA Enzyme-Linked Immunosorbent Assay
FBS Fetal bovine serum
FGF8 Fibroblast growth factor 8
FITC Fluorescein isothiocyanate Fsk Forskolin
GFAP Glial fibrillary acidic protein GFP Green fluorescent protein
HAT Hypoxanthine-Aminopterin Thymidine Hb9 Homeobox gene 9
HBSS Hank's balanced salt solution
Hh Hedgehog
HRP Horseradish peroxidase
IgG Immuonglobulin G Ihh Indian hedgehog
MN Motor neuron
NCAM Neuronal cell adhesion molecule
NF-M Neurofilament-160 kDa Ngn2 Neurogenin 2
OD Optical density
Olig2 Oligodendrocyte transcription factor 2
OPD o-phenylenediamine dihydrochloride P19 cell Mouse embryonic carcinoma cell
PAGE Polyacrylamide gel electrophoresis PBS Phosphate buffered saline
pMN Motor neuron presursor
Ptc Patched
RA Retinoic acid
RIPA Radioimmunoprecipitation
RT-PCR Revese transcriptase - polymerase chain reaction SDS Sodium dodecyl sulfate
Shh Sonic hedgehog
TBS Tris buffered saline TH Tyrosine hydroxylase
I. INTRODUCTION
A. Shh (Sonic hedgehog)
In vertebrates, three members of the Hedgehog (hh) gene family have been identified : Desert hedgehog (Dhh), Indian hedgehog (Ihh), and Sonic hedgehog
(Shh). The most-studied member is Shh, which is initially expressed in the notochord and then in the floor plate (Echelard et al. 1999; Chiang et al. 1996; for a review, see
Ingham and McMahon 2001). The signaling properties of the notochord and floor plate are associated with several developmental processes, among them the induction
or self-induction of the floor plate, the specification of neuronal identity, and the induction of sclerotome development in ventral somite.
Sonic Hedgehog (Shh) is a potent morphogen involved in the development of limbs and muscles as well as the anterior/posterior organization of the neural tube in
vertebrates (Goodrich and Scott, 1998). The 19 kDa amino terminal fragment of Shh (also called Shh-N) is released by autocatalytic proteolysis and induces graded
signaling of neighbor target cells (Marti et al., 1995; Briscoe and Ericson, 1999; McMahon, 2000). Covalent linkage of cholesterol to the C terminus of Shh may
result in tethering of Shh to the cell membrane, which plays an important role in long range signaling during patterning of chick limb and mouse digits (Zeng et al., 2001;
Lewis et al., 2001). On target cells, Shh binds Patched (Ptc) a 12 transmembrane which causes releases of Ptc-inhibition of Smoothened. (Smo), Smo is a 7
transmembrane domain G protein coupled receptor and triggers a signaling cascade that finally leads to expression of Shh target genes in the nucleus (Goodrich and
Scott, 1998; McMahon, 2000). Human Shh Rat Shh Mouse Shh Chick Shh X.laevis Shh D.rerio twhh D.rerio Shh Human Ihh Mouse Ihh Chick Ihh X. laevis bhh D. Rerio qhh D.Rerio ehh Human Dhh Mouse Dhh X.laevis hh4 X.Laevis chh Fugu hh B.floridae hh L.variegatus hh Drosophila hh Cynops Shh Pleurodel Shh Human Shh Rat Shh Mouse Shh Chick Shh X.laevis Shh D.rerio twhh D.rerio Shh Human Ihh Mouse Ihh Chick Ihh X. laevis bhh D. Rerio qhh D.Rerio ehh Human Dhh Mouse Dhh X.laevis hh4 X.Laevis chh Fugu hh B.floridae hh L.variegatus hh Drosophila hh Cynops Shh Pleurodel Shh Human Shh Rat Shh Mouse Shh Chick Shh X.laevis Shh D.rerio twhh D.rerio Shh D.rerio twhh D.rerio Shh Human Ihh Mouse Ihh Chick Ihh X. laevis bhh D. Rerio qhh D.Rerio ehh Human Dhh Mouse Dhh X.laevis hh4 X.Laevis chh X.laevis hh4 X.Laevis chh Fugu hh B.floridae hh L.variegatus hh Drosophila hh Cynops Shh Pleurodel Shh Cynops Shh Pleurodel Shh
Target genes Signaling cell Responding cell
Shh Cholesterol Shh Shh Ptc Smo PKA Cos2 Su(Fu) Slimb Ras23 Fu CK1 GSK3
Gli1, Gli2, Gli3 Zic2
Target genes Signaling cell Responding cell
Shh Cholesterol Shh Shh Ptc Smo PKA Cos2 Su(Fu) Slimb Ras23 Fu CK1 GSK3
Gli1, Gli2, Gli3 Zic2
Fig. 2. The Shh signaling pathway. Patched (Ptc), Smoothened (Smo), Protein kinase A (PKA), Costal-2 (Cos-2), Suppressor of fused (Su(Fu)), Fused (Fu), Casein
B. Transcriptional regulation of development by Shh
The embryonic expression pattern of Shh has been shown to be closely linked to
the development and differentiation of the entire ventral neuraxis (Marti et al., 1995a). Using naive neural tube explants derived from the appropriate levels of the
rostro-caudal axis, it has been demonstrated that the induction of spinal motor neurons (Roelink et al., 1994; Tanabe et al., 1995), midbrain dopaminergic neurons
(Hynes et al., 1995; Wang et al., 1995), and basal forebrain cholinergic neurons (Ericson et al., 1995) are dependent on exposure to Shh. This molecule appears to be
crucial for such patterning and phenotype specification in vivo, because mouse embryos deficient in the expression of functional Shh gene product manifest a lack of
normal ventral patterning in the CNS as well as gross atrophy of the entire cranium (Chiang et al., 1996).
1. Midbrain dopaminergic neuron
Midbrain dopaminergic neurons are the main source of dopamine in the mammalian central nervous system and are associated with one of the most
prominent human neurological disorders, Parkinson’s disease. During development, they are induced in the ventral midbrain by an interaction between two diffusible
factors, Shh and FGF8. The local identity of this part of the midbrain is probably determined by the combinatorial expression of three transcription factors, Otx2, Pax2,
and Pax5. After the last cell division, the neurons start to express transcription factors that control further differentiation and the manifestation of cellular properties
characteristic for adult dopaminergic neurons of the substantia nigra compacta and the ventral tegmentum. The first to appear is the LIM-homeodomain transcription
factor, Lmx1b. It is essential for the survival of these neurons, and it regulates the expression of another transcription factor, Pitx3, an activator of tyrosine hydroxylase
(TH). Lmx1b is followed by the orphan steroid receptor Nurr1. It is essential for the expression of dopaminergic phenotype. A day after Nurr1, two homeodomain
transcription factors, engrailed-1 and -2, are expressed.
2. Spinal cord motor neuron
In the embryonic nervous system, many different subtypes of neurons are
generated with distinct cellular and physiological properties. Neurons for controlling locomotion, located within the ventral spinal cord, are generated through the
coordinate actions of Shh and delta, which influence the subtype identity, timing of differentiation, numbers, and relative position of the cells (Bertrand et al., 2002;
Briscoe and Ericson, 2001; Jessell, 2000; Tanabe and Jessell, 1996). The dividing progenitor cells for motor neurons express a unique combination of transcription
factors in response to Shh, including Pax6, Nkx6.1, Olig2, and Mnr2 (Briscoe et al., 2000; Lee and Pfaff, 2001). These proteins have several roles in motor neuron
development: repressing transcriptional programs for other cell fates (Briscoe et al., 2000), activating neurogenesis via proneural bHLH factors (Mizuguchi et al., 2001;
Novitch et al., 2001; Scardigli et al., 2001), and promoting expression of the LIM homeodomain (LIM-HD) proteins Isl1 and Lhx3, required for motor neuron
specification (Thaler et al.,2002). Thus, single factors such as Olig2 coordinately switch on both bHLH and HD proteins (Mizuguchi et al., 2001; Novitch et al., 2001).
Shh Pax6 Nkx6.1 Olig2 Lhx3 Isl1 Hb9 Nkx2.2 Irx3 Shh Pax6 Nkx6.1 Olig2 Lhx3 Isl1 Hb9 Nkx2.2 Irx3
Fig. 3. Shh-activated transcriptional pathway of spinal motor neuron generation.
C. P19 embryonic carcinoma cell
Very little is known about the determination events that commit unspecialized
cells to differentiate into the more specialized cell types that appear later during embryonic development. Mouse embryonic carcinoma cells, the pluripotent stem
cells of malignant teratocarcinomas, may provide a culture system with which to study these events. Lines of embryonic carcinoma cells can be isolated from
teratocarcinomas and maintained in an undifferentiated state when kept in exponential growth phase in tissue culture. If left undisturbed at high density, they
differentiate in vitro into a variety of cell types including epithelium, neurons, muscle, and cartilage.
The P19 mouse embryonic carcinoma cell line is an established model of cell differentiation. Developmentally, pluripotent P19 cells give rise to the formation of
cell derivatives of all three germ layers and appear to differentiate by the same mechanisms as normal embryonic stem cells. When cultured in the presence of 10-6 M retinoic acid, a physiologically relevant morphogen, P19 cells differentiate into neuroectodermal cell lineage, such as neurons and glial cells neurons. However, we
showed the overexpression of Shh could direct P19 cells to differentiate into neuronal cells in the absence of RA possibly through the Shh signaling pathway.
In this report, we show that P19/hShhN cells, when replated as the single cells suspension, could differentiate only into neurons in the absence of RA.
II. MATERIRALS AND METHODS
A. MATERIRALS
FITC conjugated secondary antibodies from Vector Laboratory (Burlingame, CA, U.S.A); βIII-tubulin (Tuj-1) specific antibody from Berkeley antibody company
(Richmond, CA, U.S.A); neurofilament-160kDa (NF-M) specific antibody from Zymed Laboratory (South San Francisco, CA, U.S.A); 2', cyclic nucleotide
3'-phosphodiesterase (CNPase) specific antibody, Choline acetyltransferase (ChAT) and Hb9 specific antibody from Chemicon (Temecula, CA, U.S.A); DMEM and
N2-supplement from GibcoBRL (Grand Island NY, U.S.A); FBS from Hyclone Inc. (Logan, UT, U.S.A); 5E1 hybrydoma cell from DSHB (IA, U.S.A); Protein G
sepharoseTM 4 Fast Flow from Amersham Bioscience (Buckinghamshire, U.K); Mouse IgG , glial fibrillary acidic protein (GFAP) specific antibody, neuronal cell
adhesion molecule (NCAM) specific antibody, All-trans-retinoic acid(RA) , Tyrosine Hydroxylase (TH) specific antibody, 2-mercaptoethanol, o-phenylenediamine
dihydrochloride (OPD), Forskolin, Tris, Glycine, sodium dodecyl sulfate(SDS), polyacrylamide, bis-acrylamide, and TEMED from Sigma (St. Louis, MO, U.S.A);
Enhanced chemiluminescence (ECL) kit from Pierce (Rockford, IL, U.S.A); Westran PVDF membranes from Schleicher & Schuell (Dassel, Germany); RNAzolTM B
regents from TEL-TEST Inc. (Frendwood, TX, U.S.A); Frist strand cDNA synthesis kit from Roche (Indianapolis, U.S.A); Super Taq DNA polymerase from Korea B&G
(Suwon, Korea).
All other chemicals were obtained from Sigma-Aldrich (St. Louis, MO, U.S.A).
B. METHODS
1. P19 cell culture
P19 cells were maintained in Dulbecco’s modified Eagle’s medium (DMEM) with 10% fetal bovine serum (FBS), 100unit/ml penicillin, 100㎍/ml streptomycin in
a standard humidified atmosphere at 37℃. After the cultures reached 80-90% confluence, the cells were harvested with 0.25% trypsin and 1mM EDTA for 2-3min
at 37℃, re-plated after 1:10 dilution on 100mm culture dish.
2. Purification & Titration of 5E1 antibody 2.1 Affinity chromatography
Ascitic fluid containing an anti-Shh monoclonal antibody, 5E1 was mixed with on Protein G sepharose 4 Fast Flow beads. prebound gel matrix was packed in a
glass column of 1x10cm. The column was equilibrated with binding buffer (20mM sodium phosphate buffer pH 7.0). Then 10ml of binding buffer was applied to the
column at a flow rate of 0.4ml/min. The 5E1 antibody was eluted with 20ml of 0.1M Glycine-HCl pH 2.8 at a flow rate of 0.8/min. Each fraction was monitored by
Bradford assay and the peak fractions were neutralized with 0.05ml of 1M Tris-HCI pH 9.
2.2 ELISA (Enzyme-Linked Immunosorbent Assay)
Screening for the presence of an anti-Shh mAb (5E1) in the purified 5E1 and
Ascitic fluid was performed with lab-made ELISA. High binding capacity 96-well flatbottomed microtiter plates were coated with Proleukin. To each well was added
100µl of culture supernatants at 100µg/ml in 0.1% Tween-20 containing PBS (PBS-T) and then they were incubated overnight at 4℃. After washing five times with
PBS-T, potential binding sites were blocked with 200µl of 5% bovine serum albumin in PBS-T and incubated for 1hr at 4℃. The purified 5E1(1:10000) and Asctic fluid
(1:1000, 1:5000, 1:100 and 1:10), 100µl per well, were incubated overnight at 4℃. After washing five times with 0.1% Tween-20 containing PBS (PBS-T), to each well
was added 100µl of horse radish peroxidase (HRP)-conjugated goat anti-mouse IgG, diluted to 1:5000 in PBS-T, and then incubated for 2 hr at 37℃. After washing five
times, to each well was added 100µl of OPD (o-phenylenediamine dihydrochloride) substrate and then they were incubated for 10 minutes at room temperature. The
reaction was stopped by adding 50µl of 2.5M H2SO4 per well and then optical
density (OD) was evaluated with an ELISA reader using either a 490nm filter.
3. Preparation of ShhN-Conditioned medium
After P19 and P19/hShhN cells had been plated in tissue culture dish at a density of 1 x 105 cells/ml in 10% fetal bovine serum (FBS)/DMEM (Gibco) for 1
day, culture medium was switched to serum-free medium supplemented with N2 supplement and to fresh 10% fetal bovine serum (FBS)/DMEM. Two days later, the
medium was collected as P19 (PCM) and P19/hShhN (SCM)-conditioned medium. The medium filtered through a 0.2 ㎛ pore-size filter. The PCM and SCM is stored
at 4℃ for use.
4. Concentration of secreted ShhN
The concentration of Shh in cultured P19/hShh and P19/hShhN conditioned
media was determined using a Shh ELISA kit protocol (R&D). Using known concentrations of Shh (0-1000 pg/ml) a standard curve was calculated during each
assay using the same 96-well, flat bottom, high binding, EIA/RIA. The concentration of Shh in the conditioned media fell with linear range of the standard curve and thus
the Shh concentration in the media was determined from standard curve by interpolation.
5. Growth kinetics
P19, P19/hShhN cells were plated at a density of 5 x 103 cells per well of 96-well plate. Following overnight attachment, cells were maintained for 4 days with
culture media, serum free media and conditioned media. Anti-Shh, 5E1 and control mIgG added into the medium to a final concentration of 10 ㎍/ml for the 4 days.
Conditioned media was collected 48 hr during P19 and P19/hShhN cells culture and filtered through a 0.2㎛ filter. Cell proliferation was measured in culture using
Trypan-Blue exclusion method. Briefly, the cells were harvested with trypsin, and mixed with 0.4% Trypan-Blue solution. After 1min incubation, living cells were
counted.
6. MTT [3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyltetrazolium bromide]assay The proliferation activity of cells was assessed using the MTT assay. This
method is based on the ability of mitochondrial dehydrogenases in living cells to reduce soluble tetrazolium salts to a blue formazan product whose amount is directly
proportional to the number of living cells. P19, P19/hShhN cells in suspension (500µl) were added to each well of a 4-well culture plate for a final concentration of
5x103 cells/well with 5E1 (10µg/ml) or IgG (10µg/ml) and incubated for 24h, 48h, 72h and 96h at 37°C in a CO2 incubator. At appropriate time points, 300µl of 0.5
mg/ml MTT solution were added to each well, and the cultures further incubated for 2h. Subsequently, the medium with MTT was replaced with 0.2 ml dimethyl
sulphoxide. Sample (100µl) were added to each well of a 96-well culture plate. A microplate reader was used to measure absorbance at 570 nm for each well.
7. Differentiation of P19 cells
7.1 Conventional neuronal differentiation
P19 cells were allowed to aggregate in bacterial Petri dishes at a seeding
density of 1 x 105 cells/ml in the presence of 0.5 µM all-trans-retinoic acid in 10% fetal bovine serum (FBS)/DMEM. After 4 days of aggregation, cells were dissociated
into single cells by 0.05 % trypsin-EDTA , and were replated in tissue culture dish at a density of 1 x 105 cells/ml in a N2 serum-free medium [DMEM/F12 supplemented
with 5 ㎍/ml insulin, 50 ㎍/ml human transferring, 20 mM progesterone, 60 µM putresine, and 30 nM sodium selenite] supplemented with or without final
concentration 10 µM forskolin. The cells were then allowed to adhere and cultured for 4 days with replacement of the medium every 48h.
7.2 Current neuronal differentiation
P19 cells were plated in tissue culture dish at a density of 2 x 105 cells/ml in 10% fetal bovine serum (FBS)/DMEM. To induce neuronal differentiation, the media
were removed, and the cells were washed with HBSS and was replaced with N2 serum-free medium with or without final concentration 10 µM forskolin. The cells
maintained for 4 days. The same protocol was used for P19/hShhN cells.
- Conventional method
- Current method
Neuronal cells P19 cells Embryoid body
10% FBS N2 4-8d
ShhN, N2
P19 cells Neuronal cells 4d
RA +2d/-2d
8. Reverse Transcriptase Polymerase Chain Reaction (RT-PCR)
Total RNA was isolated using a RNAzolTM B and cDNA was synthesized
using First-strand cDNA synthesis kit and 1㎍ of total RNA following the manufacture’s instructions. The PCR reactions were carried out according to standard
protocols. Primer sequences (forward and reverse) and product sizes (base pairs) were as follows (Table 1). The PCR products were analyzed on a 1.5% agarose gel.
Table 1. Primer sequence for RT-PCR
Forward Reverse Size (bp)
TCCATGACAACTTTGGCATCGTGG
GAPDH GTTGCTGTTGAAGTCACAGGAGAC 395
Smo GGAACTCSAATCGCTACCC GCTGGCWGCCTTCTCACT 560 Ptc ACCTTTGGACTGCTTCTGGG GAGTCATTAACTGGAACATGG 416 Gli ATGCTGTCTCCGGCCCCTCC TCCTCCCGCCCATCCAGCATC 620 Nestin GCGGGGCGGTGCGTGACTAC CAAGAGAAGCCTGGGAACCT 395 NF-M GCCGCTACGCCAAGCTCACCG CTGTCGGTGTGTGTACAGAGG 423 TH GTTCTCAACCTGCTCTTCTCC GCATAGAGGCCCTTCAGCGTG 490 Nurr1 TTAAAAGGCCGGAGAGGTCG CTCTCTTGGGTTCCTTGAGCC 557 Ptx3 GCGCACGCACTTCACCAGCC GCCAGGCTCGAGTTACACGG 600 Olig2 CAGCGAGCACCTCAAATCTA CACCAGTCGCTTCATCTGCT 601 Nkx6.1 CACCCCACACGGCATCAAA GGGCGAAGATTTGCTGTCC 476 Ngn2 CACGCACGAGAACGACAACA CGGTTGTTGGCCTTGAGC 579 NeuroD CTCCGGGGTTATGAGATCGTCAC GCTCTAGATCTCTGACAGAGCCCAAATG 390 Hb9 ACAGGCGGCTCTCTATGGACAC TTCCCCAAGAGGTTCGACTGC 258 ChAT GGCCATTGTGAAGCGGTTTG TGACATGCTCGGGCTCAGGC 425
9. Western blotting
Whole cell lysates were prepared as follows; Cells were washed with cold
phosphate buffered saline (PBS) twice and drained. Cells were resuspended in RIPA buffer [150mM Sodium chloride, 1% NP-40, 0.1% SDS, 50mM Tris-HCl (pH8.0),
0.5% sodium deoxycholate] and kept on ice for 5 min. After centrifugation at 12000 rpm for 20min, the supernatant was collected and protein content was assayed by
Bradford method. Each 60 ㎍ proteins were separated on 8% or 12% SDS polyacrylamide gel electrophoresis and transferred to PVDF membranes. Membranes
were incubated in blocking solution [5% nonfat dry milk in 10mM Tris-HCl (ph7.4), 100mM NaCl, and 0.1% Tween 20] for 1 hour at room temperature and with mouse
anti- ShhN (1:100), mouse anti-βIII -tubulin (Tuj-1, 1:1,000), mouse anti-neuronal cell adhesion molecule (NCAM, 1:1,000), mouse anti-neurofilament-160kD (NF-M,
1:1,000), mouse anti-glial fibrillary acidic protein (GFAP, 1:1,000), and mouse anti-2', 3'-cyclic nucleotide 3'-phosphodiesterase (CNPase, 1:1,000) antibody in
TTBS [10mM Tris-HCl (ph7.4), 100mM NaCl, and 0.1% Tween 20] solution for overnight at 4°C. The secondary antibody was horseradish-peroxidase
(HRP)-conjugated to goat anti-mouse IgG (1:5,000) or anti-goat IgG (1:5000) and the proteins were visualized using an enhanced chemiluminescence (ECL) kit following
10. Immunocytochemistry
Differentiated P19 and P19/hShhN cells were fixed with 2~4%
paraformaldehyde solution or 100% methanol for 10 min at 4℃. The fixed was washed with PBS solution, three times and incubated with 10% normal horse serum
and goat serum, 0.1% BSA and 0.03% Triton X-100 in PBS (PBS-T) for 1hour. After sufficient washing, the cells were incubated with primary antibodies overnight at 4℃.
The following primary antibodies were used at the concentrations given: Rat 401 (1:200), GFP (1:200), Ki-67 (1:200), Tuj1 (1:200), NFM (1:200), TH (1:200), Isl-1
(1:200), ChAT (1:200) and Hb9 (1:200). Following several washing with 0.03% Triton X-100 containing PBS-T solution, cells were incubated in FITC conjugated
anti-mouse IgG (1:500), anti-rabbit IgG (1:500), Alexa 488 anti-mouse IgG (1:500), or Alexa 594 anti-rabbit IgG (1:500) secondary antibody for 1 hour in dark chamber.
After sufficient washing with PBS-T, cells were mounted on acras or cover slips by using Vectashield (mounting medium for fluorescence, Vector) with DAPI or
III. RESULTS
1. Expression of hShhN in P19 cells
To determine whether Shh could play a role in neuronal differentiation, the expression of components of the Shh signaling pathway in P19 cells was analyzed in
western blot. Shh was expressed in P19 cells (Figure 5 A). The N-terminal fragment receives two lipid modifications and, after being secreted, can tether to the
membrane of producing cells or diffuse, establishing concentration gradients (Porter et al. 1996; Pepinsky et al. 1998; Lewis et al. 2001; Zeng et al. 2001). Medium
conditioned by P19/hShhN cells contained Shh-N (Figure 5 A). The components of the Shh receptor complex, Ptc and Smo, as well as the three Gli transcription factors
involved downstream of Shh signaling, have been demonstrated by RT-PCR to be expressed on P19 cells (Figure 5 B).
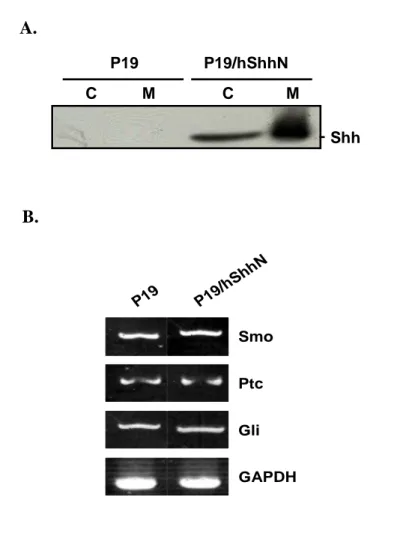
A. P19 P19/hShhN C M C M Shh B. Smo Ptc Gli GAPDH P19 P19/ hShh N
Fig. 5. Generation of ShhN-Producing P19 Cell Lines. (A) Cells, P19 and P19/hShhN, were harvested and media collected, separated by SDS-polyacrylamide gel electrophoresis (SDS-PAGE), transferred to PVDF membrane, and probed with
antibodies direct against the anti-ShhN. ShhN protein as a 20 kDa was detected both cell extracts and medium in P19/hShhN cell. (B) Expression level of Smo, Ptc and
2. Preparation of 5E1, a neutralizing antibody against Shh
5E1 hybridoma cells were obtained from DSHB (IA, U.S.A). The cells were
grown in the presence of HAT (Hypoxanthine-Aminopterin Thymidine). The cells were injected into the peritoneal cavity of BalbC female (6~8 week) to obtain ascitic
fluid. The titer of 5E1 ascitic fluid was determined by ELISA using P19/hShhN conditioned medium as an antigen. 5E1 reacted with secreted ShhN in ELISA at
dilutions up to >1:1000 (ascitic fluid) (Fig 6 A). IgG fractions were obtained by affinity purification on protein G–sepharose (Fig 6 B). Purified 5E1 were measured
the concentration of ShhN by coomassie blue staining and showed neutralizing activity with ShhN in growth kinetics. P19/hShhN were inhibited proliferation at
10~30 ㎍/ml 5E1 (Fig 7).
3. The Concentration of secreted Shh
Shh detected in media conditioned by P19/hShhN using an ELISA. As this
concentration of Shh is near the low limit of detection we used the same ELISA protocol to measure Shh in growth media containing 10% FBS, conditioned media
form P19/hShhN and differentiation media containing N2 supplement in DMEM:F12 from P19/hShh. The concentration of secreted Shh has 0.7ng/ml in differentiation
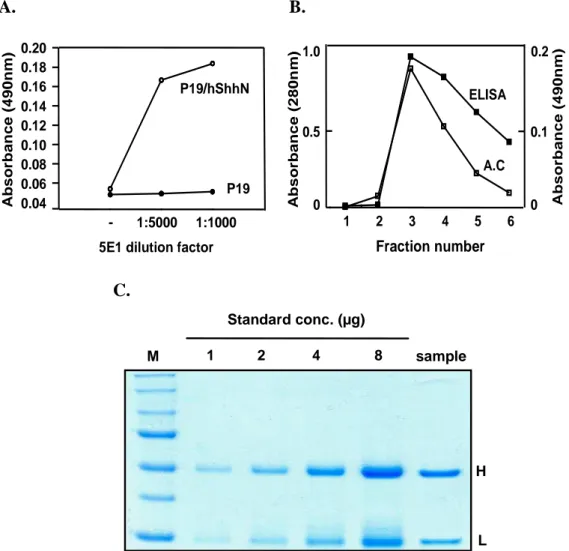
A. B. A b s o rb a n c e ( 4 9 0 n m ) - 1:5000 1:1000 0.04 0.06 0.08 0.10 0.12 0.14 0.16 0.18 0.20 P19 P19/hShhN
5E1 dilution factor
A b s o rb a n c e ( 2 8 0 n m ) ELISA A.C 0 0.5 1.0 1 2 3 4 5 6 Fraction number 0 0.1 0.2 A b s o rb a n c e ( 4 9 0 n m ) C. M 1 2 4 8 sample Standard conc. (µg) H L
Fig. 6. Preparation of 5E1, a neutralizing antibody against Shh. 5E1 reacted with secreted ShhN in ELISA at dilutions up to 1:1000~5:000 (ascitic fluid) (A). The titer of 5E1 was determined by ELISA using P19/hShhN conditioned medium as an
antigen and its purified by affinity chromatography (B). Purified 5E1 were measured the concentration of ShhN by coomassie blue staining (C).
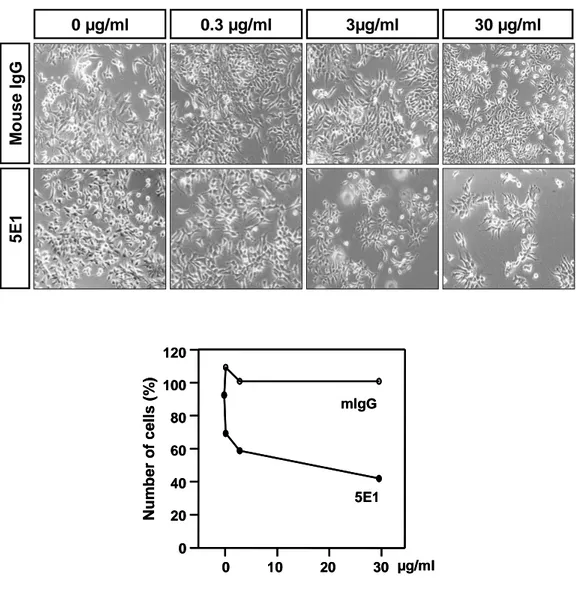
M o u s e I g G 5 E 1 0 µg/ml 0.3 µg/ml 3µg/ml 30 µg/ml N u m b e r o f c e ll s ( % ) 0 10 20 30 µg/ml 0 20 40 60 80 100 120 mIgG 5E1 N u m b e r o f c e ll s ( % ) 0 10 20 30 µg/ml 0 20 40 60 80 100 120 mIgG 5E1
Fig. 7. Soluble Shh activity is inhibited by Shh neutralizing antibody, 5E1. P19/hShhN cells were plated at density of 3 x 103 cells per well 48-well plate and
cultured with DMEM supplemented 0.5% FBS for 4 days. Cells incubated with dilution of 5E1 monoclonal antibody (mAb) or IgG and the total living cell number
4. Shh induces proliferation of P19 cells
To test whether Shh regulates P19 cell growth and differentiation, we cultured
cells for 96hr in the presence of 5E1, as Shh neutralizing antibody, or control IgG and analyzed the cultures by cell counting using Trypan-blue dye exclusion method.
These assays were performed in the presence or absence of specific Shh inhibitor, the Shh neutralizing monoclonal antibody 5E1. Purified 5E1 (10µ g/ml) was added to the
medium and the medium was replaced every 2 days. Cultures grown in P19 cells demonstrated low proliferation rate. However, a 2-fold, significant increase in cell
proliferation was observed in P19/hShhN. P19/hShhN cells were almost completely inhibited by 5E1 monoclonal antibody, but unaffected by mIgG (Figure 8 A). The
hShhN effect was blocked by 5E1. To test whether secreted Shh regulates proliferation of P19 cells, we cultured for 4 days in P19 cells conditioned medium
and P19/hShhN cells conditioned medium. Conditioned medium from P19/hShhN increased the proliferation of P19 cells, which was blocked by 5E1 (Fig 8 B). Unlike
P19 cells, the proliferating potential of P19/hshhN cells were maintained under the differentiation condition of P19 (Fig 8 C).
To confirm the effect of Shh on P19 cell proliferation, proliferation was studied by using 5E1 monoclonal antibody. Neutralizing activity of 5E1 monoclonal
antibody was studied by incubating the antibody with 5E1 or IgG. The MTT assay was then performed after 92 hours incubation. As shown in Figure 9, P19/hShhN
cells increased proliferation and inhibited by 5E1 (10 ㎍/ml) by compared to the control groups (Fig 9).
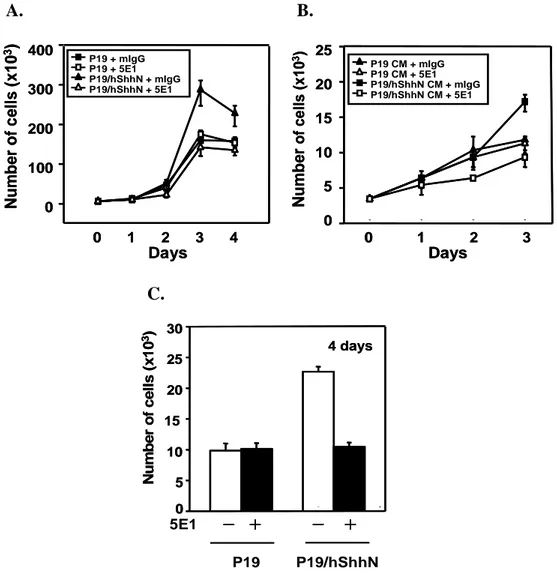
A. B. N u m b e r o f c e ll s ( x 1 0 3) 0 1 2 3 Days 4 0 100 200 300 400 P19 + mIgG P19 + 5E1 P19/hShhN + mIgG P19/hShhN + 5E1 0 1 2 3 Days N u m b e r o f c e ll s ( x 1 0 3) 0 5 10 15 20 25 P19 CM + mIgG P19 CM + 5E1 P19/hShhN CM + mIgG P19/hShhN CM + 5E1 N u m b e r o f c e ll s ( x 1 0 3) 0 1 2 3 Days 4 0 100 200 300 400 P19 + mIgG P19 + 5E1 P19/hShhN + mIgG P19/hShhN + 5E1 0 1 2 3 Days N u m b e r o f c e ll s ( x 1 0 3) 0 5 10 15 20 25 P19 CM + mIgG P19 CM + 5E1 P19/hShhN CM + mIgG P19/hShhN CM + 5E1 C. 5E1 P19 P19/hShhN 0 5 10 15 20 25 30 4 days N u m b e r o f c e ll s ( x 1 0 3) 0 5 10 15 20 25 30 4 days N u m b e r o f c e ll s ( x 1 0 3) ╋ ╋ ╋ ╋ ━ ━━ ━ ━━━━ ╋╋╋╋
Fig. 8. Shh induces proliferation of P19 cells. P19 andP19/hShhN cells were plated at a density of 3 x 103 cells per well of 48-well plate and cultured with DMEM
supplemented 10% FBS (A), 0.5% FBS (B) and N2 (C) for 4 days. Cells incubated with 10µg/ml of 5E1 monoclonal antibody (mAb) or IgG and the total living cell
number were measured by Trypan blue dye exclusion method. Data are expressed as a mean of duplicate per time point.
0 1 2 3 1 2
A
b
s
o
rb
a
n
c
e
(
5
7
0
n
m
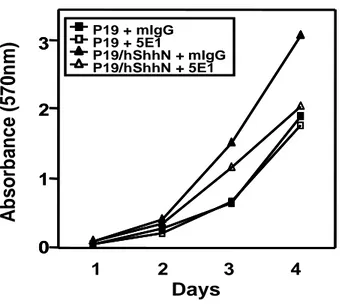
)
0 1 2 3 3 4 P19 + mIgG P19 + 5E1 P19/hShhN + mIgG P19/hShhN + 5E1 DaysFig. 9. Cell viability of P19, P19/hShhN cells was measured by MTT (3-(4,5 dimethylthiazol-2-yl)-2,5-diphenyltetrazolium bromide) assay. P19, P19/hShhN cells in suspension (500µl) were added to each well of a 4-well culture plate for a final concentration of 5x103 cells/well with 5E1 (10µg/ml) or IgG (10µg/ml) and
incubated for 24h, 48h, 72h and 96h at 37°C in a CO2 incubator. At appropriate time points, 300µl of 0.5 mg/ml MTT solution were added to each well, and the cultures
further incubated for 2h. Subsequently, the medium with MTT was replaced with 0.2 ml dimethyl sulphoxide. Sample (100µl) were added to each well of a
96-well culture plate. A microplate reader was used to measure absorbance at 570 nm for each well.
5. Shh induces neuronal differentiation of P19 cells
In vitro neuronal differentiation of P19 cells and P19/hShhN cells were grown
without or with 10µM forskolin in the presence of N2 supplement for 4 days. P19/hShhN cells formed neurite-like processes and showed a neuron-like
morphology on 4 days (Figure 10 A). RT-PCR analyzed for differentiation of P19 and P19/hShhN cells into neural stem/precursors (Nestin), neurons (NF-M) (Fig 10
B). Expression of a neuron-specific protein, NF-M and β-tubulin III were increased in P19/hShhN, which were further increased by addition of 10µM forskolin.
Expression of proteins specific for oligodendrocyte (CNPase) and astrocytes (GFAP) were not obvious (Figure 10 C). Furthermore, immunocytochemistry revealed that
cells were positive for neuron-specific markers Tuj1, suggesting that they could be induced into neurons (Figure 10 D).
A. P 1 9 P 1 9 /h S h h N 0µµµµM FSK 10µM FSK B. GAPDH NF-M Nestin 0 2 4 2 4 0 2 4 2 4 FSK P19 P19/hshhN ╋ ╋╋ ╋ ━ ━━ ━ ━━━━ ╋╋╋╋ GAPDH NF-M Nestin 0 2 4 2 4 0 2 4 2 4 FSK P19 P19/hshhN ╋ ╋╋ ╋ ━ ━━ ━ ━━━━ ╋╋╋╋
C. FSK CNPase NF-M NCAM GFAP β ββ β-tubulin III std P19/hShhN P19 ╋ ╋╋ ╋ ━ ━ ━ ━ ━━━━ ╋╋╋╋ D. T u j1 D A P I Fsk P19/ShhN P19 ╋ ╋ ╋ ╋ ━ ━ ━ ━ ━━━━ ╋╋╋╋ Fsk
Fig. 10. Neuronal differentiation of P19, P19/hShhN cells. Morphological changes of P19 cells by Shh or Foskolin (10µM) were examined under a phase contrast microscope (Magnification : x200) (A). Expression of Nestin, as neural stem cell
marker, and NF-M, as neuronal marker, mRNA was measured by RT-PCR (B). Neuronal differentiation was measured by western blot analysis with anti-Tuj1 and
NF-M specific antibody (C) and Immunocytochemistry with anti-Tuj1 specific antibody (D).
6. Shh promotes neuronal progenitor cell/ motor neurons survival or proliferation
Nestin is expressed in the myotome the neural progenitor cells of the developing embryo. While the regulatory region for expression in myotome resides in the first intron of the Nestin gene, that for expression in neural progenitor cells exists in the second intron. To ascertain if Shh was important for neuronal progenitor cell survival we added Shh conditioned medium, as a secreted Shh, to cultures of P19/pNPEeGFP cell lines (Figure 11 A). By 4 days in culture, <4% of Nestin-positive cells were present in the control conditioned medium, whereas 25-30% survive in Shh conditioned medium. GFP-positive cells were increased by secreted Shh (Figure 11 B). The expression of Nestin was further confirmed by RT-PCR. Shh increased the GFP and Nestin expression driven by the Nestin promoter/enhancer (Figure 11 C).
A.
Nestin promoter EGFP Intron II pNPEeGFP B. C o n tr o l C M S h h N C M
Control IgG 5E1
P19/pNPEeGFP % n e s ti n -G F P (+ )/ T o ta l c e ll s IgG 5E1 Control CM ShhN CM 0 5 10 15 20 25 30 35 5 10 15 20 25 30 35 ╋ ╋ ╋ ╋ ━ ━ ━ ━ ━━━━ ╋╋╋╋ ╋ ╋ ╋ ╋ ━━━━ ╋╋╋╋ ━━━━ C. Nestin GAPDH Control CM ShhN CM IgG 5E1 ╋ ╋ ╋ ╋ ━ ━ ━ ━ ╋╋╋╋ ━ ━━ ━ ╋╋╋╋ ━ ━ ━ ━ ╋╋╋╋ ━ ━ ━ ━ 0 1 2 3 0 1 2 3 N e s ti n /G A P D H m R N A IgG 5E1 Control CM ShhN CM ╋ ╋ ╋ ╋ ━ ━ ━ ━ ╋╋╋╋ ━ ━ ━ ━ ╋╋╋╋ ━ ━━ ━ ╋╋╋╋ ━ ━ ━ ━
Fig. 11. eGFP-labeled neuronal precursors in P19/pNPEeGFP. Construct for generation of P19/pNPEeGFP (A). P19/pNPEeGFP cells were cultured in Shh conditioned medium or control conditioned medium and counted GFP-positive cells (B). RT-PCR analyzed expression of Nestin (C).
7. Shh induces motor neuron differentiation in P19 cells.
The induction of MN progenitors depends on Shh activity (Briscoe and
Ericson, 2001). To examine whether the spinal progenitors present in P19/hShhN can differentiate into MN progenitor, we monitored the expression of homeobox domain
and bHLH transcription factors. In vitro neuronal differentiation of P19 cells was induced by Shh and forskolin. The P19/ShhN cells showed the morphology of the
cells changed gradually. P19/hShhN cells formed neurite-like processes and showed a neuron-like morphology on 4 days. To examine whether the spinal progenitors
present in P19/hShhN cells can differentiate into MN progenitors, we monitored the expression of HD and bHLH transcription factors that delineate sets of neural
progenitor cells. Spinal MN progenitors are found in P19/hShhN cells and expressed Nkx6.1, Olig2 and Ngn2. To determine whether MNs are generated from progenitor
cells present in P19/hShhN cells, we analyzed the expression of Hb9 and ChAT using RT-PCR and Immunocytochemistry. P19/ShhN cells increased expression of Hb9
and ChAT, suggesting that they could be induced into motor neurons (Fig 12). Immunocytochemistry revealed that the differentiated P19/hShhN cells increased
0 1 2 3 4 1 2 3 4 0 µM FSK 10 µM FSK Nkx6.1 Olig2 Ngn2 NeuroD GAPDH ChAT Hb9 P19 Nkx6.1 Olig2 Ngn2 NeuroD GAPDH ChAT Hb9 P19/hShhN
Fig. 12. RT-PCR of the expression of HD, bHLH, and motor neuron specific markers in P19 and P19/hShhN. Neuronal induction of P19 cells was performed as described in materials and methods. P19 and P19/hShhN were treated with 10µM
forskolin for 4 days. The efficiency of MN differentiation were measured by
expression level of motor neuron marker using RT-PCT. (
-
/+ indicates that forskolin treatment)H b 9 T u j1 H b 9 T u j1 H b 9 H o e c h s t P19 P19/hShhN T u j1 10µM Fsk 0µM Fsk 10µM Fsk 0µM Fsk
Fig. 13. Shh expression induces motor neuron differentiation of P19 cells. Neuronal induction of P19 cells was performed as described in materials and methods. P19 and P19/hShhN were treated with 10µM forskolin for 4 days. Cells
were visualized by Hoechst (blue) and immuno-stained with neuron specific antibody, Tuj1 (green) and motor neuron specific antibody, Hb9 (red). (Magnification : x200)
P19 P19/hShhN T u j1 10µM Fsk 0µM Fsk 10µM Fsk 0µM Fsk C h A T T u j1 C h A T T u j1 C h A T H o e c h s t
Fig. 14. Shh expression induces motor neuron differentiation of P19 cells. Neuronal induction of P19 cells was performed as described in materials and methods. P19 and P19/hShhN were treated with 10µM forskolin for 4 days. Cells
were visualized by Hoechst (blue) and immuno-stained with neuron specific antibody, Tuj1 (green) and motor neuron specific antibody, ChAT (red). (Magnification : x200)
8. Shh induces dopaminergic neuron and motor neuron differentiation in P19 cells
Shh is expressed in ventral regions of the spinal cord, hindbrain, midbrain, and diencephalons. The specification of MN progenitor by Shh signaling is mediated
through the patterned expression of homeodomain (HD) and basic helix-loop-helix (bHLH) transcription factors (Briscoe and Ericson, 2001). Dopaminergic neurons are
induced at E9 (rat) by combinatorial action of Shh and FGF8, released by the floor plate and isthmus. To examine whether P19/hShhN can differentiate into MN
progenitors, we monitored the bHLH transcription factors by RT-PCR. Olig2, a bHLH transcription factor, increased the expression in P19/hShhN cells. The
induction of MN progenitors depends on Shh activity. To determine whether DAs are generated from P19 cells by Shh, we examined the expression of transcription factor
and other DAs specific protein in differentiation. We showed that Shh increased the expression of TH, Nurr1 and Olig2 in P19 cells with foskolin (Fig 15). To determine
whether MN progenitors generated from RA-exprosed P19/hShhN in current method, we analyzed the expression of Olig2, TH, and Nurr1 protein expressed selectively.
RA-exposed P19/hShhN increased expression of Olig2 and decreased expression of TH and Nurr1. In the Immunocytochemistry, P19/hShhN cells increased expression
of Hb9 and TH, but no Hb9+ cells co-expressed TH, a marker dopaminergic neurons (Fig 16). These results provide evidence that Shh signaling is significant the
induction of DAs and MNs in current differentiation method.
TH GAPDH Nurr1 Olig2 P19 P19/hShhN 10 µM FSK P19 P19/h ShhN RA TH GAPDH Nurr1 Olig2 ╋ ╋ ╋ ╋ ━ ━━ ━ ╋ ╋ ╋ ╋ ━ ━ ━ ━
Fig. 15. Motor neurons and Dopamine neurons of differentiation in P19 cells. P19 and P19/hShhN cells were differentiated for 4days with or without RA(0.5µM) in N2 media. RT-PCR analyzed expression of TH and Nurr1, as a Dopamine neurons
T H H b 9 T H H b 9 H o e c h s t T H H b 9 T H H b 9 H o e c h s t P19 P19/hShhN 10µM Fsk 0µM Fsk 10µM Fsk 0µM Fsk
Fig. 16. Shh expression induces dopamine neurons and motor neurons differentiation of P19 cells. Neuronal induction of P19 cells was performed as described in materials and methods. P19 and P19/hShhN were treated with 10µM forskolin for 4 days. Cells were visualized by Hoechst (blue) and immuno-stained
with dopamine neuron specific antibody, TH (green) and motor neuron specific antibody, Hb9 (red). (Magnification : x200)
9. Shh promotes neuronal differentiation in P19 cells without treatment of RA or aggregation culture.
Our previous study suggested that P19 cells, when replated as the aggregates, could differentiate into neural cells with RA (McBurney et al., 1987). However,
P19/hShhN cells could differentiate into neurons without RA or aggregation culture. To compare whether P19 cells were differentiated into neurons by Shh without
treatment of RA or aggregation culture, P19 and P19/hShhN cells induced differentiation with and without treatment of RA or aggregation culture. Cells were
differentiated in the absence or presence of 10µM forskolin. RT-PCR analysis of RNA isolated from differentiated P19 and P19/hShhN cells, which indicated that the
expression of marker for neuron, NFM, for DAs, TH and Nurr1, and for MNs, Olig2 increased by Shh in current differentiation, but not in conventional differentiation
(Figure 17). P19/hShhN cells could differentiate into neuronal cells without treatment of RA or aggregation culture. Compared to the conventional method, the
current differentiation method without aggregation may provide a useful system for the study of the functions of Shh.
Nestin NF-M TH Nurr1 Olig2 GAPDH FSK RA
Aggregate Agg. + Differentiation Nonagg. + Differentiation
hShhN ╋ ╋ ╋ ╋ ━ ━━ ━ ╋ ╋ ╋ ╋ ━ ━ ━ ━ ╋ ╋ ╋ ╋ ━ ━ ━ ━ ╋ ╋╋ ╋ ━ ━━ ━ ━━━━ ╋╋╋╋ ━━━━ ╋╋╋╋ ━ ━ ━ ━ ╋ ╋ ╋ ╋ ╋╋╋╋ ╋╋╋╋ ╋╋╋╋ ╋╋╋╋ ━━━━ ━━━━ ━━━━ ━━━━ ╋ ╋╋ ╋ ━ ━ ━ ━ ━━━━ ╋╋╋╋ ━━━━ ━━━━ ╋╋╋╋ ╋╋╋╋
Fig. 17. Shh promotes neuronal differentiation in P19 cells without treatment of RA or aggregation culture. P19 cells were differentiated into neurons by Shh without treatment of RA or aggregation culture, P19 and P19/hShhN cells induced differentiation with and without treatment of RA or aggregation culture. Cells were
differentiated in the absence or presence of 10µM forskolin. RT-PCR analyzed expression of TH and Nurr1, as a Dopamine neurons marker and Olig2, as a Motor
IV. DISCUSSION
In this study, we have further addressed the function of Shh in the proliferation and the motor neuron differentiation of P19 cells. Our studies reveal that
overexpression of Shh can direct motor neuron differentiation of P19 cells in the absence of RA, when P19/hShhN cells were replated as the single sell suspension.
Furthermore, we show that Shh-overexpressing P19 cells differentiate exclusively into motor neuron.
Chemical inducers and aggregation are two key elements to impart fate choices of P19 cells. With aggregation, DMSO directs P19 cells to differentiate into
mesoderm origin muscle cells. RA, however, induces P19 cells to differentiate into ectoderm-derived neural cells including neurons and astrocytes (E.M.
Jones-Villeneuve et al., 1982, 1983; M.W. McBurney et al., 1982). According to the treatment of cells, the neural differentiation of RA-induced P19 cells can be divided
into two sequential stages, a stage of induction and a stage of differentiation. During the first stage, P19 cells are allowed to aggregate in the Petri dish and induced with
RA for four days. Based on cell morphology changes and gene expression profiles, it seems that pluripotent P19 embryonic carcinoma (EC) cells are determined into
neural progenitor cells during the first RA-induction stage. In second stage, the induced P19 cells are replated into cell culture dish as the single cell suspension or
aggregates and left to differentiate into mature neurons and astrocytes.
Binding of Shh to Ptc releases this receptor’s repression of Smo, which then transduces the signal by acting on the transcription factor of Gli family. The
transcriptionally activating Gli forms then upregulate Shh targets, including ptc, gli1, gli2 and Shh itself. Shh signaling may directly regulate the cell cycle, as it can upregulate the expression of G1-phase cyclins of type D and E, and Pathed can act directly on the G2-phase cyclin B. Alternatively, Shh other activity may activate
other signaling system that control cell proliferation. We observed that the Shh receptor Pathed and Smoothend and the target gene Gli is expressed in P19 cells. We
have showed that Shh regulates the proliferation of P19 cells using growth kinetics and MTT assay. P19/hShhN cells increased the number of total cells in growth and
differentiation condition and neutralized by 5E1. The biological activity of the Shh conditioned media was almost completely inhibited by 5E1, but unaffected by IgG.
The results suggest that the secreted hShhN was responsible for the stimulation of P19 cell proliferation .We conclude that Shh directly promotes P19 cells proliferation
in vitro.
As the neurogenesis in vivo, neurons appear earler than glial cells during
RA-induced P19 cell neural differentiation (E.M. Jones-Villeneuve et al., 1982). Interestingly, our results show that P19/hShhN cells differentiate restrictively into
neurons, but not glial cells. Our previous study suggested that P19/ShhN, when replated as the aggregates, could differentiate into neural cells without RA induction.
However, RA-inducted P19 cells were normally replated as the single cell suspension to enrich the cells of interest during their neural differentiation (M.W McBurney et
al., 1987, 1995). The wild-type P19 cells were used as the negative control. At day 3, P19/hShhN cells started to send out long neurite-like process. At day 4, P19/hShhN
cells sent out many neurite-like process to form the networks. In parallel, the expression of Tuj1 and NF-M, as a neuronal specific marker, increased during the
differentiation of P19/hShhN. This suggests that Shh may promote the neuronal differentiation in P19 cells without RA or aggregation culture.
Shh is necessary for the induction of both spinal motor neurons and midbrain dopaminergic neurons. In the developing midbrain, Shh was first characterized for its
ability to induce the production of dopaminergic neurons. In the developing spinal cord, the induction of motor neuron progenitors depends on Shh activity. We have
showed that Shh promotes motor neuron differentiation than dopaminergic neurons in current differentiation method. P19/hShhN cells increased expression of motor
neurons transcription factor, homeodomain or specific marker gene, Nkx.1, Olig2, Ngn2, NeuroD, Isl -1, Hb9 and ChAT. But, the expression of TH decreased in
P19/hShhN cells. Nkx6.1 is expressed in progenitors for both motor neurons and ventral interneurons. The expression of Olig2 and Ngn2 was further correlated with
subtype-specific neuronal marker at E12.5, the stage when progenitors actively generate various neuronal subtype. The onset of Olig2 and Ngn2 expression precedes
those of Hb9 and Isl -1, indicating that they are one of the earlist transcription factors expressed in the motor neuron lineage. Thus, among many members of bHLH factors,
the coexpression of Olig2 and Ngn2 is specifically correlated to motor neuron generation. Isl-1 and Hb9 are defined markers for motor neurons and their
progenitors in the ventral neural tube (Tsuchida et al., 1994; Arber et al., 1999; Thaler et al., 1999). These results suggest that Shh could be promoted into motor
neuron progenitor cells and motor neuron differentiation and expressed transcription factors and HD.
The results here supported previous suggestions that Shh may be useful vehicles for a variety of neurological diseases, as Parkinson disease or Spinal cord
V. CONCLUSIOS
Shh increases both proliferation and neuronal differentiation in P19 cells and
promote the neuronal differentiation in P19 cells without RA or aggregation culture. In the neuronal differentiation, Shh may promote proliferation of neuronal precursor
cells and thereby increase expression of motor neuron specific proteins. Compared to the conventional method, the current differentiation method without aggregation may
REFERENCES
1. Arber S, Han B, Mendelsohn M, Smith M, Jessell TM, Sockanathan S: Requirement for the homeobox gene Hb9 in the consolidation of motor
neuron identity. Neuron 23(4):659-74, 1999
2. Ariel Ruiz i Altaba, Verónica Palma & Nadia Dahmane: Hedgehog-Gli signalling and the growth of the brain. Nature Reviews Neuroscience
3(1):24-33, 2002
3. Barberi T, Klivenyi P, Calingasan NY, Lee H, Kawamata H, Loonam K, Perrier AL, Bruses J, Rubio ME, Topf N, Tabar V, Harrison NL, Beal MF,
Moore MA, Studer L: Neural subtype specification of fertilization and nuclear transfer embryonic stem cells and application in parkinsonian mice.
Nat Biotechnol. 10:1200-1207, 2003
4. Barnes EA, Kong M, Ollendorff V, Donoghue DJ: Patched1 interacts with cyclin B1 to regulate cell cycle progression. EMBO J 20(9):2214-2223, 2001
5. Bertrand S, Cazalets JR: The respective contribution of lumbar segments to
the generation of locomotion in the isolated spinal cord of newborn rat. Eur J Neurosci 16(9):1741-1750, 2000
6. Briscoe J, Ericson J: Specification of neuronal fates in the ventral neural tube. Curr Opin Neurobiol 11(1):43-49, 2001
7. Briscoe J, Ericson J: The specification of neuronal identity by graded Sonic
Hedgehog signalling. Semin Cell Dev Biol 10(3):353-362, 1999
8. Briscoe J, Pierani A, Jessell TM, Ericson J: A homeodomain protein code specifies progenitor cell identity and neuronal fate in the ventral neural tube.
Cell 101(4):435-445, 2000
9. Chiang C, Litingtung Y, Lee E, Young KE, Corden JL, Westphal H, Beachy PA: Cyclopia and defective axial patterning in mice lacking Sonic hedgehog
gene function. Nature 383(6599):407-413, 1996
10. E.M. Jones-Villeneuve, M.A. Rudnicki, J.F. Harris, M.W. McBurney: Retinoic acid-induced neural differentiation of embryonal carcinoma cells.
Mol. Cell. Biol. 3 2271–2279, 1983
11. E.M. Jones-Villeneuve, M.W. McBurney, K.A. Rogers, V.I. Kalnins: Retinoic acid induces embryonal carcinoma cells to differentiate into neurons
12. Ericson J, Morton S, Kawakami A, Roelink H, Jessell TM: Two critical periods of Sonic Hedgehog signaling required for the specification of motor
neuron identity. Cell 87(4):661-673, 1996
13. Ericson J, Muhr J, Placzek M, Lints T, Jessell TM, Edlund T: Sonic hedgehog induces the differentiation of ventral forebrain neurons: a common signal for
ventral patterning within the neural tube. Cell 81(5):747-756, 1995
14. Fan H, Khavari PA: Sonic hedgehog opposes epithelial cell cycle arrest. J Cell Biol 147(1):71-6, 1999
15. Fu M, Lui VC, Sham MH, Pachnis V, Tam PK: Sonic hedgehog regulates the
proliferation, differentiation, and migration of enteric neural crest cells in gut. J Cell Biol 166(5):673-684, 2004
16. Goodrich LV, Scott MP: Hedgehog and patched in neural development and
disease. Neuron 21(6):1243-1257, 1998
17. Hynes M, Porter JA, Chiang C, Chang D, Tessier-Lavigne M, Beachy PA, Rosenthal A: Induction of midbrain dopaminergic neurons by Sonic
18. Ingham PW, McMahon AP: Hedgehog signaling in animal development: paradigms and principles. Genes Dev 15(23):3059-3087, 2001
19. Jessell TM: Neuronal specification in the spinal cord: inductive signals and
transcriptional codes. Nat Rev Genet 1(1):20-29, 2000
20. Karen Lai, Brian K. Kaspar, Fred H. Gage and David V. Schaffer: Sonic hedgehog regulates adult neural progenitor proliferation in vitro and in vivo.
Nat Neuroscience 654:21-27, 2003
21. Kenney AM, Rowitch DH: Sonic hedgehog promotes G(1) cyclin expression and sustained cell cycle progression in mammalian neuronal precursors. Mol
Cell Biol 20(23):9055-9067, 2000
22. Kim JY, Koh HC, Lee JY, Chang MY, Kim YC, Chung HY, Son H, Lee YS, Studer L, McKay R, Lee SH: Dopaminergic neuronal differentiation from rat
embryonic neural precursors by Nurr1 overexpression. J Neurochem 85(6):1443-1454, 2003
23. Lee SK, Pfaff SL: Synchronization of neurogenesis and motor neuron
specification by direct coupling of bHLH and homeodomain transcription factors. Neuron 38(5):731-45, 2003