저작자표시-비영리-변경금지 2.0 대한민국 이용자는 아래의 조건을 따르는 경우에 한하여 자유롭게 l 이 저작물을 복제, 배포, 전송, 전시, 공연 및 방송할 수 있습니다. 다음과 같은 조건을 따라야 합니다: l 귀하는, 이 저작물의 재이용이나 배포의 경우, 이 저작물에 적용된 이용허락조건 을 명확하게 나타내어야 합니다. l 저작권자로부터 별도의 허가를 받으면 이러한 조건들은 적용되지 않습니다. 저작권법에 따른 이용자의 권리는 위의 내용에 의하여 영향을 받지 않습니다. 이것은 이용허락규약(Legal Code)을 이해하기 쉽게 요약한 것입니다. Disclaimer 저작자표시. 귀하는 원저작자를 표시하여야 합니다. 비영리. 귀하는 이 저작물을 영리 목적으로 이용할 수 없습니다. 변경금지. 귀하는 이 저작물을 개작, 변형 또는 가공할 수 없습니다.
농학석사 학위논문
Isolation and Characterization of Nitrogen-fixing
Bacteria and Phosphate-solubilizing Bacteria
from Oriental Melon Farm Soil
참외 농장 토양에서의 질소고정균과
인산 가용화균의 분리 및 특성 연구
2020 년 2 월
서울대학교 대학원
농생명공학부 식물미생물학전공
장 소 현
A THESIS FOR THE DEGREE OF MASTER OF SCIENCE
Isolation and Characterization of Nitrogen-fixing
Bacteria and Phosphate-solubilizing Bacteria
from Oriental Melon Farm Soil
BY
SO HYUN JANG
Department of Agricultural Biotechnology
The Graduate School of Seoul National University
ABSTRACT
Isolation and Characterization of Nitrogen-fixing
Bacteria and Phosphate-solubilizing Bacteria
from Oriental Melon Farm Soil
SO HYUN JANG Major in Plant Microbiology Department of Agricultural Biotechnology The Graduate School of Seoul National University
Oriental melon is one of the most popular summer fruits in Korea. A main producing area of oriental melon is Seongju-gun and more than 95% of oriental melon is cultivated under structure. Especially, salts accumulation in greenhouse soil is one of the most important problems for growing oriental melons. Salts accumulation, mainly caused by overuse of fertilizer, reduces plant growth rate by impeding nutrient uptake of plants. To relieve salts accumulation, one of the solutions is using microbial fertilizer, with reducing use of chemical fertilizer. Thus, using nitrogen-fixing bacteria (NFB) and phosphate-solubilizing bacteria (PSB) as microbial fertilizer for oriental melon farm can be a key for solving problems of cultivation under structure. The objective of this study is to isolate and characterize potential candidate bacteria of microbial fertilizer for oriental melon cultivation. Bacteria were isolated from six soil samples from two different oriental melon farms by using NFb, JMV and N2F agar for isolation of NFB, and PVK and NBRIP-BPB agar for isolation of PSB. After grouping isolates through REP-PCR pattern analysis, isolates were identified by 16S rDNA sequencing. Color
ii
changing observation of NFb semi-solid media was used as assessment of NFB. Measurement of phosphate solubilization index (PSI) and soluble phosphate concentration by phosphate colorimetric kit (MAK 030) was performed as qualitative and quantitative assessment of PSB, respectively. 145 strains were isolated from nitrogen fixation selective media and among them promising 10 strains were selected based on assessment of nitrogen fixing ability. 37 strains were isolated from phosphate-solubilizing selective media and among them promising 15 strains were selected based on assessment of phosphate solubilizing ability. Assessment of other plant growth promoting properties such as IAA production, extracellular protease activity, cellulose-degrading ability, and siderophore producing ability were performed to selected isolates. Among 25 selected isolates, all isolates produced low level of IAA and exhibited siderophore-producing ability, 19 isolates exhibited extracellular protease ability, and 15 isolates exhibited cellulose-degrading ability. Screening of selected bacteria for temperature range of growth and optimal growth was also performed to provide further information to determine application methods of microbial fertilizer. Thus, in this study, based on analysis of bacteria isolated from oriental melon farm soil for plant growth promoting properties, promising candidates for microbial fertilizer were selected.
Keywords: Microbial fertilizer, Nitrogen fixation, Phosphate solubilization, Oriental melon, plant growth promoting properties
CONTENTS
ABSTRACT ...
page
ABSTRACT ... ⅰ
CONTENTS ... ⅲ
LIST OF TABLES ... ⅴ
Ⅰ. INTRODUCTION ... 1
Ⅱ. MATERIALS AND METHODS ... 6
1. Isolation of PSB and NFB ... 6
2. Identification by 16S rDNA gene sequence analysis ... 9
3. Assessment of nitrogen-fixing bacteria ... 10
4. Assessment of phosphate-solubilizing bacteria ... 11
5. Analysis of auxin-producing ability ... 13
6. Analysis of siderophore-producing ability ... 14
7. Analysis of extracellular-protease producing ability ... 15
8. Analysis of cellulose-degrading ability ... 16
iv
Ⅲ. RESULTS ... 18
1. Isolation and Identification of possible candidates for NFB and PSB ... 18
2. Assessment of nitrogen-fixing ability ... 29
3. Assessment of phosphate-solubilizing ability ... 33
4. Analysis of IAA-producing ability of the selected isolates ... 38
5. Analysis of siderophore-producing ability of the selected isolates ... 40
6. Analysis of extracellular-protease producing ability of the selected isolates .. 43
7. Analysis of cellulose-degrading ability of the selected isolates ... 46
8.Analysis of temperature range for growth and optimal growth of the selected isolates ... 49
Ⅳ. DISCUSSION ... 52
Ⅴ. LITERATURE CITED ... 57
LIST OF TABLES
ABSTRACT ...
page
Table 1. Isolates from NFb agar ... 20
Table 2. Isolates from JMV agar ... 21
Table 3. Isolates from N2F agar ... 22
Table 4. Isolates from PVK agar ... 27
Table 5. Isolates from NBRIP-BPB agar ... 28
Table 6. Assessment of nitrogen-fixing bacteria ... 30
Table 7. Selected nitrogen-fixing isolates ... 32
Table 8. Qualitative assessment of phosphate-solubilizing bacteria ... 34
Table 9. Quantitative assessment of phosphate-solubilizing bacteria .... 36
Table 10. Selected phosphate-solubilizing isolates ... 37
Table 11. Analysis of IAA-producing ability of the selected isolates ... 39
Table 12. Analysis of siderophore-producing ability of the selected
isolates ... 41
Table 13. Analysis of extracellular-protease producing ability of the
selected isolates ... 44
Table 14. Analysis of cellulose-degrading ability of the selected
isolates ... 47
vi
Table 15. Analysis of temperature range for growth and optimal growth
INTRODUCTION
In Korea, oriental melon is one of the most major summer fruit which were produced 130,528 tons in 2018 (KOSIS 2019). Over than 94% of cultivation area is converged in Gyeongsangbuk-do: the main cultivation area of oriental melon is Seongju-gun, Gyeongsangbuk-do where contributes 91.9 % of total oriental melon production in Korea in 2013 (GBARES 2015; KOSIS 2014).
About 97 % of total oriental melon production area are using cultivation under structure method, as a production efficiency of cultivation under structure is much higher than that of cultivation on bare ground (GBARES 2015; KOSIS 2014). Cultivation under structure, as known as protected cultivation, is one of main cultivation method for various agricultural plants (Cho et al. 2006). Cultivating plants in greenhouse can adjust internal condition in structure to optimal environment for growing plants, that can overcome rough raw condition such as climate, season, location, meteorological disasters and social background (Sonneveld and Voogt 2009; 이병일 et al. 2004). Protected cultivation also has several problems, however, the main problem that inhibit plant growth in protected cultivation is salt accumulation in soil (Jeon et al. 2005).
Salt accumulation in soil is one of the main causes of growth disorder in protected cultivation of oriental melon. Salinization of soil is very common in greenhouse: 59 % of greenhouse in Korea, salinization of greenhouse soil is reported (RDA 2004). In addition, over than 3 years of
2
serial cultivation without treatment for salt accumulation, soil condition turns to unavailable for plant growth (유철성 1996). Ions such as PO4-, K+, Mg 2+,
Na+, SO42-, and Cl- are accumulated commonly in greenhouse soil (이병일 et
al. 2004). Not all of fertilized nutrients are uptake by plants but immobilized and last in soil which considered as the main cause of salt accumulation in greenhouse soil. On a bare ground, fertilized nutrients are washed by rainfall, however, rainfall is intercepted by structure and causes salts to accumulate in protected cultivation soil (Oh et al. 2010). Nevertheless, long cultivation term and successive cropping in greenhouse induce extensive use of chemical fertilizer (Shaharoona et al. 2008). In addition, high temperature in greenhouse increase vaporization that enhance soil salinization (Oh et al. 2010). Salt tolerance of each plants is different, however, ability of root for up-taking water and nutrient from soil is significantly decrease on certain salt concentration because of osmotic pressure (Bernstein 1975; Oh et al. 2010). To relieve this problem, several treatments such as planting fertilizer-absorbing plants, growing salt-tolerance cultivar, soil replacement, deep tillage, and allowance of rainfall in rainy season are performed (Oh et al. 2010; RDA and NAS 2014). In these days, using microbial fertilizers which contain microbes with soil management ability and plant growth promoting properties is getting attention.
Phosphate, essential macro-elements for plants, is one of the major components of chemical fertilizer. Because of its importance, overuse of phosphate fertilizers frequently happens. However, only 10-20 % of phosphate given by chemical fertilizer is up-taken by plants and remained
phosphate in soil immobilized with cations in soil, becoming inaccessible for plant.(EA and Clark 1989; Lee et al. 2012) These remaining phosphate is accumulated with repeatedly given chemical fertilizer and inhibit plant growth in cultivation under structure. Phosphate-solubilizing bacteria (PSB) are bacteria that can mobilize insoluble phosphate not only from accumulated phosphate by fertilizer, but also from organic form of phosphate which consist 4-90 % of total soil phosphate (Lee et al. 2012). Increase of plant root and top growth by phosphate and improvement of symbiotic nitrogen fixation efficiency in legume nodule were reported (Armstrong et al. 1999). Thus, application of PSB to greenhouse soil can be an efficient treatment for salinized soil and reducing chemical fertilizer.
Nitrogen fertilizers are one of the largest produced and treated fertilizer in the world: consumption of nitrogen fertilizers is sharply increased from 11.6 million tons (1961) to 104 million tons (2006) worldwide (Rakshit and Singh 2018). However, about 67 % of nitrogen given by fertilizer loss by leaching, ammonia volatilization, and denitrification, which leads to 15.9 billion dollar loss worldwide and enhancing greenhouse effect by their production process (Choi et al. 2003; Raun and Johnson 1999). Nitrogen is most abundant element in atmosphere. However, it is not available form for plants as plants can only use NO3- and NH4+ form of nitrogen. For the most part of nitrogen fixation in nature, converting from N2 to NH4+, is performed
by nitrogen-fixing bacteria. As about 95 % of worldwide terrestrial nitrogen flow occurs in microorganism-plant-soil system, a great part of plant-available nitrogen input in soil is provided by biological nitrogen fixation (Rosswall
4
1976). Nitrogen fixing can be performed by symbiotic and asymbiotic nitrogen-fixing bacteria (NFB) (Gupta et al. 2015). Using microbial fertilizer with NFB can be effective by providing nitrogen for plants as well as reducing required dose of chemical fertilizer. In the context of a reducing use of fertilizer also, using NFB as microbial fertilizer can contribute to relieving salt accumulation problems in greenhouse soil too.
Bacteria that can promote plant growth are called as plant growth promoting bacteria (PGPB). PGPBs enhance plant growth by diverse ways, such as phosphate solubilizaton, nitrogen fixation, phytohormone secretion, siderophore production, and hydrolytic enzyme production (Gupta et al. 2015; Jha and Saraf 2015).
Phytohormones such as auxins, cytokinins, and gibberelins are produced by some rhizobacteria (Jha and Saraf 2015). Among various phytohormones, auxin is critically required as a positive regulator of plant growth involved in various physiological responses including root and stem growth, reproductive growth, seed germination, and enhancement of nutrition uptake by improving root structures (Jha and Saraf 2015; Kim and Song 2012; Woodward and Bartel 2005; Anwar et al. 2016). Indole-3-acetic acid (IAA) is a typical auxin that produced by microbes in soil through diverse mechanism (Kim and Song 2012).
Soil bacteria produce siderophores, a small molecule that enhance iron uptake by chelating Fe3+ ion in soil and transporting it into cells. (Jha and Saraf 2015). Siderophore excreted by bacteria is available for plants and stimulates plant growth by supporting plant iron uptake (Alexander and
Zuberer 1991; Crowley et al. 1987). Siderophore secretion by rhizobacteria can also inhibit proliferation of harmful plant bacterial and fungal pathogen through sequestering available iron ion (Kloepper et al. 1980; Leong 1986).
Rhizobacteria produce diverse lytic enzymes such as cellulase, protease, phosphatase, chitinase, and lipase that can degrade cell wall of plant fungal pathogen and decompose insoluble component in soil. This causes suppression of pathogens and enhancement soil organic carbon and nitrogen contents (Geisseler and Horwath 2009; Gupta et al. 2015; Jung et al. 2007).
The aim of this study is to isolate and characterize nitrogen-fixing bacteria and phosphate-solubilizing bacteria from oriental melon farm soil, and screening possible candidates of microbial fertilizer for oriental melon. Isolated bacteria were screened by its nitrogen-fixing ability and phosphate solubilizing ability. Additional traits such as plant growth promoting properties, IAA production, siderophore production, extracellular protease production, and cellulose degrading ability of selected useful bacteria were examined. Also, temperature range of growth of preferred bacteria was investigated to consider their application in greenhouse soil along with production cycle of oriental melon as well.
6
MATERIALS AND METHODS
1. Isolation of PSB and NFB
Six soil samples were obtained from rhizosphere of oriental melon: three soil samples were collected at oriental melon farm located in Daehwang-ri, Seongju-eup, Seongju-gun, Gyeongsangbuk-do and other three soil samples were collected at oriental melon farm located in Sangeon-ri, Yongam-myeon, Seongju-gun, Gyeongsangbuk-do. Collected soil samples were sieved through a 2 mm aperture mesh and kept at 4 ℃ prior to use. 1 g of each soil sample was homogenized with 9.5 ml of sterilized 0.85 % saline solution and serially diluted up to 10-3, 10-4, 10-5, and 10-6 using sterilized 0.85 % saline solution. 100μl of each dilution was spread on the surface of PSB or NFB selection agar medium plate. Inoculated agar media plates were incubated at 28 ℃.
For isolation of NFB, NFb medium, JMV medium and N2-Fixing
Bacteria (N2F) medium were used as selection media. NFb medium contained (per liter) 5 g of Malic acid, 0.5g of K2HPO4, 0.2 g of MgSO4∙7H2O, 0.1 g of
NaCl, 0.02 g of CaCl2∙2H2O, 2 ml of micronutrient solution, 2 ml of
bromothymol blue solution, 4 ml of FeEDTA solution, 1 ml of vitamin solution, 4.5 g of KOH, 15 g of agar and adjusted pH to 6.5 before autoclaving. Micronutrient solution contained (per liter) 0.04 g of CuSO4∙5H2O, 0.12 g of ZnSO4∙7H2O, 1.4 g of H3BO3, 1.0 g of
NaMoO4∙2H2O, 1.175 g of MnSO4∙H2O. Bromothymol blue solution
solution contained 16.4 g of FeEDTA per liter. Vitamin solution contained 10 mg of biotin, 20 mg of pyridoxal-HCl in 100 ml of water with dissolving in hot water bath (Dobereiner et al. 1976). JMV agar contained (per liter) 5 g of mannitol, 1.8 g of KH2PO4, 0.6 g of K2HPO4, 0.2 g of MgSO4∙7H2O, 0.1 g of
NaCl, 0.2 g of CaCl2∙2H2O, 2 ml of micronutrient solution (see above), 2 ml
of bromothymol blue solution (see above), 4 ml of FeEDTA solution (see above), 1 ml of vitamin solution (see above), 25 g of agar, and and adjusted pH to 5.0-5.4 with KOH before autoclaving (Baldani et al. 2014). N2F medium contains (per liter) 0.4 g of KH2PO4, 0.1 g of K2HPO4, 0.2 g of
MgSO4∙7H2O, 0.1 g of NaCl, 10.0 mg of FeCl3, 2.0 mg of NaMoO4∙2H2O, 5 g
of Sodium malate, 50 mg of yeast extract, 15 g of agar, and adjusted pH to 7.2
± 0.1 before autoclaving (Smibert et al. 1994). Isolates with confirmation of growth on medium were selected as possible candidates of NFB.
For isolation of PSB, dilutions were spread on two PSB selection media, Pikovskaya (PVK) agar medium and National Botanical Research Institute’s phosphate growth medium containing bromophenol blue (NBRIP-BPB medium). PVK medium contained (per liter) 10 g of Glucose, 0.5 g of
yeast extract, 0.5 g of (NH4)2SO4, 0.1 g of MgSO4∙7H2O, 0.2 g of MnSO4∙H2O,
0.2 g of FeSO4∙7H2O, 0.2 g of NaCl, 0.2 g of KCl, 5 g of Ca3(PO4)2 and 15 g
of agar (Surange et al. 1997). NBRIP-BPB medium contained (per liter)10 g of Glucose, 5 g of MgCl2∙6H2O, 5 g of Ca3(PO4)2, 0.25 g of MgSO4∙7H2O, 0.2
g of KCl, 0.1 g of (NH4)2SO4, 0.025 g of BPB, 18 g of agar, and adjusted pH
8
halo zone in PVK medium or NBRIP-BPB medium were selected as candidates of PSB.
To distinguish isolated possible candidates PSB and NFB whether there are same bacteria at strain level, colony repetitive palindromic polymerase chain reaction (REP-PCR) was performed. BOXA1R primer (5’-CTACGGCAAGGCGACGCTGACG-3’) was used as previously described (De Bruijn 1992; Kim et al. 2013). Each isolate was grown on agar medium to form single pure colonies and a small amount of cells was suspended with 25 μl of PCR mixture. PCR mixture contained (per 25 μl ) 0.3 μl of distilled water, 12.5 μl of dNTP mixture (2.5 mM each), 5.0 μl of 5 X Gitschier buffer, 2.5 μl of dimethyl sulfoxide, 1.0 μl of BOXA1R primer (50 pmol μl -1), 0.4 μl of bovine serum albumin (10 mg ml-1), 0.8 μl of Taq
polymerase (5 U ml-1) with 2.5 μl of template DNA (Rademaker et al. 1998). Gitschier buffer was made by method described in Korgan et al. (1987). Condition for REP-PCR cycles were described in sequence as follows: 1 cycle of step 1 at 93 ℃ for 7 min; 35 cycles with step 2, 3, and 4 (step 2 at 92 ℃ for 1 min, step 3 at 52 ℃ for 1 min, and step 4 at 65 ℃ for 8 min); 1 cycle of step 5 at 65 ℃ for 16 min; and a final soak at 4 ℃. Electrophoresis on 1.2 % agarose gel was performed after the reaction. REP-PCR bands were visualized by UV trans-illumination (306nm). Isolates with identical REP-PCR band are considered as same strains and only one of same isolates was remain in the list.
2. Identification by 16S rDNA gene sequence analysis
Based on the selecting procedure using colony REP-PCR analysis, each of isolates was conducted 16S rDNA gene sequence analysis with 27mf and 1492r as previously described for identification (Baker et al. 2003). A small amount of cells was suspended with 50 μl of PCR mixture, which contained 5.0 μl of 10X buffer, 2.5 μl of 0.1 % BSA, 5.0 μl of 100 % DMSO, 5.0 μl of dNTP mixture (2.5 mM each), 2.5 μl of 27mf primer (20 pmol μl-1),
2.5 μl of 1492r primer (20 pmol μl-1), 0.5 μl of Taq polymerase (5 U μl-1), 22
μl of distilled water, and 5.0 μl template DNA. The cycles used were as follows: 1 cycle of step 1 at 94 ℃ for 5 min; 29 cycles with step 2, 3, and 4 (step 2 at 94 ℃ for 1 min, step 3 at 55 ℃ for 1 min, and step 4 at 72 ℃ for 1 min); 1 cycle of step 5 at 72 ℃ for 10 min; and a final soak at 4 ℃.
PCR products were purified using QIAquick PCR Purification Kit (Qiagen) and sequenced using an ABI Prism BigDye Terminator Cycle Sequencing Ready Kit (Perkin-Elmer) with the sequencing primers 515r, 926f and 1055r (Stackebrandt and Goodfellow 1991). Using method previously described by Kim et al. (2017), almost full length 16S rRNA gene were sequenced. 16S rRNA gene sequences were identified by comparing with sequences of bacterial type strains from the EzBioCloud server database (Chun et al. 2007; Yoon et al. 2017).
10
3. Assessment of nitrogen-fixing bacteria
Nitrogen-fixing ability of isolated NFB candidates were tested by color changing observation of NFB semi-solid agar medium (Kim et al. 2012). NFb semi-solid agar medium contained (per liter) 5 g of Malic acid, 0.5 g of K2HPO4, 0.2 g of MgSO4∙7H2O, 0.1 g of NaCl, 0.02 g of CaCl2∙2H2O, 2 ml of
micronutrient solution (see above), 2 ml of bromothymol blue solution (see above), 4 ml of FeEDTA solution (see above), 1 ml of vitamin solution (see above), 4.5 g of KOH, 1.6 g of agar and adjusted pH to 6.5 before autoclaving (Baldani et al. 2014; Dobereiner et al. 1976). Growth on NFb agar medium of each isolates were pretested. The isolates were inoculated in NFb semi-solid agar medium and incubated at 26 ℃. Green to blue color change of NFb semi-solid media indicates that inoculated isolate has nitrogen-fixing ability (Kim et al. 2012). KACC 11805 (Azotobacter chroococcum) and KACC 16233 (Pleomorphomonas diazotrophica) were regarded as positive controls (Damir et al. 2011; Madhaiyan et al. 2013).
4. Assessment of phosphate-solubilizing bacteria
Assessment of phosphate-solubilizing bacteria was conducted via qualitative and quantitative study. Qualitative assessment of phosphate-solubilizing bacteria was performed by measurement of phosphate solubilization index (PSI). Isolates were incubated in R2A broth (LAB M, UK) at 26 ℃, 150 rpm. Inoculum was prepared as OD600=0.5 by
centrifugation and 20 μl of each inoculum was dropped on the middle of NBRIP-BPB agar medium surface and incubated at 26 ℃. Diameter of the colony and halo zone was measured at 7 and 14 days after inoculation. PSI of each isolates were calculated as follows: PSI = (Diameter of the halo zone) / (Diameter of the colony) (Kumar and Narula 1999). KACC 14533 (Chitinophaga oryziterrae) was regarded as a positive control (Chung et al. 2012)
Based on the results of the qualitative methods, several isolates were selected for quantitative study. Quantitative assessment of phosphate-solubilizing bacteria was conducted by measuring soluble phosphate concentration via phosphate colorimetric kit MAK 030 (Sigma-Aldrich, USA). Each selected isolate was incubated in R2A broth (LAB M) and cells were harvested by centrifugation and washed 2 times using 0.85% saline. Pellets were resuspended with 0.85 % saline to prepare inoculum adjusted OD600 to
0.5 and inoculated in NBRIP-BPB broth media with 1 % (V/V). NBRIP-BPB broth media contained (per liter)10 g of glucose, 5 g of MgCl2∙6H2O, 5 g of
Ca3(PO4)2, 0.25 g of MgSO4∙7H2O, 0.2 g of KCl, 0.1 g of (NH4)2SO4, 0.025 g
12
Inoculated NBRIP-BPB broth media were incubated for 10 days at 28 ℃, 150 rpm in shaking incubator. After 10 days of incubating, cell cultures were centrifuged at 14000 rpm at 4 ℃ for 10 minutes to obtain supernatant. Soluble phosphate concentration was measured by manufacturer’s instruction of MAK 030 (Sigma-Aldrich). 1 μl of supernatant were mixed with 849 μl of distilled water and 150 μl of phosphate reagent in 1 ml cuvette and incubated 30 min for room temperature. After incubation, absorbance at 650 nm was measured. To convert absorbance to actual soluble phosphate concentration, newly phosphate standards for colorimetric detection was required for every batch. To draw a standard curve, the same colorimetric analysis was performed with phosphate solution of 0, 5, 10, 15, 20, and 25 nmol per cuvette. KACC 14533 (Chitinophaga oryziterrae) was regarded as a positive control and ATCC 13032 (Corynebacterium glutamicum) was regarded as a negative control (Chung et al. 2012).
5. Analysis of auxin-producing ability
Based on the nitrogen-fixing ability study and phosphate-solubilizing activity study, isolates with effective nitrogen-fixing ability or phosphate-solublizing ability were conducted auxin producing test based on the method descried by Ali et al.(2009) with some modifications. For the test, isolates were grown in L-broth media at 28 ℃, 150 rpm on a rotary shaking incubator. L-broth media contained (per liter) 10 g of tryptone, 5 g of NaCl, 5 g of yeast extract, 1 g of glucose and adjusted pH to 7.0 with NaOH (Smibert et al. 1994). 100 μl inoculums adjusted to OD600=0.5 prepared by L-broth media
cultivation, were cultivated at L-broth supplemented with L-tryptophan (sterilized by filtering, final concentration 1 mg L-1), 28 ℃, 150 rpm on a
rotary shaking incubator for 7 days. 1 ml of supernatants were collected by centrifugation at 2300 g for 15 minutes at 4 ℃. IAA solutions for creating standard curve for IAA concentration were made at concentration at 0, 10, 20, 50, and 100 μl ml-1 with ethanol. IAA solutions were mixed with 2ml
Salkowski’s reagent and incubated at room temperature for 30 minutes in dark condition. After incubating mixtures in dark, optical density at 530 nm were measured with spectrophotometer and producing standard curve between IAA concentration and OD530. With the same methods, OD530 of
supernatant-Salkowski’s reagent mix were measured and concentration of IAA were calculated by using standard curve. KACC 18104 (Acinetobacter guillouiae) was regarded as a positive control (Kim and Song 2012).
14
6. Analysis of siderophore-producing ability
Confirmation of siderophore-producing ability of selected isolates was performed by streaking on Chrome Azurol S (CAS) agar plate (Schwyn and Neilands 1987). CAS agar media were prepared by description of Alexander and Zuberer (1991). Isolates which decolorized halo zone on CAS agar media, from initial dark blue color to yellowish color, were considered as siderophore excreting bacteria.KACC 11757 (Serratia sp.) was considered as a positive control (Indiragandhi et al. 2007).
7. Analysis of extracellular-protease producing ability
Extracellular-protease production was tested by plate assay using skim milk agar plate. Skim milk agar plate contained (per liter) 28 g of skim milk powder, 5 g of casein, 2.5 g of yeast extract, 1 g of dextrose and 15 g of agar (Sharma et al. 2015). Selected isolates were inoculated on each plate and incubated 28 ℃ incubator. 7 days and 14 days after inoculation, plates were checked for formation of halo zone that indicates inoculated isolate has extracellular protease activity. KACC 17938 (Mucilaginibacter carri) was regarded as a positive control (Kim et al. 2016).
16
8. Analysis of cellulose-degrading ability
Screening for cellulose degrading ability of selected isolates were performed by plate assay using cellulose Congo-red agar plates. Cellulose Congo-red agar contained (per liter) 2 g of cellulose, 2 g of gelatin, 0.5 g of KH2PO4, 0.25 g of Congo-red and 15 g of agar with adjusting pH= 6.8-7.2 before autoclaving (Gupta et al. 2012). 7 days and 14 days after inoculation, plates were checked for formation of halo zone. Decolorization of cellulose-Congo red agar media indicated inoculated isolate has cellulose-degrading ability (Gupta et al. 2012; Lu et al. 2004). KACC 18485 (Oryzihumus soli sp. nov.) was regarded as a positive control (Kim et al. 2017a).
9. Analysis of temperature range for growth and optimal
growth
Selected NFB and PSB were subjected to assessment for growth temperature. Each isolate was liquid cultivated on R2A broth media (LAB M) at 28 ℃, 150 rpm on a rotary shaker. These cultivations were transferred to fresh R2A broth which were cultivated at 4, 10 15, 20, 25, 28, 37, 40, 45, and 50 ℃, 150 rpm on a rotary shaker. Observation of bacterial growth on three days after inoculation was performed to identify its temperature range of optimal growth. Bacteria growth after 14 days were assessed to determine temperature range of growth.
18
RESULTS
1. Isolation and Identification of possible candidates for
NFB and PSB
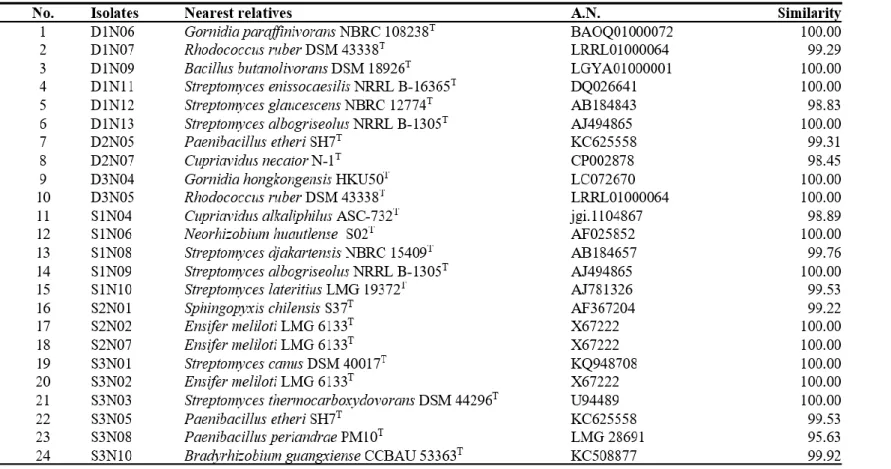
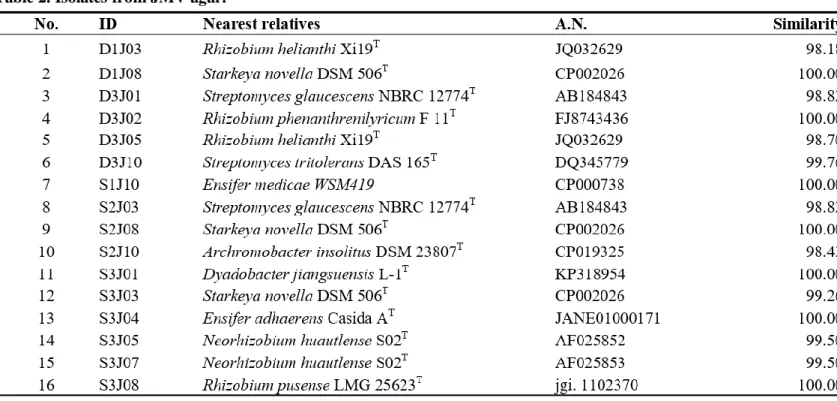
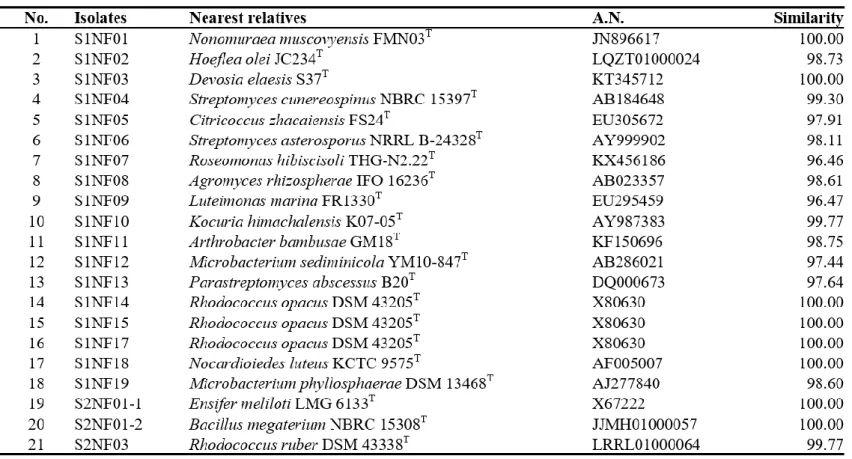
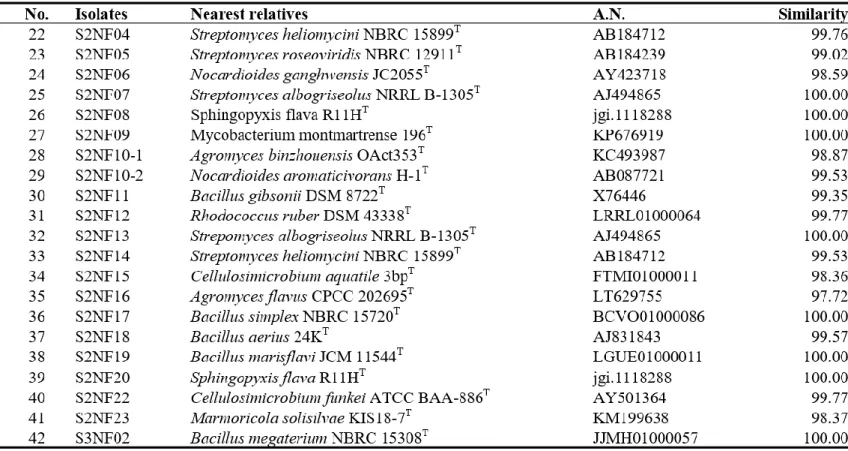
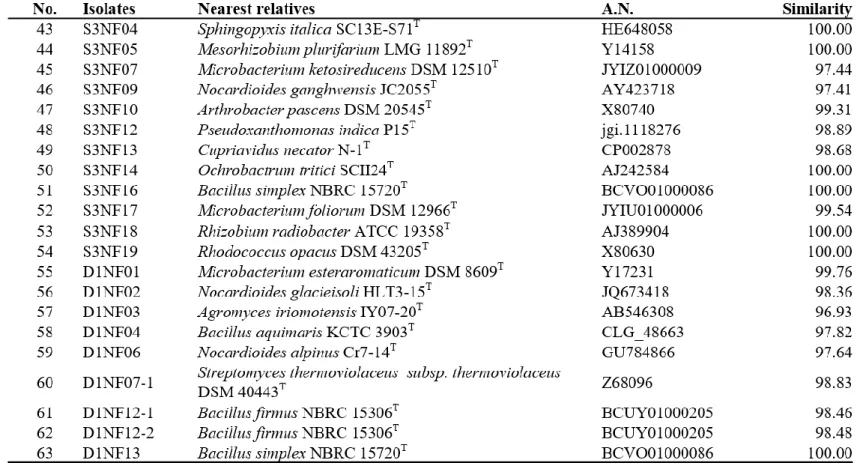
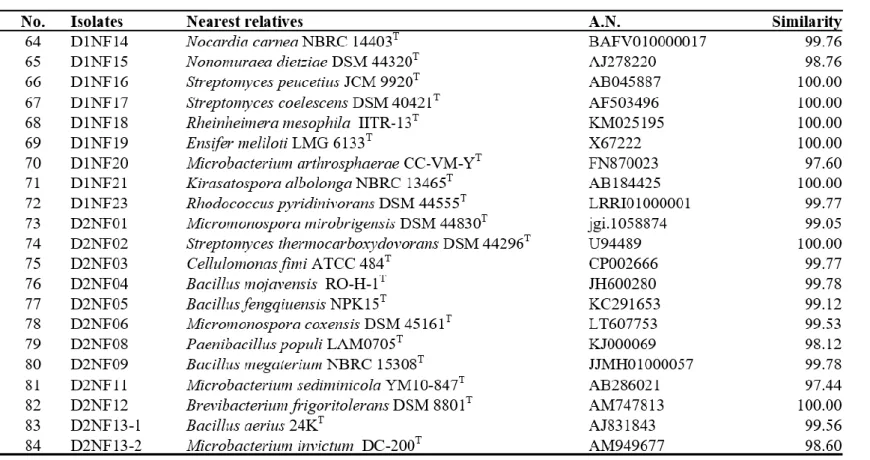
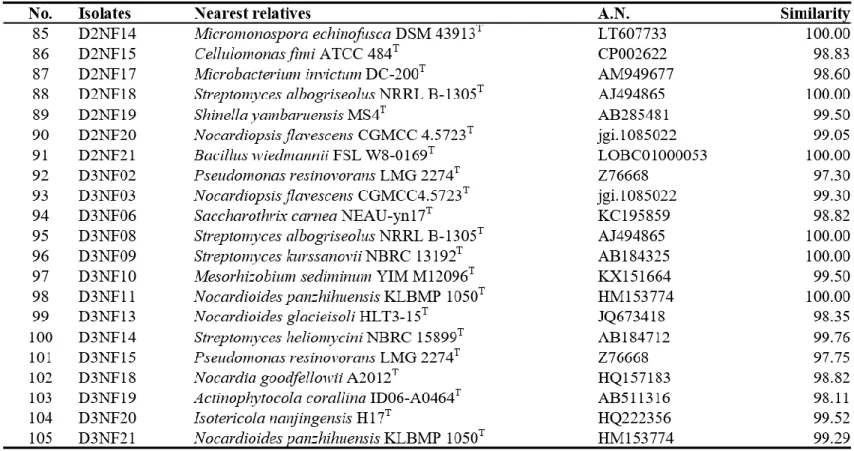
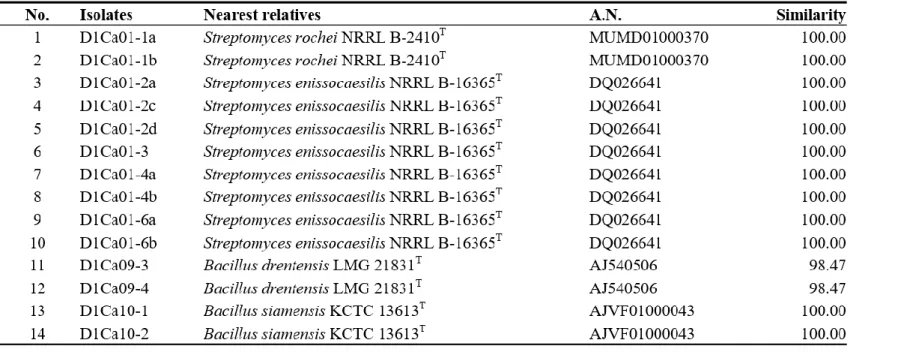
Bacteria were isolated from 6 soil samples by selective media assays and performed colony PCR experiment. Bacteria with identical REP-PCR band were regarded as they are same bacteria at a strain level and only one isolates from them was chosen. A total of 182 strains were selected: 145 strains were possible candidates of NFB and 37 strains were possible candidates of PSB. Isolates considered as NFB consisted of 124 isolates form NFb agar media (Table 1), 6 isolates from JMV agar media (Table 2), and 105 isolates from N2F agar media (Table 3). Isolates considered as PSB consisted of 14 isolates from PVK agar media (Table 4) and 23 isolates from NBRIP-BPB agar media (Table 5). All isolated bacteria were identified by 16S rDNA sequence analysis.
Isolates from NFb agar media showed > 98 % similarity with
Bacillus, Bradyrhizobium, Cupriavidus, Ensifer, Gornida, Neorhizobium, Paenibacillus, Rhodococcus, Sphingopyxis, and Streptomyces species, except
S3N08 which had > 95.63 similarity with Paenibacillus periandrae (Table 1). Isolates from JMV agar media were found to be related to Archromobacter,
Dyadobacter, Ensifer, Neorhizobium, Rhizobium, Starkeya, and Streptomyces,
having > 98 % sequence similarity to known species (Table 2). Isolates from N2F agar medium were related to previously reported species:
Cellulomonas, Cellulomicrobium, Citricoccus, Cupriavidus, Devosia, Ensifer, Hoeflea, Isotericola, Kirasatospora, Kocuria, Luteimonas, Marmoricola,
Mesorhizobium, Microbacterium, Micromonospora, Mycobacterium,
Nocardia, Nocardioides, Nocardiopsis, Nonomuraea, Ochrobactrum,
Paenibacillus, Parastreptomyces, Pseudomonas, Pseudoxanthomonas, Rheinheimera, Rhizobium, Rhodococccus, Roseomonas, Saccharothrix, Shinella, Sphingopyxis, and Streptomyces (Table 3).
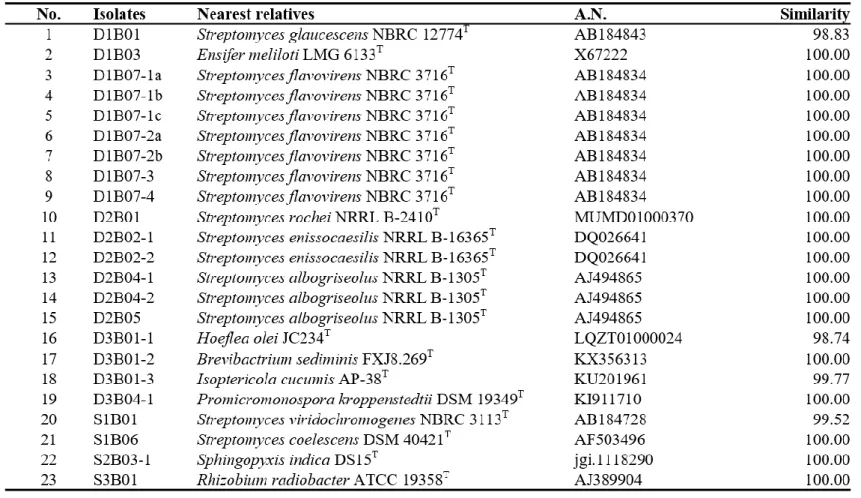
Isolates which form halo zone on PVK agar media showed 98 % > similarity with Bacillus and Streptomyces (Table 4). Isolates decolorizing NBRIP-BPB agar media were determined to be related to formerly Bacillus,
Brevibacterium, Ensifer, Hoelflea, Isoptericola, Promicrimonospora,
Rhizobium, Sphingophyxis, and Streptomyces, having 98 % > sequence
20
22
24
26
28
2. Assessment of nitrogen-fixing ability
The confirmation of the nitrogen-fixing ability of isolated candidates for NFB was carried out by observing of color change of NFb semi-solid agar medium. Growth on NFb agar medium was pretested and bacteria which were able to grow and change the color of NFb agar medium (data not shown) were subjected to NFb semi-solid agar medium assay with positive control of
KACC 11805 (Azotobacter chroococcum) and KACC 16233
(Pleomorphomonas diazotrophica). 22 isolates of total 44 bacteria except two positive controls were exhibited effective color change of NFb semi-solid agar medium, green to blue (Table 6). Two positive controls required different days for effectively color change of the medium: KACC 11805 changed NFb semi-solid agar media color much faster than KACC 16233. Based on how fast and effective color change of NFb semi-solid agar medium, 10 NFB were finally selected: Gordonia paraffinivorans, Streptomyces leeuwenhoekii,
Pseudomonas alcaligenes, Cupriavidus alkaliphilus, Rhodococcus jostii,
Achromobacter xylosoxidans, Nocardioides aromaticivorans, Streptomyces canus, Ochrobactrum tritici, and Rhizobium radiobacter (Table 7).
30
32
3. Assessment of phosphate-solubilizing ability
Validation of phosphate-solubilizing ability was performed by two steps: qualitative assessment by measurement of PSI and quantitative assessment by measurement of soluble phosphate concentration. 37 isolated possible PSB and positive control KACC 14533 (Chitinophaga oryziterrae) were examined by measuring PSIs. All isolates revealed PSI over than 1.00. 21 out of 37 strains showed higher PSI than positive control KACC 14533 (Table 8). With positive control KACC 14533 (Chitinophaga oryziterrae) and negative control ATCC 13032 (Corynebacterium glutamicum) this 21 strains were subjected to quantitative assessment of phosphate-solubilizing ability (Table 9). All selected strains from qualitative assessment were able to solubilize phosphate over than almost twice concentration of negative control, which were lower than 0.5 nmole/μl of soluble phosphate. Based on quantitative estimation of phosphate-solubilizing ability, 15 isolates were selected as promising PSB: Streptomyces glaucescens, Streptomyces
flavovirens, Streptomyces coelescens, Streptomyces viridochromogenes, nine
different strains of Streptomyces enissocaesilis, and two different strains of
34
Table 8. - continued
36
Table 9. Quantitative assessment of phosphate-solubilizing bacteria.
38
4. Analysis of IAA-producing ability of the selected
isolates
All the 10 selected promising NFB and 15 selected promising PSB were performed assessments of other plant growth promoting traits (Table 7, Table 10). Estimation of IAA production was conducted with all 25 selected isolates and positive control KACC 18104 (Acinetobacter guillouiae) which produced over than 140 μg /ml of IAA. All the 25 isolates exhibited low level of IAA concentration compared with positive control: range from S2J10 with 0.68±0.34 μg /ml IAA to S3NF18 with 35.38±0.09 μg /ml IAA (Table 11).
Table 11. Analysis of IAA-producing ability of the selected isolates.
40
5. Analysis of siderophore-producing ability of the
selected isolates
Confirmation of siderophore production was performed with selected isolates and positive control KACC 11757 (Serratia sp.) and bacteria that decolorizing CAS agar media to yellowish color were regarded as positive on siderophore production. All of selected 10 NFB and 15 PSB excreted siderophore on both 7 and 14 days after inoculation (Table 12).
42
Table 12. - continued
6. Analysis of extracellular-protease producing ability of
the selected isolates
Screening of 25 isolates and positive control KACC 17938 (Mucilaginibacter carri) for extracellular protease activity were performed and isolates forming halo zone on skim milk agar media were considered as positive. Three out of 10 NFB and 11 out of 15 PSB showed positive on 7 days after inoculation: Five out of 10 NFB and 14 out of 15 PSB showed positive on 14 days after inoculation (Table 13).
44
Table 13. - continued
46
7. Analysis of cellulose-degrading ability of the selected
isolates
Plate assay of 25 isolates and positive control KACC 18485 (Oryzihumus soli sp. nov.) for cellulose-degrading ability were conducted and isolates forming halo zone on cellulose Congo-red agar media were considered as positive. Three out of 10 NFB and 11 out of 15 PSB showed positive on 7 days after inoculation: Three out of 10 NFB and 12 out of 15 PSB showed positive on 14 days after inoculation (Table 14).
48
Table 14. - continued
8. Analysis of temperature range for growth and optimal
growth of the selected isolates
All the 25 selected bacteria were screened for temperature range of growth: 4, 10, 15, 20, 25, 28, 37, 40, 45, and 50 ℃. All isolates were tended to exhibit weak growth on extreme temperature and almost of isolates had their optimal temperature for growth in range of 28 ℃ to 37 ℃ (Table 15). One PSB and three NFB were able to grow in 4 ℃. Seven PSB and three NFB were grow in 50 ℃ tended to show weak on growth than other settings, however, D1B01, D1B07-2b, D1Ca10-1, D1Ca10-2, D2B02-1, D1N06, and D1N09 showed ordinary growth at 50 ℃ than other isolates which also revealed positive (data not shown).
50
52
DISCUSSIONS
The most part of oriental melon in Korea are produced by cultivation under structure method in Seongju-gun, Gyeongsangbuk-do. Greenhouse cultivation has various benefits, however, salt accumulation occurred in greenhouse soil is a big drawback of oriental melon cultivation: doubled oriental melon production by alleviating soil salinization was reported (RDA and NAS 2014). As a variety of methods have been performed to relieve soil salinization, nowadays, using microbial fertilizer is offered as a reasonable solution. In this study, isolation and characterization of bacteria candidates of microbial fertilizer with various plant growth promoting traits for greenhouse oriental melon cultivation were performed.
Microbial fertilizer market is getting larger, which is predicted as 4.17 billion dollars by 2023. The analysis of the worldwide biofertilizer market shows that biofertilizers with nitrogen-fixing property and phosphate-solubilizing property are the most used, which indicates these two plant growth promoting traits are important in agriculture (Timmusk et al. 2017). Thus, screening PSB and NFB for greenhouse cultivation of oriental melon is effective for selecting bacteria for microbial fertilizer. Assessment for these two important plant growth-promoting properties were carried out and confirmation of other plant growth promoting traits was performed: IAA production, siderophore production, cellulose-degrading ability, and extracellular protease activity.
In many cases of protected cultivation, excessive nutrition is problem than depletion of nutrients. Application of microbial fertilizer can reduce
required amount of chemical fertilizer which is a main reason of salt accumulation in greenhouse soil, stimulate plant growth, and also increase soil fertility (Timmusk et al. 2017). In 2008, the study for evaluation of the effect of inoculating bacteria to wheat with several plant growth promoting properties, phosphorus solubilizing activity, IAA production, ACC-deaminase activity, and chitinase activity was conducted by Shaharoona et al. (2008). As a result, wheat production rate of a group with bacteria inoculation and 75 % of recommended doses of NPK fertilizer shows almost same as that of a group with 100 % of recommended doses of NPK fertilizer with no inoculation of bacteria (Shaharoona et al. 2008). Also, increase in available phosphate concentration and crop productivity by seed inoculation of PGPB with phosphate-solubilizing ability, IAA production, and siderophore production was reported (Kaur and Reddy 2014).
Five plate assays with different composition and pH were conducted for isolating various useful bacteria: total 145 possible NFB were isolated from three selective media tests for NFB (Table 1, Table 2, Table 3) and 37 possible PSB were isolated from two selective media tests for PSB (Table 4, Table 5). The difference between numbers and species range of isolated NFB and PSB might occur because of observation step of halo zone on PSB selective media and fewer plate assay type. As the isolate S3N08 was shown to have low similarity (95.63 %) with Paenibacillus periandrae, this isolate was considered as a novel species. Morphological, physiological and biochemical characterization of S3N08 is underway. S3N08 was deposited in
54
the Korean Agricultural Culture Collection and NITE Biological Resource Center.
There are various application methods of microbial fertilizer, however, inoculated bacteria have to compete with native bacteria that already occupied in rhizosphere. In this study, bacteria were isolated from the target area of microbial fertilizer for oriental melon. Therefore, successful stable settlement and sustainability of PGPB as microbial fertilizer is expected. Based on the screening for nitrogen-fixing ability and phosphate-solubilizing ability of 182 isolates, 10 NFB and 15 PSB were finally selected (Table 7, Table 10). Among selected 10 NFB, S3N01 (Streptomyces canus), S3NF14 (Ochrobactrum tritici), S3NF18 (Rhizobium radiobacter) are considered as having various plant growth promoting traits, and among selected 15 PSB, D1Ca01-4a and D2B02-1, different strains of Streptomyces enissocaesilis, were regarded to have various plant growth promoting traits.
As the condition for oriental melon cultivation is similar with that for microbial growth, managing rhizosphere microorganism is essential (GBARES 2015). This can be advantage for beneficial microorganisms but also for pathogen. The usage of microbial fertilizer is expected to protect oriental melon from harmful microbes by competition with pathogens. In addition, siderophore-excreting bacteria have much advantages in competition especially with plant fungal pathogen which are commonly unable to uptake siderophore that sequestered iron ion (Kloepper et al. 1980; Leong 1986). All of the selected isolates in this study produced siderophore (Table 12). Also, suppression of pathogen by extracellular protease or cellulose which produced
by rhizobacteria was reported (Gupta et al. 2015; Jung et al. 2007). Thus, isolates with extracellular protease activity or cellulase are expected to protect roots from pathogens (Table 13, Table 14).
The effectiveness of applicating PGPB with IAA-producing ability was reported many times. All the selected isolates were produced IAA which is essential for plant growth (Table 11). As gibberelins are considered as important phytohormone for oriental melon fruiting along with auxin, further screening for gibberellin production of isolates will increase productivity of microbial fertilizer (GBARES 2015).
Temperature range of growth and optimal growth of the selected bacteria was analyzed, which provide further information for constructing efficient microbial fertilizer application methods. Cultivation condition of oriental melon in greenhouse has wide range of temperature. Oriental melons are fruit that prefer an air temperature near 30 ℃ and can tolerate heat up to 40 ℃ during daytime. Optimal air temperature at night of oriental melon cultivation is 18-20 ℃, so 12-16 ℃ at night is appropriate in terms of economy. The temperature range of root growth is reported as 8-40 ℃, and the ideal temperature range is known as 20-25 ℃ (GBARES 2015). As growth temperature ranges of selected isolates are comparable with that of oriental melon (Table 15), promising plant growth promoting results and adaptability of isolates is predicted. Various application is expected as possible because of growth temperatures of isolates is diverse and wide-ranged. As temperature in greenhouse soil occasionally reaches to 50 ℃,
56
strains which express favorable growth at 50 ℃ are worthy of notice (GBARES 2015).
However, in this study, pot trial or field trial for confirmation of increases in crop productivity and soil fertility by inoculation was not performed. To suggest finer candidates for microbial fertilizer, additional research about optimal coordination of isolates and their property is required.
Although nitrogen-fixing activity was tested by NFb semi-solid agar media method, additional assessment is needed. The test of nitrogen-fixing ability was performed only to the bacteria which can effectively grow on NFb agar media, because this method was based on observation of color change of NFb semi-solid agar media that the results can be made only if inoculated bacteria can grow in NFb medium. As only a part of NFB in soil can cultivated on a specific medium and this method cannot quantify nitrogen-fixing capability, a follow-up experiment for nitrogen-nitrogen-fixing capability is required.
In conclusion, isolation and characterization of plant growth promoting bacteria from oriental melon farm soil in this study provide useful information for microbial fertilizer development for greenhouse oriental melon cultivation in Korea.
LITERATURE CITED
Alexander, D., and Zuberer, D. 1991. Use of chrome azurol S reagents to evaluate siderophore production by rhizosphere bacteria. Biology and Fertility of soils 12:39-45.
Ali, B., Sabri, A., Ljung, K., and Hasnain, S. 2009. Auxin production by plant associated bacteria: impact on endogenous IAA content and growth of Triticum aestivum L. Letters in applied microbiology 48:542-547. Anwar, S., Ali, B., and Sajid, I. 2016. Screening of rhizospheric
actinomycetes for various in-vitro and in-vivo plant growth promoting (PGP) traits and for agroactive compounds. Frontiers in microbiology 7:1334.
Armstrong, D., Griffin, K., Danner, M., Mees, C., and Nguyen, D. 1999. Effects of Phosphorus on Nitrogen Fixation. Better Crops with Plant Food 83:30-31.
Baker, G., Smith, J. J., and Cowan, D. A. 2003. Review and re-analysis of domain-specific 16S primers. Journal of microbiological methods 55:541-555.
Baldani, J. I., Reis, V. M., Videira, S. S., Boddey, L. H., and Baldani, V. L. D. 2014. The art of isolating nitrogen-fixing bacteria from non-leguminous plants using N-free semi-solid media: a practical guide for microbiologists. Plant and Soil 384:413-431.
Bernstein, L. 1975. Effects of salinity and sodicity on plant growth. Annual review of phytopathology 13:295-312.
58
Cho, K.-R., Kang, C.-S., Won, T.-J., and Park, K.-Y. 2006. Effects of compressed expansion rice hull application and drip irrigation on the alleviation of salt accumulation in the plastic film house soil. Korean Journal of Soil Science and Fertilizer 39:372-379.
Choi, E., Lee, H., Yoon, K., Kwon, D., Sohn, J., Park, S., Han, M., and Ghim, S. 2003. Isolation of nitrogen-fixing bacteria from gramineous crops and measurement of nitrogenase activity. Korean Journal of Microbiology Biotechnology.
Chun, J., Lee, J.-H., Jung, Y., Kim, M., Kim, S., Kim, B. K., and Lim, Y.-W. 2007. EzTaxon: a web-based tool for the identification of prokaryotes based on 16S ribosomal RNA gene sequences. International journal of systematic and evolutionary microbiology 57:2259-2261.
Chung, E. J., Park, T. S., Jeon, C. O., and Chung, Y. R. 2012. Chitinophaga oryziterrae sp. nov., isolated from the rhizosphere soil of rice (Oryza sativa L.). International journal of systematic and evolutionary microbiology 62:3030-3035.
Damir, O., Mladen, P., Božidar, S., and Srñan, N. 2011. Cultivation of the bacterium Azotobacter chroococcum for preparation of biofertilizers. African Journal of Biotechnology 10:3104-3111.
De Bruijn, F. J. 1992. Use of repetitive (repetitive extragenic palindromic and enterobacterial repetitive intergeneric consensus) sequences and the polymerase chain reaction to fingerprint the genomes of Rhizobium meliloti isolates and other soil bacteria. Appl. Environ. Microbiol. 58:2180-2187.
Dobereiner, J., Marriel, I., and Nery, M. 1976. Ecological distribution of Spirillum lipoferum Beijerinck. Canadian Journal of Microbiology 22:1464-1473.
EA, P., and Clark, F. 1989. Soil microbiology and biochemistry. Academic Press;.
GBARES. 2015. A series of Korean melon. Gyeongsangbuk-do agricultural research & extension services.
Gupta, G., Parihar, S. S., Ahirwar, N. K., Snehi, S. K., and Singh, V. 2015. Plant growth promoting rhizobacteria (PGPR): current and future prospects for development of sustainable agriculture. J Microb Biochem Technol 7:096-102.
Gupta, P., Samant, K., and Sahu, A. 2012. Isolation of cellulose-degrading bacteria and determination of their cellulolytic potential. International journal of microbiology 2012.
Indiragandhi, P., Anandham, R., Madhaiyan, M., Poonguzhali, S., Kim, G., Saravanan, V., and Sa, T. 2007. Cultivable bacteria associated with larval gut of prothiofos‐resistant, prothiofos‐susceptible and field‐caught populations of diamondback moth, Plutella xylostella and their potential for, antagonism towards entomopathogenic fungi and host insect nutrition. Journal of Applied Microbiology 103:2664-2675. Jeon, W.-T., Park, H.-M., Chung, J.-B., Park, K.-D., Park, C.-Y., and Yang,
W.-H. 2005. Effect of reduced fertilization considering residual soil nutrients on rice yield and salt removal in greenhouse vegetables and
60
rice cropping system. Korean Journal of Soil Science and Fertilizer 38:85-91.
Jung, H.-K., Kim, J.-R., Woo, S.-M., and Kim, S.-D. 2007. Selection of the auxin, siderophore, and cellulase-producing PGPR, Bacillus licheniformis K11 and its plant growth promoting mechanisms. Applied Biological Chemistry 50:23-28.
Kim, D.-U., Kim, S.-G., Lee, H., Park, A.-Y., and Ka, J.-O. 2017a. Oryzihumus soli sp. nov., isolated from soil and emended description of the genus Oryzihumus. International journal of systematic and evolutionary microbiology 67:3960-3964.
Kim, D.-U., Lee, H., Kim, H., Kim, S.-G., Park, S. Y., and Ka, J.-O. 2016. Mucilaginibacter carri sp. nov., isolated from a car air conditioning system. International journal of systematic and evolutionary microbiology 66:1754-1759.
Kim, H., Kim, D.-U., Lee, H., Yun, J., and Ka, J.-O. 2017b. Syntrophic biodegradation of propoxur by Pseudaminobacter sp. SP1a and Nocardioides sp. SP1b isolated from agricultural soil. International Biodeterioration & Biodegradation 118:1-9.
Kim, N. H., Kim, D.-U., Kim, I., and Ka, J.-O. 2013. Syntrophic biodegradation of butachlor by Mycobacterium sp. J7A and Sphingobium sp. J7B isolated from rice paddy soil. FEMS microbiology letters 344:114-120.
Kim, W.-J., and Song, H.-G. 2012. Interactions between biosynthetic pathway and productivity of IAA in some rhizobacteria. The Korean Journal of Microbiology 48:1-7.
Kim, Y.-K., Hong, S.-J., Shim, C.-K., Kim, M.-J., Choi, E.-J., Lee, M.-H., Park, J.-H., Han, E.-J., An, N.-H., and Jee, H.-J. 2012. Functional analysis of Bacillus subtilis isolates and biological control of red pepper powdery mildew using Bacillus subtilis R2-1. Research in Plant Disease 18:201-209.
Kogan, S. C., Doherty, M., and Gitschier, J. 1987. An improved method for prenatal diagnosis of genetic diseases by analysis of amplified DNA sequences. New England Journal of Medicine 317:985-990.
KOSIS. 2014. Korean Statistical Information Service Statistical DB. KOSIS. 2019. Korean Statistical Information Service Statistical DB.
Kumar, V., and Narula, N. 1999. Solubilization of inorganic phosphates and growth emergence of wheat as affected by Azotobacter chroococcum mutants. Biology and Fertility of Soils 28:301-305.
Lee, K.-K., Mok, I.-K., Yoon, M.-H., Kim, H.-J., and Chung, D.-Y. 2012. Mechanisms of phosphate solubilization by PSB (Phosphate-solubilizing Bacteria) in soil. Korean Journal of Soil Science and Fertilizer 45:169-176.
Lu, W.-J., Wang, H.-T., Nie, Y.-F., Wang, Z.-C., Huang, D.-Y., Qiu, X.-Y., and Chen, J.-C. 2004. Effect of inoculating flower stalks and vegetable waste with ligno-cellulolytic microorganisms on the composting
62
process. Journal of Environmental Science and Health, Part B 39:871-887.
Madhaiyan, M., Jin, T. Y., Roy, J. J., Kim, S.-J., Weon, H.-Y., Kwon, S.-W., and Ji, L. 2013. Pleomorphomonasdiazotrophica sp. nov., an endophytic N-fixing bacterium isolated from root tissue of Jatropha curcas L. International journal of systematic and evolutionary microbiology 63:2477-2483.
Mehta, S., and Nautiyal, C. S. 2001. An efficient method for qualitative screening of phosphate-solubilizing bacteria. Current microbiology 43:51-56.
Oh, S.-E., Son, J.-S., Ok, Y.-S., and Joo, J.-H. 2010. A modified methodology of salt removal through flooding and drainage in a plastic film house soil. Korean Journal of Soil Science and Fertilizer 43:565-571.
Rademaker, J., Louws, F. J., and De Bruijn, F. 1998. Characterization of the diversity of ecologically important microbes by rep-PCR genomic fingerprinting. Molecular microbial ecology manual 3:1-27.
Rakshit, A., and Singh, H. B. 2018. Advances in Seed Priming. Springer. Raun, W. R., and Johnson, G. V. 1999. Improving nitrogen use efficiency for
cereal production. Agronomy journal 91:357-363.
RDA. 2004. Annual Report of the Monitoring Project on Agricultural Environment Quality.p. 9-12.
RDA, and NAS. 2014. (농가현장 애로해결을 위한) 염류집적 토양관리 길잡이 :영농활용자료와 농가실행 사례. Department of Agricultural Environment, NAS, Suwon.
Rosswall, T. 1976. THE INTERNAL NITROGEN CYCLE
BETWEENMICROORGANISMS, VEGETATIONAND SOIL.
Nitrogen, phosphorus, and sulfur: global cycles. SCOPE Rep 7:157-167.
Schwyn, B., and Neilands, J. 1987. Universal chemical assay for the detection and determination of siderophores. Analytical biochemistry 160:47-56. Shaharoona, B., Naveed, M., Arshad, M., and Zahir, Z. A. 2008.
Fertilizer-dependent efficiency of Pseudomonads for improving growth, yield, and nutrient use efficiency of wheat (Triticum aestivum L.). Applied microbiology and biotechnology 79:147-155.
Sharma, A. K., Sharma, V., Saxena, J., Yadav, B., Alam, A., and Prakash, A. 2015. Isolation and screening of extracellular protease enzyme from bacterial and fungal isolates of soil. Int. J. Sci. Res. Environ. Sci 3:0334-0340.
Smibert, R., Krieg, N., Gerhardt, P., Murray, R., and Wood, W. 1994. Methods for general and molecular bacteriology. American Society for Microbiology, Washington DC:171-173.
Sonneveld, C., and Voogt, W. 2009. Plant nutrition in future greenhouse production. Pages 393-403 in: Plant Nutrition of Greenhouse Crops. Springer.
Stackebrandt, E., and Goodfellow, M. 1991. Nucleic acid techniques in bacterial systematics. Wiley.
Surange, S., Wollum Ii, A., Kumar, N., and Nautiyal, C. S. 1997. Characterization of Rhizobium from root nodules of leguminous trees
64
growing in alkaline soils. Canadian Journal of Microbiology 43:891-894.
Timmusk, S., Behers, L., Muthoni, J., Muraya, A., and Aronsson, A.-C. 2017. Perspectives and challenges of microbial application for crop improvement. Frontiers in plant science 8:49.
Yoon, S.-H., Ha, S.-M., Kwon, S., Lim, J., Kim, Y., Seo, H., and Chun, J. 2017. Introducing EzBioCloud: a taxonomically united database of 16S rRNA gene sequences and whole-genome assemblies. International journal of systematic and evolutionary microbiology 67:1613.
유철성. 1996. 고소득 채소 시설재배. Osung publishing house.
이병일, 문원, and 손정익. 2004. 시설재배학. Korea national open university.
참외 농장 토양에서의 질소고정균과
인산 가용화 균의 분리 및 특성 연구
장소현
초록
한국의 7 대 과채류 중 하나인 참외는 국내 생산량 중 약 85 %가 성주군에서 출하되며 대부분 시설재배로 재배된다. 하지만 시설 재배 시 염류 집적 등으로 인한 참외 생산량 감소가 야기되기도 한다. 이러한 염류 집적를 해소하기 위하여 인산 가용화 균주를 포함한 미생물 비료 시비가 제시되었다. 미생물 비료는 인산 가용화 및 질소고정, 옥신 생산, siderophore 생산, 셀룰로오스 분해, 세포 외 단백질 분해효소 분비 등의 다양한 식물 촉진 특성을 보유한 균주를 사용하여 작물의 생산성을 증대시킬 뿐만 아니라 화학비료의 시비량을 절감시켜 환경 보전에 기여할 수 있다. 따라서 본 연구는 참외 농장 토양에서 균주들을 분리한 후 이들의 특성 연구를 통하여 참외 시설재배지용 미생물 비료 후보 균주를 선발하고자 진행되었다. 연구 결과, 성주군에 위치한 참외 농장 토양에서 채취한 6 개의 시료에서 NFb, JMV, N2F 배지를 이용하여 질소고정 후보 균주들을 분리하였으며, PVK, NBRIP 배지에서의 투명대 형성 확인으로 인산가용화 후보 균주들을 분리하였다. 분리한 균주를 대상으로 REP-PCR 분석과 16S rDNA 염기서열 분석을 통하여 질소고정 후보 균주 145 균주와 인산가용화 후보 균주 37 균주를 분리 및 동정하였다. 질소고정 후보 균주들은 NFb 배지에서의 배양이 확인된 균주들을 대상으로66
NFb 반고체 배지의 색 변화를 이용하여 질소고정능 평가를 진행한 후 총 10 균주를 선발하였다. 인산 가용화 후보 균주는 NBRIP-BPB 배지에서의 PSI(phosphate solubilization index)를
통하여 균주를 1 차적으로 선별한 후, 인산 비색 측정
키트(MAK030)을 이용한 정량적 시험 결과를 바탕으로
인산가용화 균주 15 균주를 선발하여 이후 추가적인 식물 생장 촉진 특성 보유 여부 실험을 진행하였다. Siderophore 생산은 CAS 배지를 이용하여 확인하였으며, salkowski’s reagent 를 이용하여 균주의 옥신 생산량을 측정하였다. 또한 skim-milk 배지의 투명대 형성 여부로 세포 외 단백질 분해효소 분비 여부와 cellulose Congo-red 배지의 투명대 형성 여부로 셀룰로오스 분해능 보유 여부를 확인하였다. 또한 각 균주의 생장 온도와 최적 생장 온도를 확인하였다. 그 결과 선발한 모든 균주에서 siderophore 생산과 낮은 수준의 옥신 생산이 확인되었다. 또한 선발한 25 균주 증 19 균주가 세포 외 단백질 분해효소를 분비하였으며, 15 균주가 셀룰로오스 분해능을 나타내었다. 또한 미생물 비료의 적용 방법 결정에 도움이 될 선발된 균주들의 생장 온도와 최적 생장 온도를 확인하였다. 본 연구로 선발된 균주들은 참외 시설재배지 대상 미생물 비료 후보 미생물 자원으로 사용될 수 있을 것으로 생각된다. 주요어: 인산 가용화, 질소고정, 미생물 비료, 참외, 식물 생장 촉진 미생물 학 번: 2018-25974